
6-Shogaol Ameliorates Liver Inflammation and Fibrosis in Mice on a Methionine- and Choline-Deficient Diet by Inhibiting Oxidative Stress, Cell Death, and Endoplasmic Reticulum Stress

Abstract
:1. Introduction
2. Results
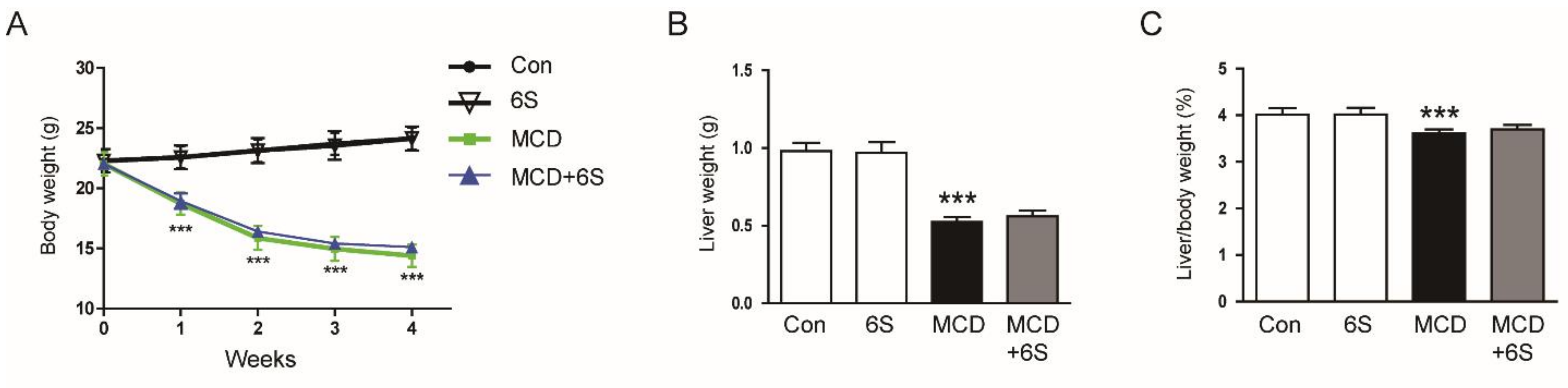
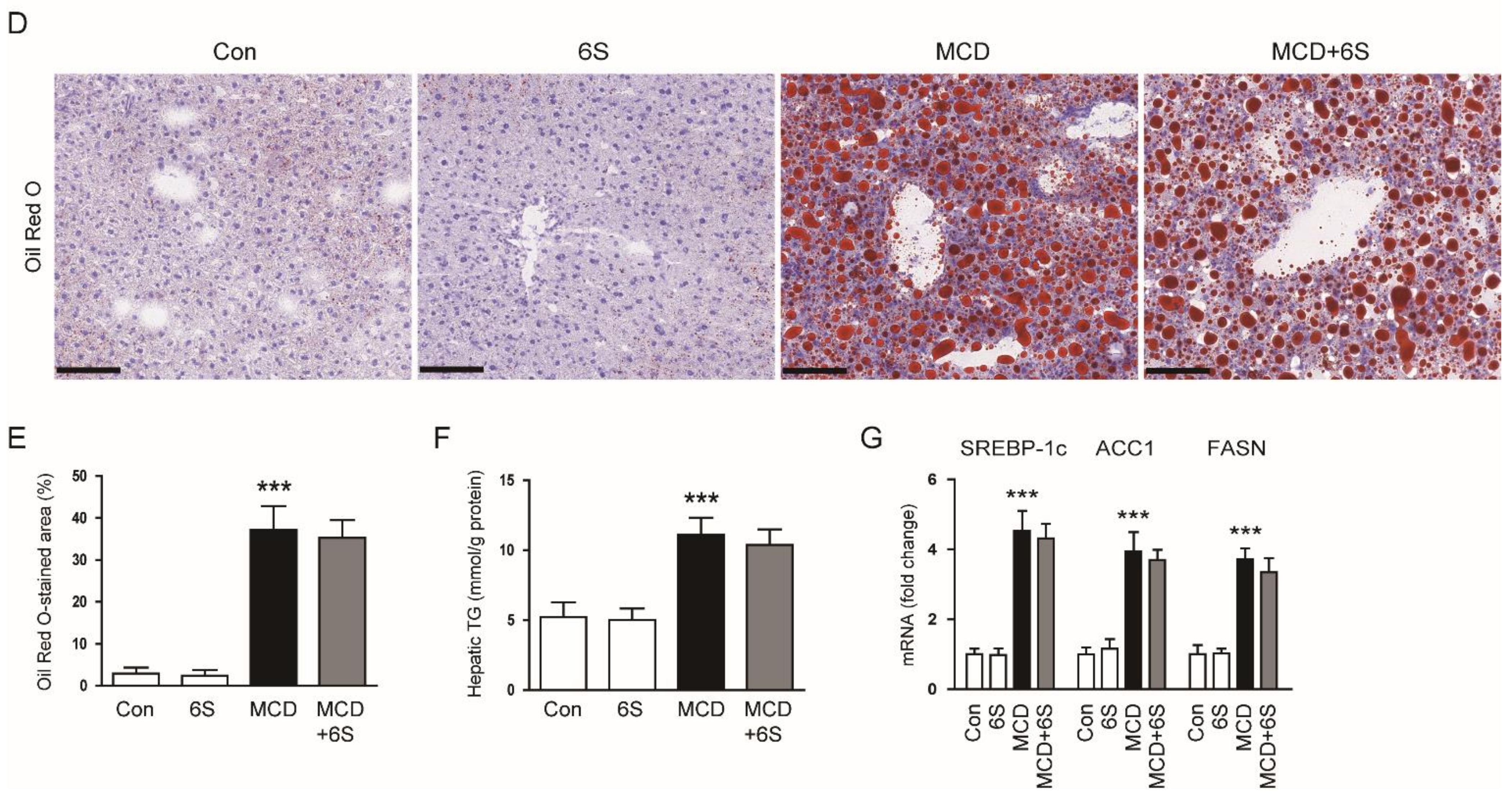
2.1. 6-Shogaol Does Not Change Body Weight, Liver Weight, and Hepatic Lipid Accumulation in MCD Diet-Fed Mice
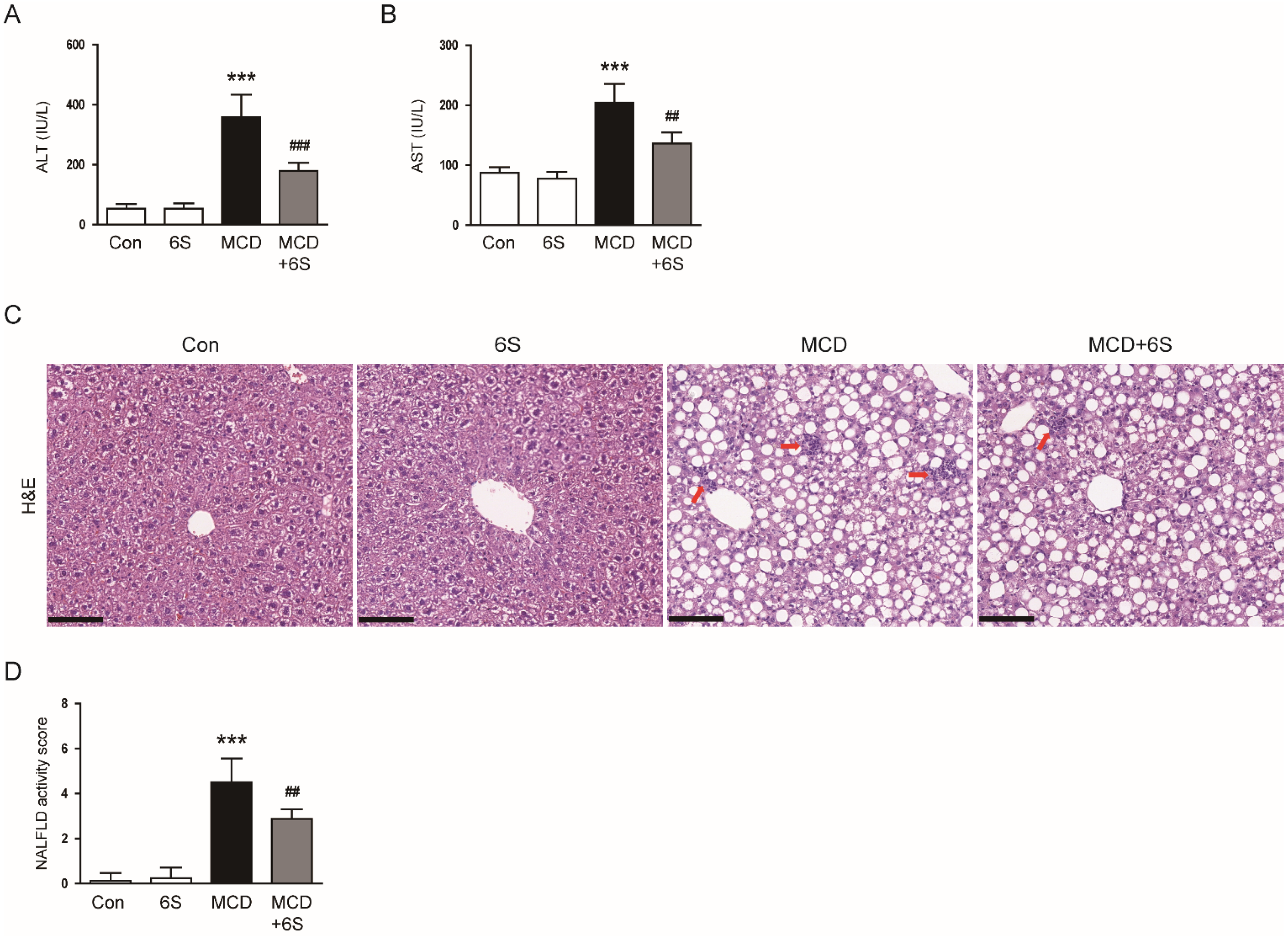
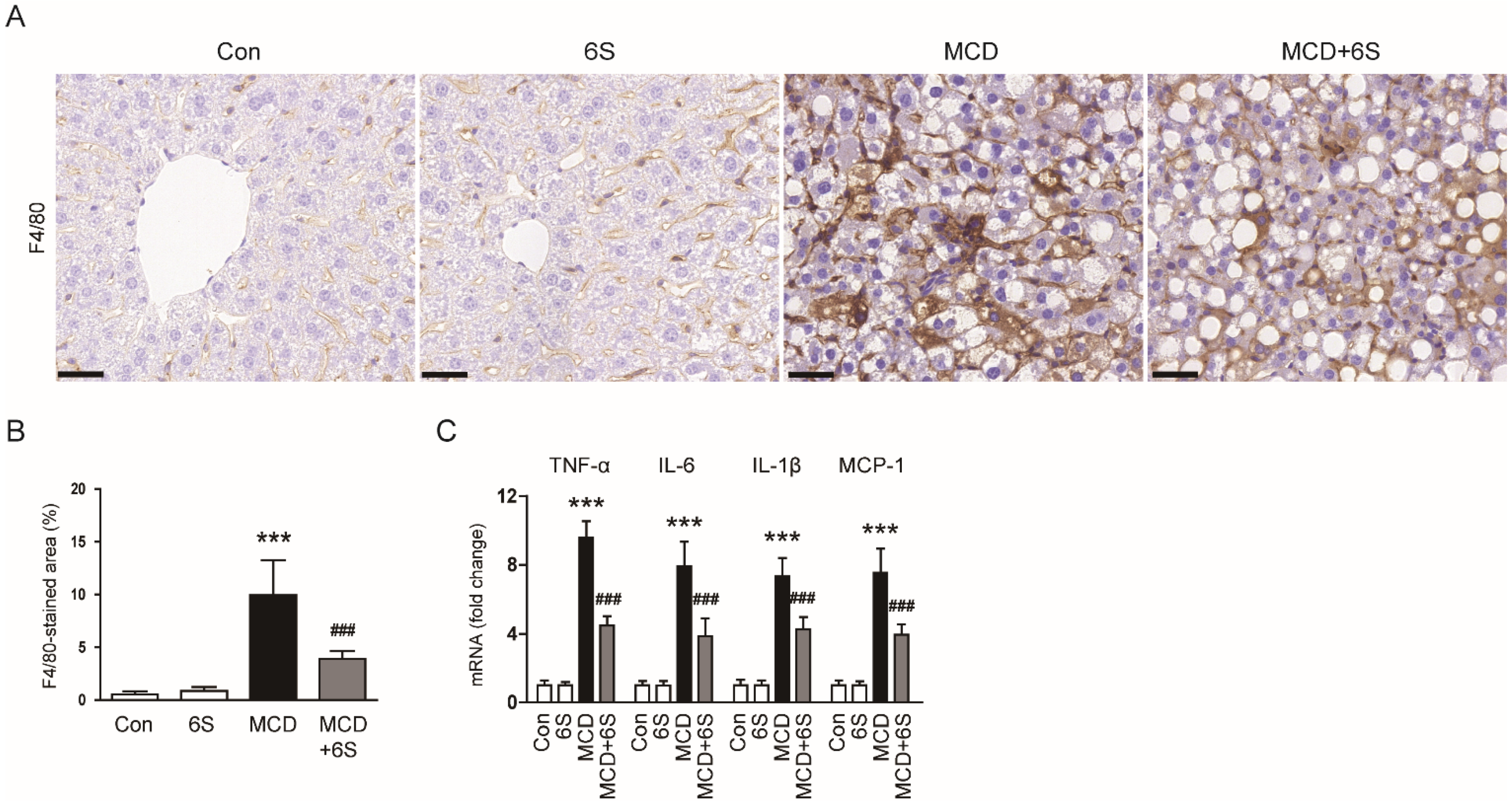
2.2. 6-Shogaol Mitigates Liver Injury and Inflammation in MCD-Diet-Fed Mice
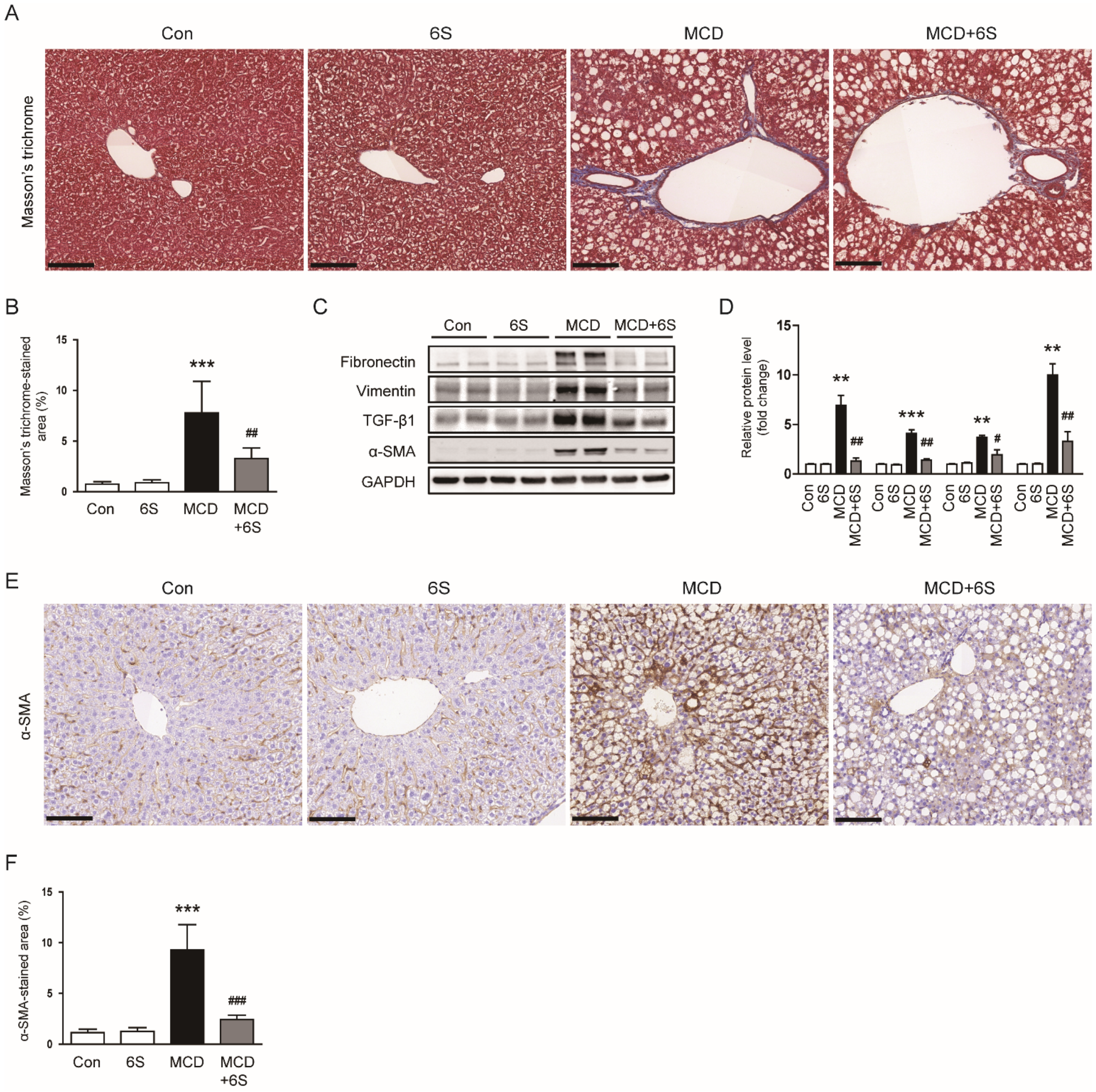
2.3. 6-Shogaol Mitigates Liver Fibrosis in MCD-Diet-Fed Mice
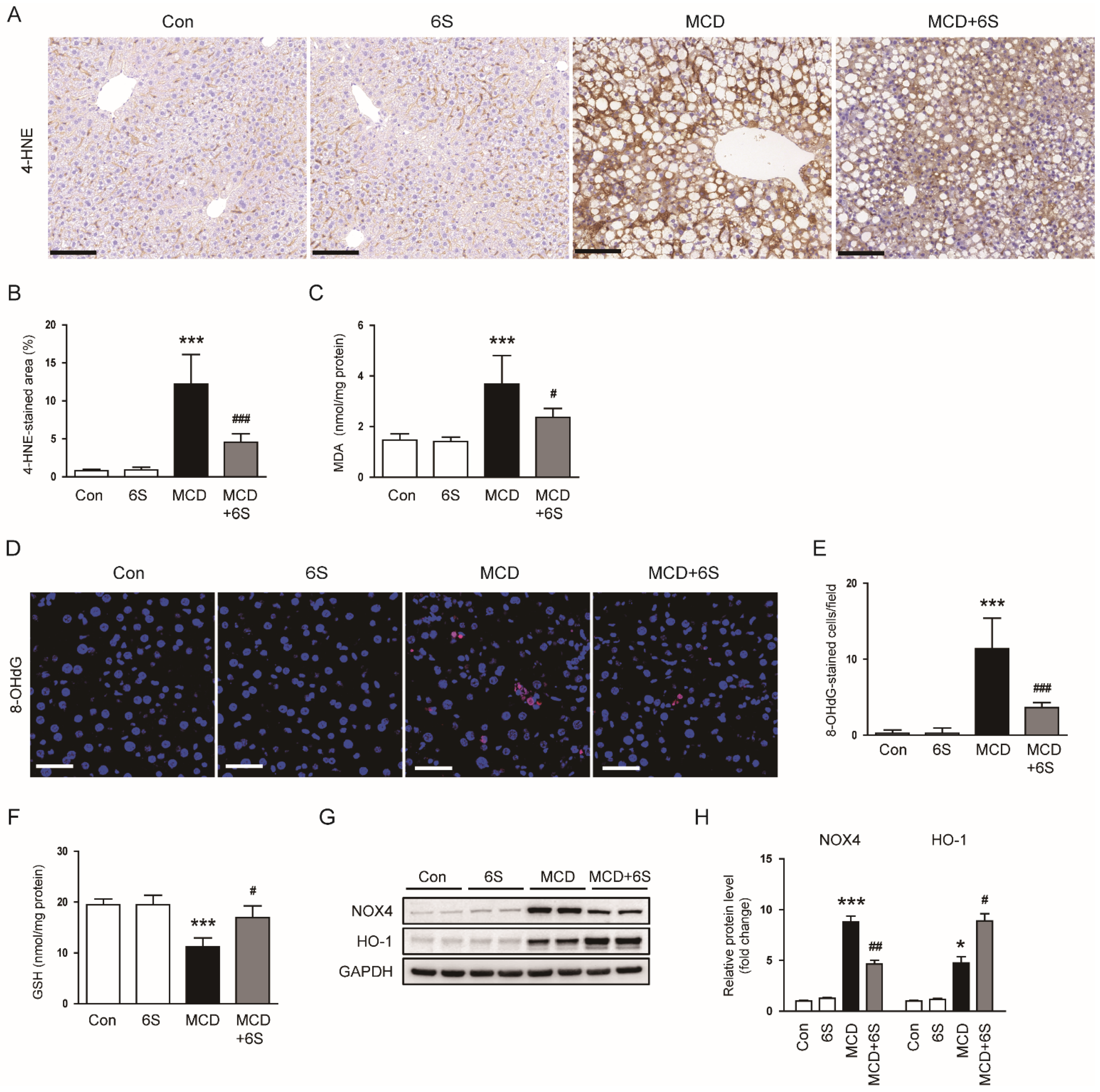
2.4. 6-Shogaol Reduces Oxidative Stress in MCD-Diet-Fed Mice
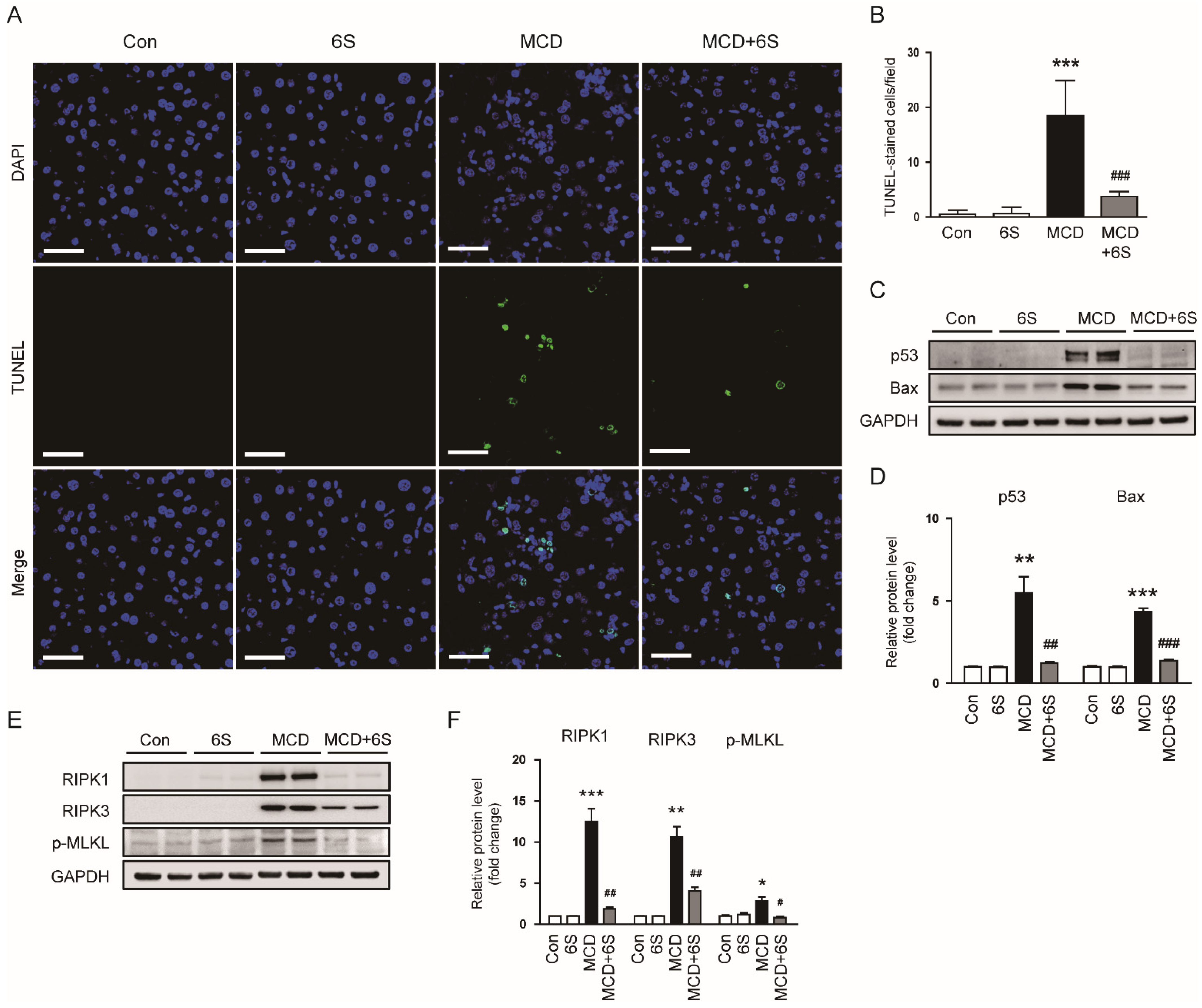
2.5. 6-Shogaol Inhibits Apoptosis and Necroptosis in MCD-Diet-Fed Mice
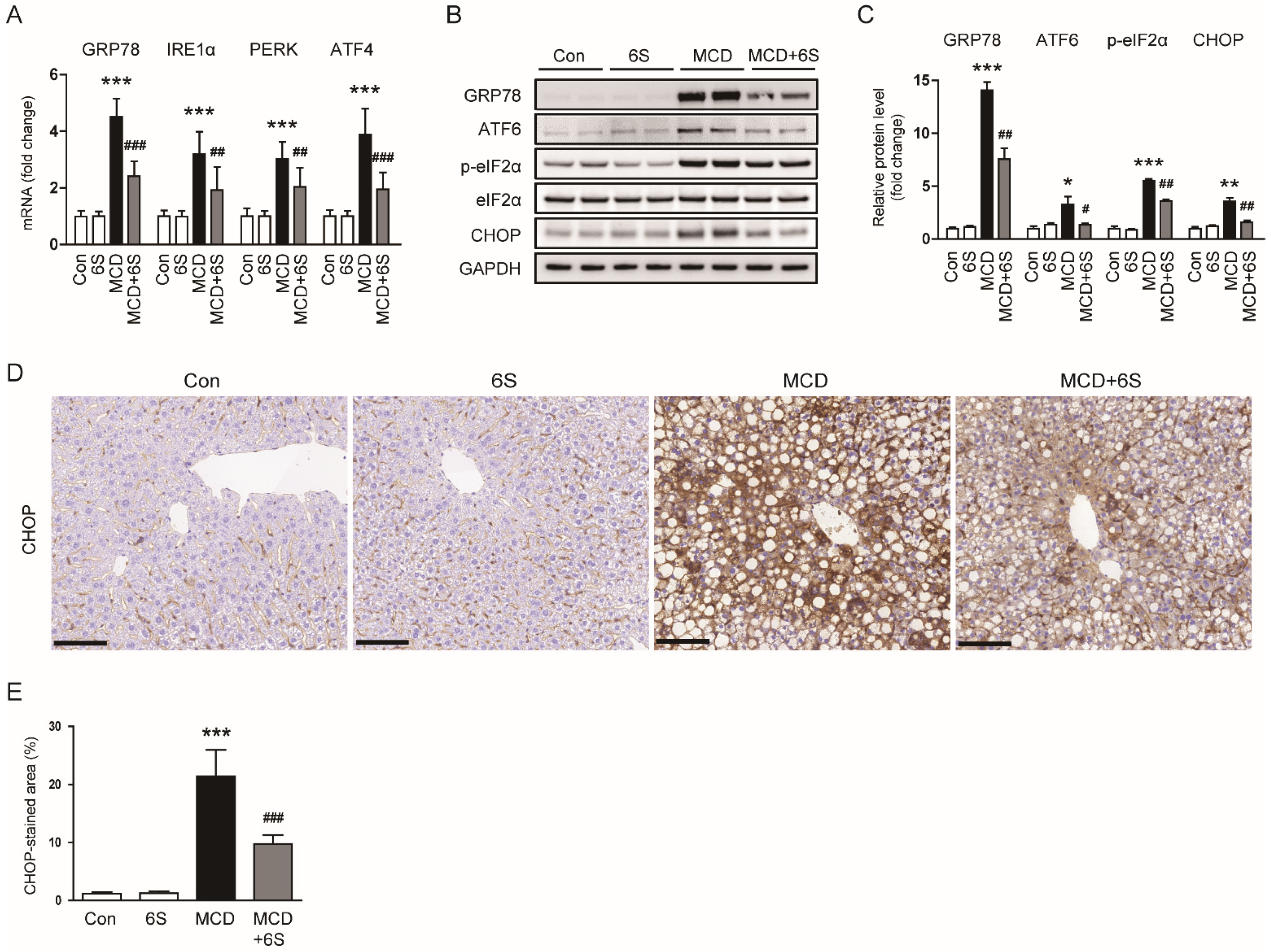
2.6. 6-Shogaol Suppresses ER Stress in MCD-Diet-Fed Mice
3. Discussion
4. Materials and Methods
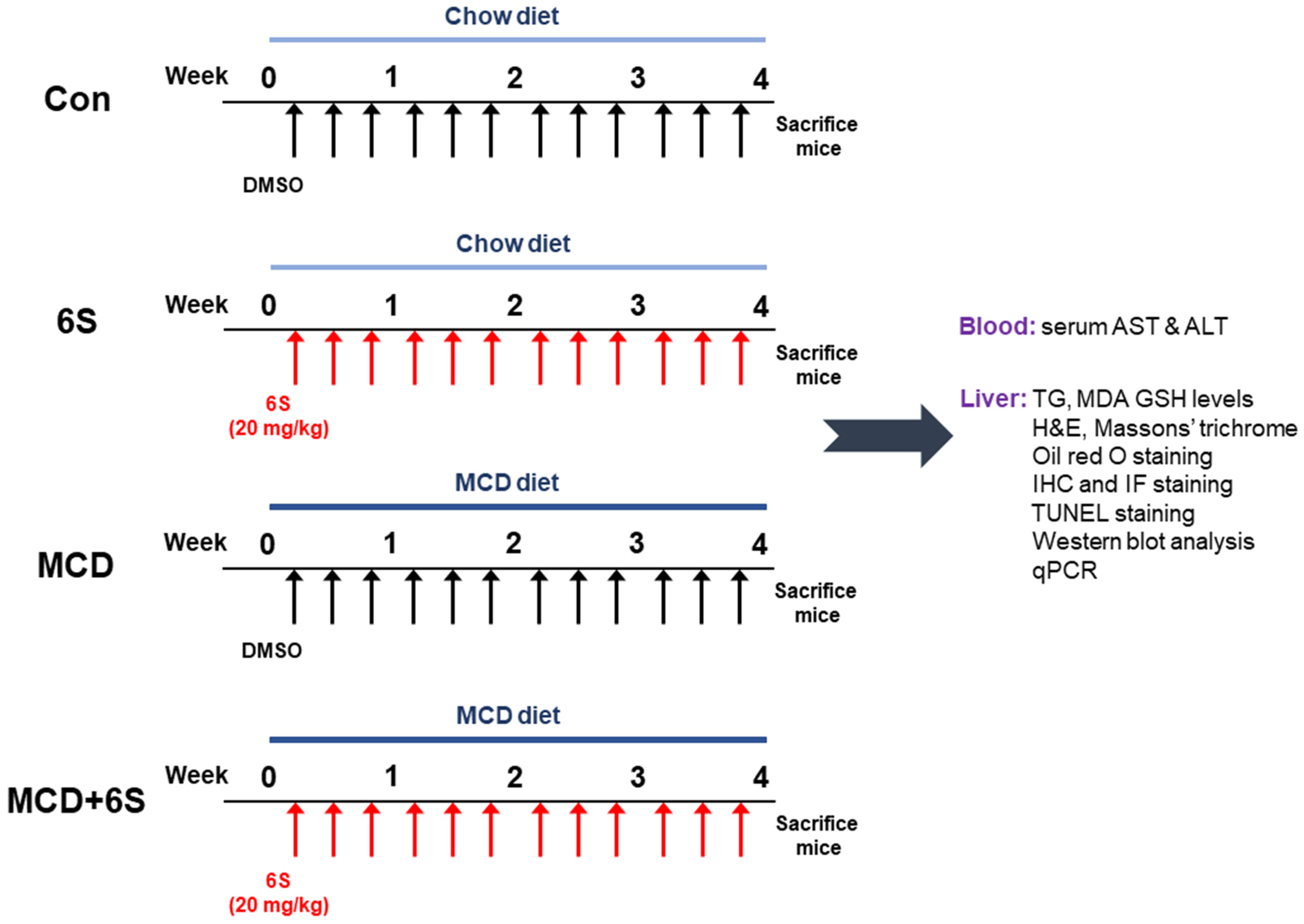
4.1. Animal Experiments
4.2. Biochemical Analysis
4.3. H&E, Masson’s Trichrome, and Oil Red O Staining
4.4. IHC and IF Staining
4.5. TUNEL Staining
4.6. Western Blot Analysis
4.7. qPCR
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Younossi, Z.M. Non-alcoholic fatty liver disease—A global public health perspective. J. Hepatol. 2019, 70, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Fazel, Y.; Koenig, A.B.; Sayiner, M.; Goodman, Z.D.; Younossi, Z.M. Epidemiology and natural history of non-alcoholic fatty liver disease. Metabolism 2016, 65, 1017–1025. [Google Scholar] [CrossRef]
- Guo, X.; Yin, X.; Liu, Z.; Wang, J. Non-Alcoholic Fatty Liver Disease (NAFLD) Pathogenesis and Natural Products for Prevention and Treatment. Int. J. Mol. Sci. 2022, 23, 15489. [Google Scholar] [CrossRef] [PubMed]
- Arroyave-Ospina, J.C.; Wu, Z.; Geng, Y.; Moshage, H. Role of Oxidative Stress in the Pathogenesis of Non-Alcoholic Fatty Liver Disease: Implications for Prevention and Therapy. Antioxidants 2021, 10, 174. [Google Scholar] [CrossRef]
- Shojaie, L.; Iorga, A.; Dara, L. Cell Death in Liver Diseases: A Review. Int. J. Mol. Sci. 2020, 21, 9682. [Google Scholar] [CrossRef] [PubMed]
- Lebeaupin, C.; Vallée, D.; Hazari, Y.; Hetz, C.; Chevet, E.; Bailly-Maitre, B. Endoplasmic reticulum stress signaling and the pathogenesis of non-alcoholic fatty liver disease. J. Hepatol. 2018, 69, 927–947. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Li, X.; Meng, X.; Lei, J.; Xia, Y.; Yu, L. Anticancer perspective of 6-shogaol: Anticancer properties, mechanism of action, synergism and delivery system. Chin. Med. 2023, 18, 138. [Google Scholar] [CrossRef] [PubMed]
- Bischoff-Kont, I.; Fürst, R. Benefits of Ginger and Its Constituent 6-Shogaol in Inhibiting Inflammatory Processes. Pharmaceuticals 2021, 14, 571. [Google Scholar] [CrossRef]
- Arcusa, R.; Villaño, D.; Marhuenda, J.; Cano, M.; Cerdà, B.; Zafrilla, P. Potential Role of Ginger (Zingiber officinale Roscoe) in the Prevention of Neurodegenerative Diseases. Front. Nutr. 2022, 9, 809621. [Google Scholar] [CrossRef]
- Gwon, M.-G.; Gu, H.; Leem, J.; Park, K.-K. Protective Effects of 6-Shogaol, an Active Compound of Ginger, in a Murine Model of Cisplatin-Induced Acute Kidney Injury. Molecules 2021, 26, 5931. [Google Scholar] [CrossRef]
- Oğuz Cumaoğlu, M.; Cumaoğlu, B.; Tekin, Y.; Günay, N. Therapeutic effect of 6-shogaol on acetaminophen-induced hepatotoxicity in mice: An experimental study. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 7371–7378. [Google Scholar] [PubMed]
- Guo, X.; Qiu, J.; Qian, Y. 6-Shogaol Mitigates Sepsis-Associated Hepatic Injury through Transcriptional Regulation. Nutrients 2021, 13, 3427. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.L.; Chai, Y.N.; Duan, F.Y.; Zhang, H.J.; Han, X.Y.; Chen, L.Y.; Duan, F. 6-Shogaol alleviates CCl4-induced liver fibrosis by attenuating inflammatory response in mice through the NF-κB pathway. Acta Biochim. Pol. 2022, 69, 363–370. [Google Scholar] [CrossRef]
- Haczeyni, F.; Yeh, M.M.; Ioannou, G.N.; Leclercq, I.A.; Goldin, R.; Dan, Y.Y.; Yu, J.; Teoh, N.C.; Farrell, G.C. Mouse models of non-alcoholic steatohepatitis: A reflection on recent literature. J. Gastroenterol. Hepatol. 2018, 33, 1312–1320. [Google Scholar] [CrossRef]
- Latorre, J.; Díaz-Trelles, R.; Comas, F.; Gavaldà-Navarro, A.; Milbank, E.; Dragano, N.; Morón-Ros, S.; Mukthavaram, R.; Ortega, F.; Castells-Nobau, A.; et al. Downregulation of hepatic lipopolysaccharide binding protein improves lipogenesis-induced liver lipid accumulation. Mol. Ther. Nucleic Acids 2022, 29, 599–613. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Oh, J.Y.; Kim, S.H.; Oh, I.J.; Lee, Y.-h.; Lee, K.W.; Lee, W.H.; Kim, J.-H. Pharmaceutical Efficacy of Gypenoside LXXV on Non-Alcoholic Steatohepatitis (NASH). Biomolecules 2020, 10, 1426. [Google Scholar] [CrossRef]
- Jang, H.-J.; Leem, J.; Kim, G.M. Protective Effects of Apamin on Acetaminophen-Induced Hepatotoxicity in Mice. Curr. Issues Mol. Biol. 2023, 45, 4389–4399. [Google Scholar] [CrossRef]
- Tan, J.; Zhang, J.; Wang, M.; Wang, Y.; Dong, M.; Ma, X.; Sun, B.; Liu, S.; Zhao, Z.; Chen, L.; et al. DRAM1 increases the secretion of PKM2-enriched EVs from hepatocytes to promote macrophage activation and disease progression in ALD. Mol. Ther. Nucleic Acids 2021, 27, 375–389. [Google Scholar] [CrossRef]
- Park, J.-H.; Leem, J.; Lee, S.-J. Protective Effects of Carnosol on Renal Interstitial Fibrosis in a Murine Model of Unilateral Ureteral Obstruction. Antioxidants 2022, 11, 2341. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Choi, Y.; Leem, J.; Song, J.E. Heme Oxygenase-1 Induction by Cobalt Protoporphyrin Ameliorates Cholestatic Liver Disease in a Xenobiotic-Induced Murine Model. Int. J. Mol. Sci. 2021, 22, 8253. [Google Scholar] [CrossRef]
- Ma, Y.; Chang, N.; Liu, Y.; Liu, F.; Dong, C.; Hou, L.; Qi, C.; Yang, L.; Li, L. Silencing IQGAP1 alleviates hepatic fibrogenesis via blocking bone marrow mesenchymal stromal cell recruitment to fibrotic liver. Mol. Ther. Nucleic Acids 2021, 27, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.; Kim, J.-Y.; Leem, J. Protective Effects of Orexin A in a Murine Model of Cisplatin-Induced Acute Kidney Injury. J. Clin. Med. 2022, 11, 7196. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-M.; Lian, C.-F.; Sun, Q.-W.; Wang, T.-T.; Liu, Y.-Y.; Ye, J.; Gao, L.-L.; Yang, Y.-F.; Liu, S.-N.; Shen, Z.-F.; et al. Ramulus Mori (Sangzhi) Alkaloids Alleviate High-Fat Diet-Induced Obesity and Nonalcoholic Fatty Liver Disease in Mice. Antioxidants 2022, 11, 905. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.A.; Shin, M.-R.; Choi, J.; Kim, M.; Park, H.-J.; Roh, S.-S. Co-Treatments of Gardeniae Fructus and Silymarin Ameliorates Excessive Oxidative Stress-Driven Liver Fibrosis by Regulation of Hepatic Sirtuin1 Activities Using Thioacetamide-Induced Mice Model. Antioxidants 2023, 12, 97. [Google Scholar] [CrossRef]
- Schwabe, R.F.; Luedde, T. Apoptosis and necroptosis in the liver: A matter of life and death. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 738–752. [Google Scholar] [CrossRef] [PubMed]
- Ajoolabady, A.; Kaplowitz, N.; Lebeaupin, C.; Kroemer, G.; Kaufman, R.J.; Malhi, H.; Ren, J. Endoplasmic reticulum stress in liver diseases. Hepatology 2023, 77, 619–639. [Google Scholar] [CrossRef] [PubMed]
- Wong, V.W.; Ekstedt, M.; Wong, G.L.; Hagström, H. Changing epidemiology, global trends and implications for outcomes of NAFLD. J. Hepatol. 2023, 79, 842–852. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; International Natural Product Sciences Taskforce; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Najmi, A.; Javed, S.A.; Al Bratty, M.; Alhazmi, H.A. Modern Approaches in the Discovery and Development of Plant-Based Natural Products and Their Analogues as Potential Therapeutic Agents. Molecules 2022, 27, 349. [Google Scholar] [CrossRef]
- Salvoza, N.; Giraudi, P.J.; Tiribelli, C.; Rosso, N. Natural Compounds for Counteracting Nonalcoholic Fatty Liver Disease (NAFLD): Advantages and Limitations of the Suggested Candidates. Int. J. Mol. Sci. 2022, 23, 2764. [Google Scholar] [CrossRef]
- Stephenson, K.; Kennedy, L.; Hargrove, L.; Demieville, J.; Thomson, J.; Alpini, G.; Francis, H. Updates on Dietary Models of Nonalcoholic Fatty Liver Disease: Current Studies and Insights. Gene Expr. 2018, 18, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Deng, X.; Bai, T.; Wang, S.; Jiang, Q.; Xu, K. Resolvin D1 mitigates non-alcoholic steatohepatitis by suppressing the TLR4-MyD88-mediated NF-κB and MAPK pathways and activating the Nrf2 pathway in mice. Int. Immunopharmacol. 2020, 88, 106961. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.V.; Michelotti, G.A.; Xie, G.; Pereira, T.A.; Boursier, J.; Bohnic, B.; Gyu, C.D.; Diehl, A.M. Mouse models of diet-induced nonalcoholic steatohepatitis reproduce the heterogeneity of the human disease. PLoS ONE 2015, 10, e0127991. [Google Scholar] [CrossRef] [PubMed]
- Ferré, P.; Phan, F.; Foufelle, F. SREBP-1c and lipogenesis in the liver: An update1. Biochem. J. 2021, 478, 3723–3739. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-C.; Wu, K.-L.; Yen, C.-K.; Chen, C.-N.; Chang, S.-F.; Huang, W.-S. 6-Shogaol Antagonizes the Adipocyte-Conditioned Medium-Initiated 5-Fluorouracil Resistance in Human Colorectal Cancer Cells through Controlling the SREBP-1 Level. Life 2021, 11, 1067. [Google Scholar] [CrossRef]
- Kazankov, K.; Jørgensen, S.M.D.; Thomsen, K.L.; Møller, H.J.; Vilstrup, H.; George, J.; Schuppan, D.; Grønbæk, H. The role of macrophages in nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 145–159. [Google Scholar] [CrossRef]
- Kim, Y.G.; Kim, M.O.; Kim, S.H.; Kim, H.J.; Pokhrel, N.K.; Lee, J.H.; Lee, H.J.; Kim, J.Y.; Lee, Y. 6-Shogaol, an active ingredient of ginger, inhibits osteoclastogenesis and alveolar bone resorption in ligature-induced periodontitis in mice. J. Periodontol. 2020, 91, 809–818. [Google Scholar] [CrossRef]
- Kawase, Y.; Sunagawa, Y.; Shimizu, K.; Funamoto, M.; Hamabe-Horiike, T.; Katanasaka, Y.; Shimizu, S.; Hawke, P.; Mori, K.; Komiyama, M.; et al. 6-Shogaol, an Active Component of Ginger, Inhibits p300 Histone Acetyltransferase Activity and Attenuates the Development of Pressure-Overload-Induced Heart Failure. Nutrients 2023, 15, 2232. [Google Scholar] [CrossRef]
- Schwabe, R.F.; Tabas, I.; Pajvani, U.B. Mechanisms of Fibrosis Development in Nonalcoholic Steatohepatitis. Gastroenterology 2020, 158, 1913–1928. [Google Scholar] [CrossRef]
- Bocca, C.; Protopapa, F.; Foglia, B.; Maggiora, M.; Cannito, S.; Parola, M.; Novo, E. Hepatic Myofibroblasts: A Heterogeneous and Redox-Modulated Cell Population in Liver Fibrogenesis. Antioxidants 2022, 11, 1278. [Google Scholar] [CrossRef]
- Wang, D.; Li, J.; Luo, G.; Zhou, J.; Wang, N.; Wang, S.; Zhao, R.; Cao, X.; Ma, Y.; Liu, G.; et al. Nox4 as a novel therapeutic target for diabetic vascular complications. Redox Biol. 2023, 64, 102781. [Google Scholar] [CrossRef] [PubMed]
- Matuz-Mares, D.; Vázquez-Meza, H.; Vilchis-Landeros, M.M. NOX as a Therapeutic Target in Liver Disease. Antioxidants 2022, 11, 2038. [Google Scholar] [CrossRef] [PubMed]
- Sasson, A.; Kristoferson, E.; Batista, R.; McClung, J.A.; Abraham, N.G.; Peterson, S.J. The pivotal role of heme Oxygenase-1 in reversing the pathophysiology and systemic complications of NAFLD. Arch. Biochem. Biophys. 2021, 697, 108679. [Google Scholar] [CrossRef] [PubMed]
- Ryter, S.W. Heme Oxygenase-1: An Anti-Inflammatory Effector in Cardiovascular, Lung, and Related Metabolic Disorders. Antioxidants 2022, 11, 555. [Google Scholar] [CrossRef] [PubMed]
- Han, S.J.; Kim, M.; D’Agati, V.D.; Lee, H.T. 6-Shogaol protects against ischemic acute kidney injury by modulating NF-κB and heme oxygenase-1 pathways. Am. J. Physiol. Renal Physiol. 2019, 317, F743–F756. [Google Scholar] [CrossRef]
- Yang, L.; Yang, F.; Teng, L.; Katayama, I. 6-Shogaol Protects Human Melanocytes against Oxidative Stress through Activation of the Nrf2-Antioxidant Response Element Signaling Pathway. Int. J. Mol. Sci. 2020, 21, 3537. [Google Scholar] [CrossRef] [PubMed]
- Weinlich, R.; Oberst, A.; Beere, H.M.; Green, D.R. Necroptosis in development, inflammation and disease. Nat. Rev. Mol. Cell Biol. 2017, 18, 127–136. [Google Scholar] [CrossRef]
- Liu, J.; Li, B.; Li, W.; Pan, T.; Diao, Y.; Wang, F. 6-Shogaol Inhibits Oxidative Stress-Induced Rat Vascular Smooth Muscle Cell Apoptosis by Regulating OXR1-p53 Axis. Front. Mol. Biosci. 2022, 9, 808162. [Google Scholar] [CrossRef]
- Riaz, T.A.; Junjappa, R.P.; Handigund, M.; Ferdous, J.; Kim, H.-R.; Chae, H.-J. Role of Endoplasmic Reticulum Stress Sensor IRE1α in Cellular Physiology, Calcium, ROS Signaling, and Metaflammation. Cells 2020, 9, 1160. [Google Scholar] [CrossRef]
- Görlach, A.; Bertram, K.; Hudecova, S.; Krizanova, O. Calcium and ROS: A mutual interplay. Redox Biol. 2015, 6, 260–271. [Google Scholar] [CrossRef]
- Cirone, M. ER Stress, UPR Activation and the Inflammatory Response to Viral Infection. Viruses 2021, 13, 798. [Google Scholar] [CrossRef] [PubMed]
- Duvigneau, J.C.; Luís, A.; Gorman, A.M.; Samali, A.; Kaltenecker, D.; Moriggl, R.; Kozlov, A.V. Crosstalk between inflammatory mediators and endoplasmic reticulum stress in liver diseases. Cytokine 2019, 124, 154577. [Google Scholar] [CrossRef] [PubMed]
- Willy, J.A.; Young, S.K.; Stevens, J.L.; Masuoka, H.C.; Wek, R.C. CHOP links endoplasmic reticulum stress to NF-κB activation in the pathogenesis of nonalcoholic steatohepatitis. Mol. Biol. Cell 2015, 26, 2190–2204. [Google Scholar] [CrossRef] [PubMed]
- Calfon, M.; Zeng, H.; Urano, F.; Till, J.H.; Hubbard, S.R.; Harding, H.P.; Clark, S.G.; Ron, D. IRE1 couples endoplasmic reticulum load to secretory capacity by processing the XBP-1 mRNA. Nature 2002, 415, 92–96. [Google Scholar] [CrossRef]
- Maurel, M.; Chevet, E.; Tavernier, J.; Gerlo, S. Getting RIDD of RNA: IRE1 in cell fate regulation. Trends Biochem. Sci. 2014, 39, 245–254. [Google Scholar] [CrossRef]
- Kim, S.H.; Lim, Y.; Park, J.B.; Kwak, J.-H.; Kim, K.-J.; Kim, J.-H.; Song, H.; Cho, J.-Y.; Hwang, D.Y.; Kim, K.S.; et al. Comparative study of fatty liver induced by methionine and choline-deficiency in C57BL/6N mice originating from three different sources. Lab. Anim. Res. 2017, 33, 157–164. [Google Scholar] [CrossRef]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Nonalcoholic Steatohepatitis Clinical Research Network. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′→3′) | Accession No. |
|---|---|---|
| SREBP-1c | F: ACGGAGCCATGGATTGCACA R: AAGGGTGCAGGTGTCACCTT | NM_001358314 |
| ACC1 | F: GAATCTCCTGGTGACAATGCTTATT R: GGTCTTGCTGAGTTGGGTTAGCT | NM_133360 |
| FASN | F: CTGAGATCCCAGCACTTCTTGA R: GCCTCCGAAGCCAAATGAG | NM_007988 |
| TNF-α | F: ACTTCGGGGTGATCGGTCCCC R: TGGTTTGCTACGACGTGGGCTAC | NM_013693 |
| IL-6 | F: TACCACTTCACAAGTCGGAGGC R: CTGCAAGTGCATCATCGTTGTTC | NM_031168 |
| IL-1β | F: TGCAGCTGGAGAGTGTGGATCCC R: TGTGCTCTGCTTGTGAGGTGCTG | NM_008361 |
| MCP-1 | F: GGGCCTGCTGTTCACAGTT R: CCAGCCTACTCATTGGGAT | NM_011333 |
| GRP78 | F: TGGTATTCTCCGAGTGACAGC R: AGTCTTCAATGTCCGCATCC | NM_001163434 |
| IRE1α | F: GCATCACCAAGTGGAAGTATC R: ACCATTGAGGGAGAGGCATAG | NM_023913 |
| PERK | F: AGCACTCAGATGGAGAGAGTCAG R: GCTATGGGAGTTGTTGGACTGT | NM_004836 |
| ATF4 | F: GAGCTTCCTGAACAGCGAAGTG R: TGGCCACCTCCAGATAGTCATC | NM_009716 |
| GAPDH | F: ACTCCACTCACGGCAAATTC R: TCTCCATGGTGGTGAAGACA | NM_001289726 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, A.Y.; Kim, K.; Kwon, H.H.; Leem, J.; Song, J.E. 6-Shogaol Ameliorates Liver Inflammation and Fibrosis in Mice on a Methionine- and Choline-Deficient Diet by Inhibiting Oxidative Stress, Cell Death, and Endoplasmic Reticulum Stress. Molecules 2024, 29, 419. https://doi.org/10.3390/molecules29020419
Yang AY, Kim K, Kwon HH, Leem J, Song JE. 6-Shogaol Ameliorates Liver Inflammation and Fibrosis in Mice on a Methionine- and Choline-Deficient Diet by Inhibiting Oxidative Stress, Cell Death, and Endoplasmic Reticulum Stress. Molecules. 2024; 29(2):419. https://doi.org/10.3390/molecules29020419
Chicago/Turabian StyleYang, Ah Young, Kiryeong Kim, Hyun Hee Kwon, Jaechan Leem, and Jeong Eun Song. 2024. "6-Shogaol Ameliorates Liver Inflammation and Fibrosis in Mice on a Methionine- and Choline-Deficient Diet by Inhibiting Oxidative Stress, Cell Death, and Endoplasmic Reticulum Stress" Molecules 29, no. 2: 419. https://doi.org/10.3390/molecules29020419