

Phytochemical Evaluation of Terminalia canescens DC. Radlk. Extracts with Antibacterial and Antibiotic Potentiation Activities against Selected β-Lactam Drug-Resistant Bacteria

Abstract
:1. Introduction
2. Results
2.1. Antimicrobial Susceptibility Studies
2.2. Calculation of Fractional Inhibitory Concentration
2.3. Evaluation of Extract and Antibiotic Synergistic Interactions at Different Ratios
2.4. Identification of Compounds in the TCAM and TCAW Extracts
2.5. Quantification of Toxicity
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Plant Collection and Extraction
4.3. Antibacterial Studies
4.3.1. Growth of Bacterial Cultures
4.3.2. Disc Diffusion Assay
4.3.3. Liquid Microdilution MIC Assay
4.4. Determination of Combinational Effects and Optimal Ratios through Isobologram
4.5. Non-Targeted Head Space LC-MS Conditions for Quantitative Analysis
4.6. Toxicity Studies
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Akova, M. Epidemiology of antimicrobial resistance in bloodstream infections. Virulence 2016, 7, 252–266. [Google Scholar] [CrossRef]
- Finch, R.G.; Metlay, J.P.; Davey, P.G.; Baker, L.J. Educational interventions to improve antibiotic use in the community: Report from the International Forum on Antibiotic Resistance (IFAR) colloquium, 2002. Lancet Infect. Dis. 2004, 4, 44–53. [Google Scholar] [CrossRef]
- Holmberg, S.D.; Solomon, S.L.; Blake, P.A. Health and economic impacts of antimicrobial resistance. Rev. Infect. Dis. 1987, 9, 1065–1078. [Google Scholar] [CrossRef]
- Michael, C.A.; Dominey-Howes, D.; Labbate, M. The antimicrobial resistance crisis: Causes, consequences, and management. Front. Public Health 2014, 2, 145. [Google Scholar] [CrossRef]
- Spellberg, B.; Srinivasan, A.; Chambers, H.F. New societal approaches to empowering antibiotic stewardship. JAMA 2016, 315, 1229–1230. [Google Scholar] [CrossRef]
- Hoffman, S.J.; Caleo, G.M.; Daulaire, N.; Elbe, S.; Matsoso, P.; Mossialos, E.; Rizvi, Z.; Røttingen, J.-A. Strategies for achieving global collective action on antimicrobial resistance. Bull. World Health Organ. 2015, 93, 867–876. [Google Scholar] [CrossRef] [PubMed]
- Chandra, H.; Bishnoi, P.; Yadav, A.; Patni, B.; Mishra, A.P.; Nautiyal, A.R. Antimicrobial resistance and the alternative resources with special emphasis on plant-based antimicrobials—A review. Plants 2017, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.M.; COSTA, P.A.; Ribon, A.O.; Purgato, G.A.; Gaspar, D.-M.; Diaz, M.A. Plant extracts display synergism with different classes of antibiotics. An. Acad. Bras. Ciências 2019, 91, e20180117. [Google Scholar] [CrossRef] [PubMed]
- Eloff, J.; Katerere, D.; McGaw, L. The biological activity and chemistry of the southern African Combretaceae. J. Ethnopharmacol. 2008, 119, 686–699. [Google Scholar] [CrossRef] [PubMed]
- Garcez, F.R.; Garcez, W.S.; Miguel, D.L.; Serea, A.A.; Prado, F.C. Chemical constituents from Terminalia glabrescens. J. Braz. Chem. Soc. 2003, 14, 461–465. [Google Scholar] [CrossRef]
- Cheesman, M.J.; White, A.; Matthews, B.; Cock, I.E. Terminalia ferdinandiana fruit and leaf extracts inhibit methicillin-resistant Staphylococcus aureus growth. Planta Medica 2019, 85, 1253–1262. [Google Scholar] [CrossRef]
- Zai, M.J.; Cheesman, M.J.; Cock, I.E. Terminalia petiolaris A.Cunn ex Benth. extracts have antibacterial activity and potentiate conventional antibiotics against β-lactam-drug-resistant bacteria. Antibiotics 2023, 12, 1643. [Google Scholar] [CrossRef] [PubMed]
- Eloff, J. Which extractant should be used for the screening and isolation of antimicrobial components from plants? J. Ethnopharmacol. 1998, 60, 1–8. [Google Scholar] [CrossRef] [PubMed]
- van Vuuren, S.; Viljoen, A. Plant-based antimicrobial studies–methods and approaches to study the interaction between natural products. Planta Medica 2011, 77, 1168–1182. [Google Scholar] [CrossRef] [PubMed]
- Bonev, B.; Hooper, J.; Parisot, J. Principles of assessing bacterial susceptibility to antibiotics using the agar diffusion method. J. Antimicrob. Chemother. 2008, 61, 1295–1301. [Google Scholar] [CrossRef] [PubMed]
- Cheesman, M.J.; Ilanko, A.; Blonk, B.; Cock, I.E. Developing new antimicrobial therapies: Are synergistic combinations of plant extracts/compounds with conventional antibiotics the solution? Pharmacogn. Rev. 2017, 11, 57. [Google Scholar]
- Rayan, P.; Matthews, B.; McDonnell, P.; Cock, I. Terminalia ferdinandiana extracts as inhibitors of Giardia duodenalis proliferation: A new treatment for giardiasis. Parasitol. Res. 2015, 114, 2611–2620. [Google Scholar] [CrossRef]
- Kwast, K.E.; Hand, S.C. Oxygen and pH regulation of protein synthesis in mitochondria from Artemia franciscana embryos. Biochem. J. 1996, 313, 207–213. [Google Scholar] [CrossRef]
- Cushnie, T.T.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef]
- Tereschuk, M.a.L.; Riera, M.V.; Castro, G.R.; Abdala, L.R. Antimicrobial activity of flavonoids from leaves of Tagetes minuta. J. Ethnopharmacol. 1997, 56, 227–232. [Google Scholar] [CrossRef]
- LeBel, M. Ciprofloxacin: Chemistry, mechanism of action, resistance, antimicrobial spectrum, pharmacokinetics, clinical trials, and adverse reactions. Pharmacother. J. Hum. Pharmacol. Drug Ther. 1988, 8, 3–30. [Google Scholar] [CrossRef]
- El-Abyad, M.S.; Morsi, N.M.; Zaki, D.A.; Shaaban, M. Preliminary screening of some Egyptian weeds for antimicrobial activity. Microbios 1990, 62, 47–57. [Google Scholar] [PubMed]
- Dall’Agnol, R.; Ferraz, A.; Bernardi, A.; Albring, D.; Nör, C.; Sarmento, L.; Lamb, L.; Hass, M.; Von Poser, G.; Schapoval, E. Antimicrobial activity of some Hypericum species. Phytomedicine 2003, 10, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Kumar, S. Perspective on plant products as antimicrobials agents: A review. Pharmacologia 2013, 4, 469–480. [Google Scholar] [CrossRef]
- Tsuchiya, H. Membrane interactions of phytochemicals as their molecular mechanism applicable to the discovery of drug leads from plants. Molecules 2015, 20, 18923–18966. [Google Scholar] [CrossRef]
- Scalbert, A. Antimicrobial properties of tannins. Phytochemistry 1991, 30, 3875–3883. [Google Scholar] [CrossRef]
- Jöbstl, E.; Howse, J.R.; Fairclough, J.P.A.; Williamson, M.P. Noncovalent cross-linking of casein by epigallocatechin gallate characterized by single molecule force microscopy. J. Agric. Food Chem. 2006, 54, 4077–4081. [Google Scholar] [CrossRef]
- Farha, A.K.; Yang, Q.-Q.; Kim, G.; Li, H.-B.; Zhu, F.; Liu, H.-Y.; Gan, R.-Y.; Corke, H. Tannins as an alternative to antibiotics. Food Biosci. 2020, 38, 100751. [Google Scholar] [CrossRef]
- Lewis, K. Riddle of biofilm resistance. Antimicrob. Agents Chemother. 2001, 45, 999–1007. [Google Scholar] [CrossRef]
- Bakkiyaraj, D.; Nandhini, J.R.; Malathy, B.; Pandian, S.K. The anti-biofilm potential of pomegranate (Punica granatum L.) extract against human bacterial and fungal pathogens. Biofouling 2013, 29, 929–937. [Google Scholar] [CrossRef]
- NHMRC. Australian Code of Practice for the Care and Use of Animals for Scientific Purposes; National Health and Medical Research Council: Canberra, Australia, 2004; pp. 3–28.





{kind=link}
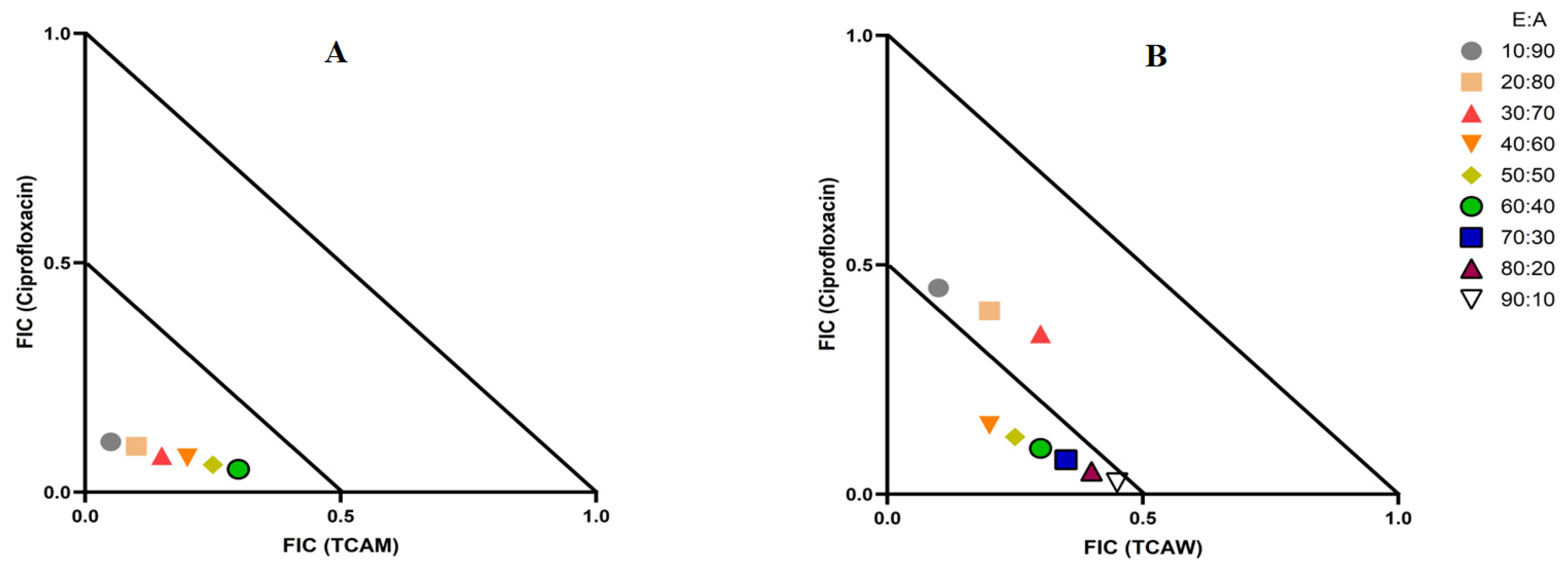{kind=link}
{kind=link}
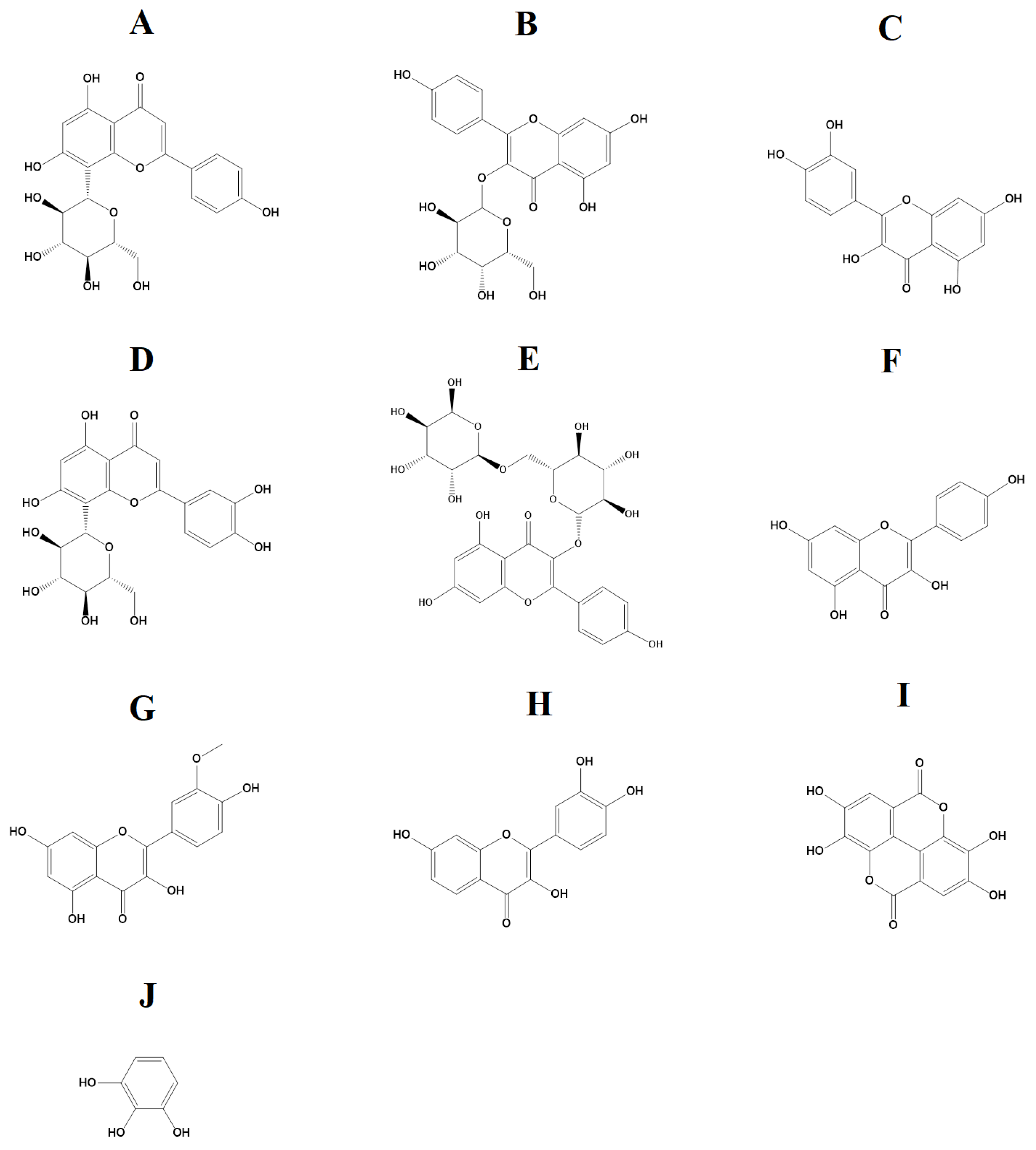{kind=link}
| Extract and Antibiotic | MIC (µg/mL) | |||||
|---|---|---|---|---|---|---|
| E. coli | ESBL E. coli | S. aureus | MRSA | K. pneumoniae | ESBL K. pneumoniae | |
| TCAM | 1919 | 1919 | 960 | 1919 | 960 | 960 |
| TCAW | 2050 | 2050 | 1025 | 2050 | 2050 | 2050 |
| TCAE | - | - | - | - | - | - |
| Tetracycline | - | - | 1.25 | - | - | - |
| Chloramphenicol | - | - | 0.31 | - | 1.25 | 1.25 |
| Ciprofloxacin | 2.5 | - | 0.62 | 2.5 | 2.5 | 1.25 |
| Gentamicin | 0.039 | 0.039 | 0.03 | 0.03 | 0.03 | 0.03 |
| Erythromycin | - | - | 1.25 | - | 2.5 | - |
| Negative control | - | - | - | - | - | - |
| Bacteria | Extract | Tetracycline | Chloramphenicol | Ciprofloxacin | Gentamicin | Erythromycin |
|---|---|---|---|---|---|---|
| E. coli | TCAM | - | - | 0.31 | 2.13 | - |
| TCAW | - | - | 0.75 | 2.1 | ||
| TCAE | - | - | - | - | - | |
| ESBL E. coli | TCAM | - | - | - | 2.72 | - |
| TCAW | - | - | - | 2.66 | - | |
| TCAE | - | - | - | - | - | |
| S. aureus | TCAM | 0.63 | 2 | 1.50 | 5.70 | 0.63 |
| TCAW | 0.75 | 3 | 2 | 22 | 1.5 | |
| TCAE | - | - | - | - | - | |
| MRSA | TCAM | - | - | 0.62 | 5.45 | - |
| TCAW | - | - | 0.75 | 5.32 | - | |
| TCAE | - | - | - | - | - | |
| K. pneumoniae | TCAM | - | 1.25 | 1.12 | 11.4 | 0.56 |
| TCAW | - | 1 | 0.37 | 10.6 | 0.75 | |
| TCAE | - | - | - | - | - | |
| ESBL K. pneumoniae | TCAM | - | 1.25 | - | 5.70 | - |
| TCAW | - | 1 | - | 5.33 | - | |
| TCAE | - | - | - | - | - |
| Retention Time (Min) | Empirical Formula | Molecular Mass | Putative Identification | Relative Abundance (% Total Area) | ||
|---|---|---|---|---|---|---|
| TCAM | TCAW | |||||
| Flavonoids | 6.18 | C21H20O10 | 432 | Vitexin | 1.38 | |
| 7.07 | C21H20O11 | 448 | Trifolin | 3.82 | ||
| 6.31 | C21H20O12 | 464 | Quercitin-3β-d-glucoside | 0.99 | ||
| 6.36 | C27H30O16 | 610 | Quercitin 3-O-rhamnoside-7 -O-glucoside | 0.50 | ||
| 6.31 | C15H10O7 | 302 | Quercetin | 0.64 | ||
| 5.54 | C21H20O11 | 448 | Orientin | 7.91 | ||
| 7.14 | C27H30O15 | 594 | Nictoflorin | 0.98 | ||
| 7.13 | C15H10O6 | 286 | Kaempferol | 0.19 | ||
| 7.24 | C16H12O7 | 316 | Isorhamnetin | 0.31 | ||
| 8.51 | C22H20O12 | 476 | Hispidulin 7-glucuronide | 2.53 | ||
| 7.07 | C15H10O6 | 286 | Fisetin | 1.10 | ||
| 6.10 | C21H18O14 | 494 | 8-Hydroxytricetin 7-glucuronide | 0.04 | ||
| 7.25 | C22H22O12 | 478 | 5,7-Dihydroxy-2-(4-hydroxy-3-methoxyphenyl)-3-{[3,4,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxy}-4H-chromen-4-one | 0.43 | ||
| 6.88 | C21H20O11 | 448 | 4-(3,4-Dihydroxyphenyl)-7-hydroxy-5-{[(2S,3R,4S,5S,6R)-3,4,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxy}-2H-chromen-2-one | 3.46 | ||
| 6.19 | C15H10O9 | 334 | 3,5,6,7,2′,3′,4′-Heptahydroxyflavone | 0.08 | ||
| 5.27 | C20H18O13 | 466 | 2-(3,4-Dihydroxyphenyl)-3,5,7-trihydroxy-8-{[(2R,3R,4S,5S,6R)-3,4,5,6-tetrahydroxytetrahydro-2H-pyran-2-yl]oxy}-4H-chromen-4-one | 0.04 | ||
| 6.58 | C28H24O14 | 584 | 2″-O-Galloylisovitexin | 0.19 | ||
| 7.43 | C21H18O12 | 462 | (2S,3S,4S,5R,6S)-6-{[5,7-Dihydroxy-2-(4-hydroxyphenyl)-4-oxo-4H-chromen-3-yl]oxy}-3,4,5-trihydroxyoxane-2-carboxylic acid | 0.13 | ||
| 9 | C15H10O7 | 302 | Quercetin | 0.18 | ||
| 6.29 | C21H20O11 | 448 | Orientin | 9.77 | ||
| 0.35 | C21H20O14 | 496 | Hibiscetin 3-glucoside | 0.49 | ||
| 13.70 | C18H14O9 | 374 | Gossypetin 7-methyl ether 8-acetate | 0.02 | ||
| 10.03 | C15H10O6 | 286 | Fisetin | 0.11 | ||
| 7.63 | C20H22O5 | 342 | Brosimacutin C | 0.02 | ||
| 10.61 | C16H12O7 | 316 | 2-(3,4-dihydroxyphenyl)-3,5,7-trihydroxy-6-methyl-4H-chromen-4-one | 0.17 | ||
| 7.35 | C21H20O10 | 432 | 1,5-Anhydro-1-[5,7-dihydroxy-3-(4-hydroxyphenyl)-4-oxo-4H-chromen-8-yl]hexitol | 1.03 | ||
| 6.95 | C28H24O15 | 600 | (2S,3R,4R,5S,6S)-2-{[2-(3,4-Dihydroxyphenyl)-5,7-dihydroxy-4-oxo-4H-chromen-3-yl]oxy}-3,5-dihydroxy-6-methyloxan-4-yl 3,4,5-trihydroxybenzoate | 0.01 | ||
| Tannins | 6.09 | C14H6O8 | 303 | Ellagic acid (Isomer 1) | 3.84 | |
| 7.47 | C14H6O8 | 303 | Ellagic acid (Isomer 2) | 4.06 | ||
| 1.64 | C6H6O3 | 126 | Pyrogallol | 2.23 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zai, M.J.; Cheesman, M.J.; Cock, I.E. Phytochemical Evaluation of Terminalia canescens DC. Radlk. Extracts with Antibacterial and Antibiotic Potentiation Activities against Selected β-Lactam Drug-Resistant Bacteria. Molecules 2024, 29, 1385. https://doi.org/10.3390/molecules29061385
Zai MJ, Cheesman MJ, Cock IE. Phytochemical Evaluation of Terminalia canescens DC. Radlk. Extracts with Antibacterial and Antibiotic Potentiation Activities against Selected β-Lactam Drug-Resistant Bacteria. Molecules. 2024; 29(6):1385. https://doi.org/10.3390/molecules29061385
Chicago/Turabian StyleZai, Muhammad Jawad, Matthew James Cheesman, and Ian Edwin Cock. 2024. "Phytochemical Evaluation of Terminalia canescens DC. Radlk. Extracts with Antibacterial and Antibiotic Potentiation Activities against Selected β-Lactam Drug-Resistant Bacteria" Molecules 29, no. 6: 1385. https://doi.org/10.3390/molecules29061385