
In Silico and In Vitro Identification of 1,8-Dihydroxy-4,5-dinitroanthraquinone as a New Antibacterial Agent against Staphylococcus aureus and Enterococcus faecalis

Abstract
:1. Introduction
2. Results
2.1. Virtual Screenings Based on Molecular Docking and Pharmacophore Model
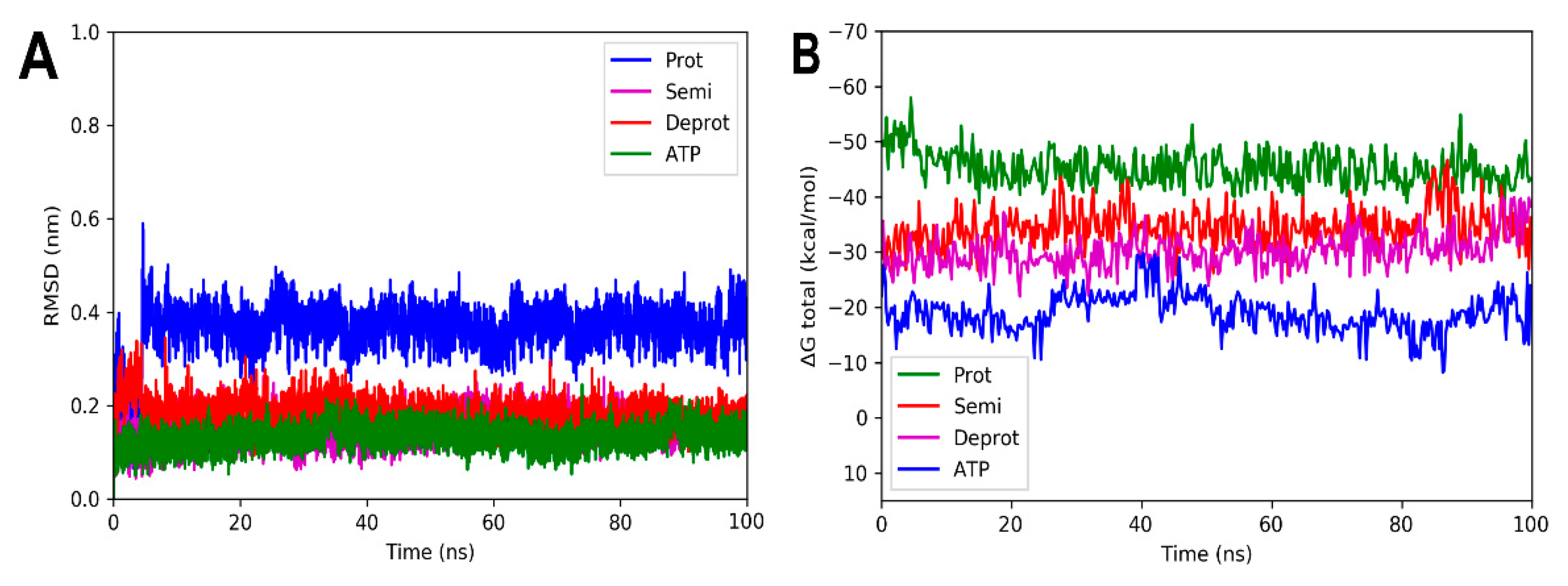
2.2. Molecular Dynamic Simulations and Total Binding Free Energy Calculations
2.3. Decomposition of Binding Free Energy Analysis
2.4. Pharmacokinetic and Target Fishing Predictions
2.5. In Vitro Evaluation of the Antibacterial Activity of DHDNA
2.6. Evaluation of the Potential of DHDNA to Resensitize Antibiotic-Resistant Bacteria
2.7. Determination of Minimum Inhibitory Concentrations
2.8. Time-Kill Kinetic Analysis
3. Materials and Methods
3.1. Ligands and Targets Preparation for In Silico Analyses
3.2. Molecular Docking Analyses
3.3. Pharmacophore-Based Virtual Screening
3.4. Molecular Dynamic Simulations
3.5. Binding Free Energy Calculation
3.6. Pharmacokinetic, Target Fishing, and Cytotoxic Predictions
3.7. Material
3.8. Isolation and Identification of Bacteria
3.9. Antibacterial Activity and Minimal Inhibitory Concentration Assays
3.10. Evaluation of the Sensitizing Potential of DHDNA in Antibiotic-Resistant Bacteria
3.11. Time-Kill Kinetic Assay
3.12. Data Analysis and Visualizations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial Resistance: A Global Multifaceted Phenomenon. Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Hawkey, P.M.; Jones, A.M. The Changing Epidemiology of Resistance. J. Antimicrob. Chemother. 2009, 64, i3–i10. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Ahmed, M.Z.; Rafique, S.; Almasoudi, S.E.; Shah, M.; Jalil, N.A.C.; Ojha, S.C. Recent Approaches for Downplaying Antibiotic Resistance: Molecular Mechanisms. Biomed. Res. Int. 2023, 2023, 5250040. [Google Scholar] [CrossRef] [PubMed]
- Vivas, R.; Barbosa, A.A.T.; Dolabela, S.S.; Jain, S. Multidrug-Resistant Bacteria and Alternative Methods to Control Them: An Overview. Microb. Drug Resist. 2019, 25, 890–908. [Google Scholar] [CrossRef] [PubMed]
- Amorim, J.C.; Carpio, J.M. Alpha-Naphthoflavone as a Novel Scaffold for the Design of Potential Inhibitors of the APH(3′)-IIIa Nucleotide-Binding Site of Enterococcus Faecalis. Microorganisms 2023, 11, 2351. [Google Scholar] [CrossRef] [PubMed]
- Lambert, P.A. Bacterial Resistance to Antibiotics: Modified Target Sites. Adv. Drug Deliv. Rev. 2005, 57, 1471–1485. [Google Scholar] [CrossRef] [PubMed]
- Kumawat, M.; Nabi, B.; Daswani, M.; Viquar, I.; Pal, N.; Sharma, P.; Tiwari, S.; Devojit Kumar Sarma, S.S.; Kumar, M.; Kumawat, M.; et al. Role of Bacterial Efflux Pump Proteins in Antibiotic Resistance across Microbial Species. Microb. Pathog. 2023, 181, 106182. [Google Scholar] [CrossRef]
- Ghai, I.; Ghai, S. Understanding Antibiotic Resistance via Outer Membrane Permeability. Infect. Drug Resist. 2018, 11, 523–530. [Google Scholar] [CrossRef]
- Kim, D.W.; Thawng, C.N.; Choi, J.H.; Lee, K.; Cha, C.J. Polymorphism of Antibiotic-Inactivating Enzyme Driven by Ecology Expands the Environmental Resistome. ISME J. 2018, 12, 267–276. [Google Scholar] [CrossRef]
- Miller, J.R.; Ohren, J.; Sarver, R.W.; Mueller, W.T.; De Dreu, P.; Case, H.; Thanabal, V. Phosphopantetheine Adenylyltransferase from Escherichia Coli: Investigation of the Kinetic Mechanism and Role in Regulation of Coenzyme A Biosynthesis. J. Bacteriol. 2007, 189, 8196–8205. [Google Scholar] [CrossRef]
- Kim, K.H.; Lopez-Casillas, F.; Bai, D.H.; Luo, X.; Pape, M.E. Role of Reversible Phosphorylation of Acetyl-CoA Carboxylase in Long-Chain Fatty Acid Synthesis. FASEB J. 1989, 3, 2250–2256. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, R.; Zhang, Y.-M.; Charles, O.; Rock, S.J. Coenzyme A: Back in Action. Prog. Lipid Res. 2005, 44, 125–153. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Sharma, P.; Singh, T.P.; Sharma, S. Phosphopantetheine Adenylyltransferase: A Promising Drug Target to Combat Antibiotic Resistance. Biochim. Biophys. Acta–Proteins Proteom. 2021, 1869, 140566. [Google Scholar] [CrossRef] [PubMed]
- De Jonge, B.L.M.; Walkup, G.K.; Lahiri, S.D.; Huynh, H.; Neckermann, G.; Utley, L.; Nash, T.J.; Brock, J.; San Martin, M.; Kutschke, A.; et al. Discovery of Inhibitors of 4′-Phosphopantetheine Adenylyltransferase (PPAT) to Validate PPAT as a Target for Antibacterial Therapy. Antimicrob. Agents Chemother. 2013, 57, 6005–6015. [Google Scholar] [CrossRef] [PubMed]
- Malik, E.M.; Christa, E. Müller Anthraquinones As Pharmacological Tools and Drugs. Med. Res. Rev. 2016, 36, 705–748. [Google Scholar] [CrossRef] [PubMed]
- Malik, M.S.; Alsantali, R.I.; Jassas, R.S.; Alsimaree, A.A.; Syed, R.; Alsharif, M.A.; Kalpana, K.; Morad, M.; Althagafi, I.I.; Ahmed, S.A. Journey of Anthraquinones as Anticancer Agents-a Systematic Review of Recent Literature. RSC Adv. 2021, 11, 35806–35827. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jiang, J.G. Health Functions and Structure-Activity Relationships of Natural Anthraquinones from Plants. Food Funct. 2018, 9, 6063–6080. [Google Scholar] [CrossRef]
- Yusuf, M.; Mohammad, F.; Shabbir, M. Eco-Friendly and Effective Dyeing of Wool with Anthraquinone Colorants Extracted from Rubia Cordifolia Roots: Optimization, Colorimetric and Fastness Assay, Coloring Studies with Anthraquinone Colorants Extracted from Rubia Cordifolia Roots on Wool. J. King Saud Univ.–Sci. 2017, 29, 137–144. [Google Scholar] [CrossRef]
- Zhang, M.-M.; Gong, Z.-C.; Zhao, Q.; Xu, D.-Q.; Fu, R.-J.; Tang, Y.-P.; Chen, Y.-Y. Time-Dependent Laxative Effect of Sennoside A, the Core Functional Component of Rhubarb, Is Attributed to Gut Microbiota and Aquaporins. J. Ethnopharmacol. 2023, 311, 116431. [Google Scholar] [CrossRef]
- Sayed, H.M.; Ramadan, M.A.; Salem, H.H.; Ahmad, I.; Patel, H.; Fayed, M.A.A. Phytochemical Investigation, In Silico/In Vivo Analgesic, and Anti-Inflammatory Assessment of the Egyptian Cassia occidentalis L. Steroids 2023, 196, 109245. [Google Scholar] [CrossRef]
- Kesharwani, D.; Das Paul, S.; Paliwal, R.; Satapathy, T. Exploring Potential of Diacerin Nanogel for Topical Application in Arthritis: Formulation Development, QbD Based Optimization and Pre-Clinical Evaluation. Colloids Surf. B Biointerfaces 2023, 223, 113160. [Google Scholar] [CrossRef] [PubMed]
- Arrousse, N.; Harras, M.F.; El Kadiri, S.; Haldhar, R.; Ichou, H.; Bousta, D.; Grafov, A.; Rais, Z.; Taleb, M. New Anthraquinone Drugs and Their Anticancer Activities: Cytotoxicity, DFT, Docking and ADMET Properties. Results Chem. 2023, 6, 100996. [Google Scholar] [CrossRef]
- Zhu, Y.; Yu, J.; Chen, T.; Liu, W.; Huang, Y.; Li, J.; Zhang, B.; Zhu, G.; He, Z.; Long, Y.; et al. Design, Synthesis, and Biological Evaluation of a Series of New Anthraquinone Derivatives as Anti-ZIKV Agents. Eur. J. Med. Chem. 2023, 258, 115620. [Google Scholar] [CrossRef] [PubMed]
- Alias, C.; Feretti, D.; Viola, G.V.C.; Zerbini, I.; Bisceglie, F.; Pelosi, G.; Zani, C.; Buschini, A.; Carcelli, M.; Rogolino, D.; et al. Allium Cepa Tests: A Plant-Based Tool for the Early Evaluation of Toxicity and Genotoxicity of Newly Synthetized Antifungal Molecules. Mutat. Res. Toxicol. Environ. Mutagen. 2023, 889, 503654. [Google Scholar] [CrossRef]
- Mahanty, S.; Rathinasamy, K. The Natural Anthraquinone Dye Purpurin Exerts Antibacterial Activity by Perturbing the FtsZ Assembly. Bioorg. Med. Chem. 2021, 50, 116463. [Google Scholar] [CrossRef] [PubMed]
- Carpio Arévalo, J.M.; Amorim, J.C. An In-Silico Analysis Reveals 7,7′-Bializarin as a Promising DNA Gyrase B Inhibitor on Gram-Positive and Gram-Negative Bacteria. Comput. Biol. Med. 2021, 135, 104626. [Google Scholar] [CrossRef] [PubMed]
- Amorim, J.C.; Cabrera Bermeo, A.E.; Vásquez, V.E.; Urgilés, M.R.M.; León, J.M.; Carpio, A. An Silico Evaluation of Anthraquinone Derivatives as Potential Inhibitors of DNA Gyrase B of Mycobacterium Tuberculosis. Microorganisms 2022, 10, 2434. [Google Scholar] [CrossRef]
- Duan, F.; Xin, G.; Niu, H.; Huang, W. Chlorinated Emodin as a Natural Antibacterial Agent against Drug-Resistant Bacteria through Dual Influence on Bacterial Cell Membranes and DNA. Sci. Rep. 2017, 7, 12721. [Google Scholar] [CrossRef]
- Wang, J.; Qu, Q.; Liu, X.; Cui, W.; Yu, F.; Chen, X.; Xing, X.; Zhou, Y.; Yang, Y.; Bello-Onaghise, G.; et al. 1-Hydroxyanthraquinone Exhibited Antibacterial Activity by Regulating Glutamine Synthetase of Staphylococcus Xylosus as a Virulence Factor. Biomed. Pharmacother. 2020, 123, 109779. [Google Scholar] [CrossRef]
- Lu, C.; Wang, H.; Lv, W.; Xu, P.; Zhu, J.; Xie, J.; Liu, B.; Lou, Z. Antibacterial Properties of Anthraquinones Extracted from Rhubarb against Aeromonas Hydrophila. Fish. Sci. 2011, 77, 375–384. [Google Scholar] [CrossRef]
- Boháčová, V.; Dočolomanský, P.; Breier, A.; Gemeiner, P.; Ziegelhöffer, A. Interaction of Lactate Dehydrogenase with Anthraquinone Dyes: Characterization of Ligands for Dye-Ligand Chromatography. J. Chromatogr. B Biomed. Appl. 1998, 715, 273–281. [Google Scholar] [CrossRef] [PubMed]
- De Moliner, E.; Moro, S.; Sarno, S.; Zagotto, G.; Zanotti, G.; Pinna, L.A.; Battistutta, R. Inhibition of Protein Kinase CK2 by Anthraquinone-Related Compounds: A Structural Insight. J. Biol. Chem. 2003, 278, 1831–1836. [Google Scholar] [CrossRef] [PubMed]
- Amaro, R.E.; Baudry, J.; Chodera, J.; Demir, Ö.; McCammon, J.A.; Miao, Y.; Smith, J.C. Ensemble Docking in Drug Discovery. Biophys. J. 2018, 114, 2271–2278. [Google Scholar] [CrossRef] [PubMed]
- Seidel, T.; Ibis, G.; Bendix, F.; Wolber, G. Strategies for 3D Pharmacophore-Based Virtual Screening. Drug Discov. Today Technol. 2010, 7, e221–e228. [Google Scholar] [CrossRef] [PubMed]
- Sunseri, J.; Koes, D.R. Pharmit: Interactive Exploration of Chemical Space. Nucleic Acids Res. 2016, 44, W442–W448. [Google Scholar] [CrossRef] [PubMed]
- Szymańska, M.; Majerz, I. Effect of Substitution of Hydrogen Atoms in the Molecules of Anthrone and Anthraquinone. Molecules 2021, 26, 502. [Google Scholar] [CrossRef]
- Chu, J.J.H.; Lee, R.C.H.; Ang, M.J.Y.; Wang, W.L.; Lim, H.A.; Wee, J.L.K.; Joy, J.; Hill, J.; Brian Chia, C.S. Antiviral Activities of 15 Dengue NS2B-NS3 Protease Inhibitors Using a Human Cell-Based Viral Quantification Assay. Antivir. Res. 2015, 118, 68–74. [Google Scholar] [CrossRef]
- Song, Z.M.; Zhang, J.L.; Zhou, K.; Yue, L.M.; Zhang, Y.; Wang, C.Y.; Wang, K.L.; Xu, Y. Anthraquinones as Potential Antibiofilm Agents Against Methicillin-Resistant Staphylococcus Aureus. Front. Microbiol. 2021, 12, 709826. [Google Scholar] [CrossRef]
- Celik, S.; Ozkok, F.; Ozel, A.E.; Müge Sahin, Y.; Akyuz, S.; Sigirci, B.D.; Kahraman, B.B.; Darici, H.; Karaoz, E. Synthesis, FT-IR and NMR Characterization, Antimicrobial Activity, Cytotoxicity and DNA Docking Analysis of a New Anthraquinone Derivate Compound. J. Biomol. Struct. Dyn. 2020, 38, 756–770. [Google Scholar] [CrossRef]
- Friedman, M.; Xu, A.; Lee, R.; Nguyen, D.N.; Phan, T.A.; Hamada, S.M.; Panchel, R.; Tam, C.C.; Kim, J.H.; Cheng, L.W.; et al. The Inhibitory Activity of Anthraquinones against Pathogenic Protozoa, Bacteria, and Fungi and the Relationship to Structure. Molecules 2020, 25, 3101. [Google Scholar] [CrossRef]
- Da Silva, R.A.G.; Wong, J.J.; Antypas, H.; Choo, P.Y.; Goh, K.; Jolly, S.; Liang, C.; Sing, L.T.K.; Veleba, M.; Hu, G.; et al. Mitoxantrone Targets Both Host and Bacteria to Overcome Vancomycin Resistance in Enterococcus Faecalis. Sci. Adv. 2023, 9, eadd9280. [Google Scholar] [CrossRef] [PubMed]
- Manojlovic, N.T.; Novakovic, M.; Stevovic, V.; Solujic, S. Antimicrobial Metabolites from Three Serbian Caloplaca. Pharm. Biol. 2005, 43, 718–722. [Google Scholar] [CrossRef]
- Kemegne, G.A.; Mkounga, P.; Essia Ngang, J.J.; Sado Kamdem, S.L.; Nkengfack, A.E. Antimicrobial Structure-Activity Relationship of Five Anthraquinones of Emodine Type Isolated from Vismia Laurentii. BMC Microbiol. 2017, 17, 41. [Google Scholar] [CrossRef] [PubMed]
- Peerzada, Z.; Kanhed, A.M.; Desai, K.B. Effects of Active Compounds from Cassia Fistula on Quorum Sensing Mediated Virulence and Biofilm Formation in Pseudomonas Aeruginosa. RSC Adv. 2022, 12, 15196–15214. [Google Scholar] [CrossRef] [PubMed]
- Breijyeh, Z.; Jubeh, B.; Karaman, R. Resistance of Gram-Positive Bacteria to Current Antibacterial Agents and Overcoming Approaches. Molecules 2020, 25, 2888. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Peng, W.; Qin, R.; Yan, Z.; Cen, Y.; Zheng, X.; Pan, X.; Jiang, W.; Li, B.; Li, X.; et al. The Direct Anti-MRSA Effect of Emodin via Damaging Cell Membrane. Appl. Microbiol. Biotechnol. 2015, 99, 7699–7709. [Google Scholar] [CrossRef] [PubMed]
- Alves, D.S.; Pérez-Fons, L.; Estepa, A.; Micol, V. Membrane-Related Effects Underlying the Biological Activity of the Anthraquinones Emodin and Barbaloin. Biochem. Pharmacol. 2004, 68, 549–561. [Google Scholar] [CrossRef]
- Richter, M.F.; Drown, B.S.; Riley, A.P.; Garcia, A.; Shirai, T.; Svec, R.L.; Hergenrother, P.J. Predictive Compound Accumulation Rules Yield a Broad-Spectrum Antibiotic. Nature 2017, 545, 299–304. [Google Scholar] [CrossRef]
- Richter, M.F.; Hergenrother, P.J. The Challenge of Converting Gram-Positive-Only Compounds into Broad-Spectrum Antibiotics. Ann. N. Y. Acad. Sci. 2019, 1435, 18–38. [Google Scholar] [CrossRef]
- Azelmat, J.; Larente, J.F.; Grenier, D. The Anthraquinone Rhein Exhibits Synergistic Antibacterial Activity in Association with Metronidazole or Natural Compounds and Attenuates Virulence Gene Expression in Porphyromonas Gingivalis. Arch. Oral Biol. 2015, 60, 342–346. [Google Scholar] [CrossRef]
- Machado, F.P.; Rodrigues, I.C.; Gales, L.; Pereira, J.A.; Costa, P.M.; Dethoup, T.; Mistry, S.; Silva, A.M.S.; Vasconcelos, V.; Kijjoa, A. New Alkylpyridinium Anthraquinone, Isocoumarin, C-Glucosyl Resorcinol Derivative and Prenylated Pyranoxanthones from the Culture of a Marine Sponge-Associated Fungus, Aspergillus Stellatus KUFA 2017. Mar. Drugs 2022, 20, 672. [Google Scholar] [CrossRef] [PubMed]
- Dell’Annunziata, F.; Folliero, V.; Palma, F.; Crudele, V.; Finamore, E.; Sanna, G.; Manzin, A.; De Filippis, A.; Galdiero, M.; Franci, G. Anthraquinone Rhein Exhibits Antibacterial Activity against Staphylococcus Aureus. Appl. Sci. 2022, 12, 8691. [Google Scholar] [CrossRef]
- Irwin, J.J.; Tang, K.G.; Young, J.; Dandarchuluun, C.; Wong, B.R.; Khurelbaatar, M.; Moroz, Y.S.; Mayfield, J.; Sayle, R.A. ZINC20—A Free Ultralarge-Scale Chemical Database for Ligand Discovery. J. Chem. Inf. Model. 2020, 60, 6065–6073. [Google Scholar] [CrossRef] [PubMed]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Eva Zurek, G.R.H. Avogadro: An Advanced Semantic Chemical Editor, Visualization, and Analysis Platform. J. Cheminform. 2012, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- Sander, T.; Freyss, J.; Von Korff, M.; Rufener, C. DataWarrior: An Open-Source Program for Chemistry Aware Data Visualization and Analysis. J. Chem. Inf. Model. 2015, 55, 460–473. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.J.; Kang, J.Y.; Mikami, B.; Lee, H.H.; Suh, S.W. Crystal Structure of Phosphopantetheine Adenylyltransferase from Enterococcus Faecalis in the Ligand-Unbound State and in Complex with ATP and Pantetheine. Mol. Cells 2011, 32, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Moreau, R.J.; Skepper, C.K.; Appleton, B.A.; Blechschmidt, A.; Balibar, C.J.; Benton, B.M.; Drumm, J.E., III; Feng, B.Y.; Geng, M.; Li, C.; et al. Fragment-Based Drug Discovery of Inhibitors of Phosphopantetheine Adenylyltransferase from Gram-Negative Bacteria. J. Med. Chem. 2018, 61, 3309–3324. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Korb, O.; Stützle, T.; Exner, T.E. PLANTS: Application of Ant Colony Optimization to Structure-Based Drug Design. In International Workshop on Ant Colony Optimization and Swarm Intelligence; Springer: Berlin/Heidelberg, Germany, 2006; Volume 4150, pp. 247–258. [Google Scholar] [CrossRef]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J.C. GROMACS: Fast, Flexible, and Free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef]
- Best, R.B.; Zhu, X.; Shim, J.; Lopes, P.E.M.; Mittal, J.; Feig, M.; MacKerell, A.D. Optimization of the Additive CHARMM All-Atom Protein Force Field Targeting Improved Sampling of the Backbone φ, ψ and Side-Chain Χ1 and Χ2 Dihedral Angles. J. Chem. Theory Comput. 2012, 8, 3257–3273. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; Postma, J.P.M.; Van Gunsteren, W.F.; Dinola, A.; Haak, J.R. Molecular Dynamics with Coupling to an External Bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef]
- Ewald, P.P. Die Berechnung Optischer Und Elektrostatischer Gitterpotentiale. Ann. Phys. 1921, 369, 253–287. [Google Scholar] [CrossRef]
- Hess, B.; Bekker, H.; Berendsen, H.J.C.; Fraaije, J.G.E.M. LINCS: A Linear Constraint Solver for Molecular Simulations. J. Comput. Chem. 1997, 18, 1463–1472. [Google Scholar] [CrossRef]
- Kollman, P.A.; Massova, I.; Reyes, C.; Kuhn, B.; Huo, S.; Chong, L.; Lee, M.; Lee, T.; Duan, Y.; Wang, W.; et al. Calculating Structures and Free Energies of Complex Molecules: Combining Molecular Mechanics and Continuum Models. Acc. Chem. Res. 2000, 33, 889–897. [Google Scholar] [CrossRef] [PubMed]
- Valdés-Tresanco, M.S.; Valdés-Tresanco, M.E.; Valiente, P.A.; Moreno, E. Gmx_MMPBSA: A New Tool to Perform End-State Free Energy Calculations with GROMACS. J. Chem. Theory Comput. 2021, 17, 6281–6291. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A Free Web Tool to Evaluate Pharmacokinetics, Drug-Likeness and Medicinal Chemistry Friendliness of Small Molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated Data and New Features for Efficient Prediction of Protein Targets of Small Molecules. Nucleic Acids Res. 2019, 47, W357–W3664. [Google Scholar] [CrossRef]
- Varsou, D.D.; Melagraki, G.; Sarimveis, H.; Afantitis, A. MouseTox: An Online Toxicity Assessment Tool for Small Molecules through Enalos Cloud Platform. Food Chem. Toxicol. 2017, 110, 83–93. [Google Scholar] [CrossRef]
- Azucena, R.C.I.; Roberto, C.L.J.; Martin, Z.R.; Rafael, C.Z.; Leonardo, H.H.; Gabriela, T.P.; Araceli, C.R. Drug Susceptibility Testing and Synergistic Antibacterial Activity of Curcumin with Antibiotics against Enterotoxigenic Escherichia Coli. Antibiotics 2019, 8, 43. [Google Scholar] [CrossRef]
- Huband, M.D.; Bradford, P.A.; Otterson, L.G.; Basarab, G.S.; Kutschke, A.C.; Giacobbe, R.A.; Patey, S.A.; Alm, R.A.; Johnstone, M.R.; Potter, M.E.; et al. In Vitro Antibacterial Activity of AZD0914, a New Spiropyrimidinetrione DNA Gyrase/Topoisomerase Inhibitor with Potent Activity against Gram-Positive, Fastidious Gram-Negative, and Atypical Bacteria. Antimicrob. Agents Chemother. 2015, 59, 467–474. [Google Scholar] [CrossRef]
- Garrigós, C.; Murillo, O.; Lora-Tamayo, J.; Verdaguer, R.; Tubau, F.; Cabellos, C.; Cabo, J.; Ariza, J. Fosfomycin-Daptomycin and Other Fosfomycin Combinations as Alternative Therapies in Experimental Foreign-Body Infection by Methicillin-Resistant Staphylococcus Aureus. Antimicrob. Agents Chemother. 2013, 57, 606–610. [Google Scholar] [CrossRef] [PubMed]
- Belley, A.; Neesham-Grenon, E.; Arhin, F.F.; McKay, G.A.; Parr, T.R.; Moeck, G. Assessment by Time-Kill Methodology of the Synergistic Effects of Oritavancin in Combination with Other Antimicrobial Agents against Staphylococcus Aureus. Antimicrob. Agents Chemother. 2008, 52, 3820–3822. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Results of Growth of S. aureus, E. faecalis, and E. coli Exposed to Selected Anthraquinones. | |||
|---|---|---|---|
| S. aureus | E. faecalis | E. coli | |
1,8-dihydroxy-4,5-dinitroanthraquinone | Absence | Absence | Presence |
1,8-dihydroxyanthraquinone | Presence | Presence | Presence |
1,8-dichloroanthraquinone | Presence | Presence | Presence |
| Effect of the Presence of Sub-MIC Concentrations of DHDNA in Bacteria Resistant to Selected Antibiotics | |
|---|---|
| S. aureus | |
| Ciprofloxacin (5 µg) | Resistant |
| Azithromycin (15 µg) | Resistant |
| Chloramphenicol (30 µg) | Resistant |
| Erythromycin (15 µg) | Resistant |
| Tetracycline (30 µg) | Resistant |
| Trimethoprim/sulfamethoxazole (25 µg) | Resistant |
| E. faecalis | |
| Ciprofloxacin (5 µg) | Resistant |
| Clindamycin (2 µg) | Resistant |
| Cefoxitin (30 µg) | Resistant |
| Cefuroxime (30 µg) | Resistant |
| Tetracycline (30 µg) | Resistant |
| Trimethoprim/sulfamethoxazole (25 µg) | Resistant |
| E. coli | |
| Ciprofloxacin (5 µg) | Resistant |
| Clarithromycin (15 µg) | Resistant |
| Ampicillin (10 µg) | Resistant |
| Amoxicillin (25 µg) | Resistant |
| Cephalexin (30 µg) | Resistant |
| Cefuroxime (30 µg) | Resistant |
| Chloramphenicol (30 µg) | Resistant |
| Trimethoprim/sulfamethoxazole (25 µg) | Resistant |
| MIC (µg/mL) | ||
|---|---|---|
| S. aureus | E. faecalis | |
| 1,8-dihydroxy-4,5-dinitroanthraquinone | 31.125 | 62.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amorim, J.; Vásquez, V.; Cabrera, A.; Martínez, M.; Carpio, J. In Silico and In Vitro Identification of 1,8-Dihydroxy-4,5-dinitroanthraquinone as a New Antibacterial Agent against Staphylococcus aureus and Enterococcus faecalis. Molecules 2024, 29, 203. https://doi.org/10.3390/molecules29010203
Amorim J, Vásquez V, Cabrera A, Martínez M, Carpio J. In Silico and In Vitro Identification of 1,8-Dihydroxy-4,5-dinitroanthraquinone as a New Antibacterial Agent against Staphylococcus aureus and Enterococcus faecalis. Molecules. 2024; 29(1):203. https://doi.org/10.3390/molecules29010203
Chicago/Turabian StyleAmorim, Juliana, Viviana Vásquez, Andrea Cabrera, Maritza Martínez, and Juan Carpio. 2024. "In Silico and In Vitro Identification of 1,8-Dihydroxy-4,5-dinitroanthraquinone as a New Antibacterial Agent against Staphylococcus aureus and Enterococcus faecalis" Molecules 29, no. 1: 203. https://doi.org/10.3390/molecules29010203