
Phytochemical Diversity Comparison in Leaves and Roots of Wild and Micropropagated Latvian Sea Holly (Eryngium maritimum L.)

Abstract
:1. Introduction
2. Results and Discussion
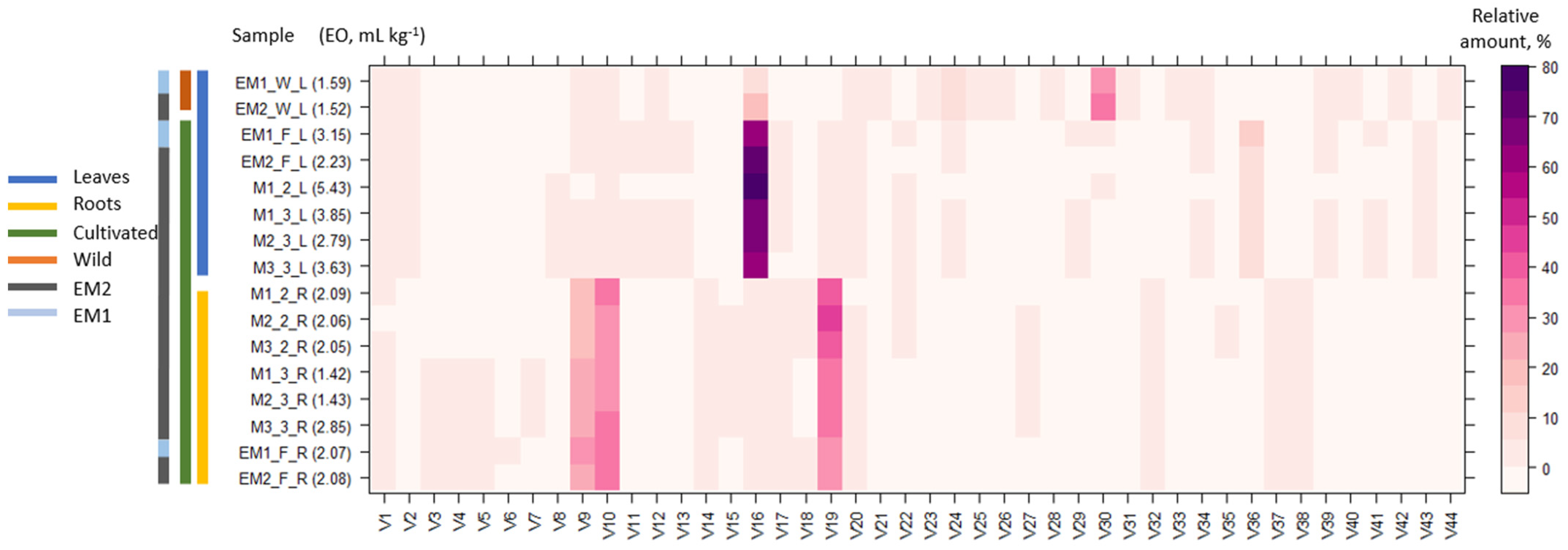
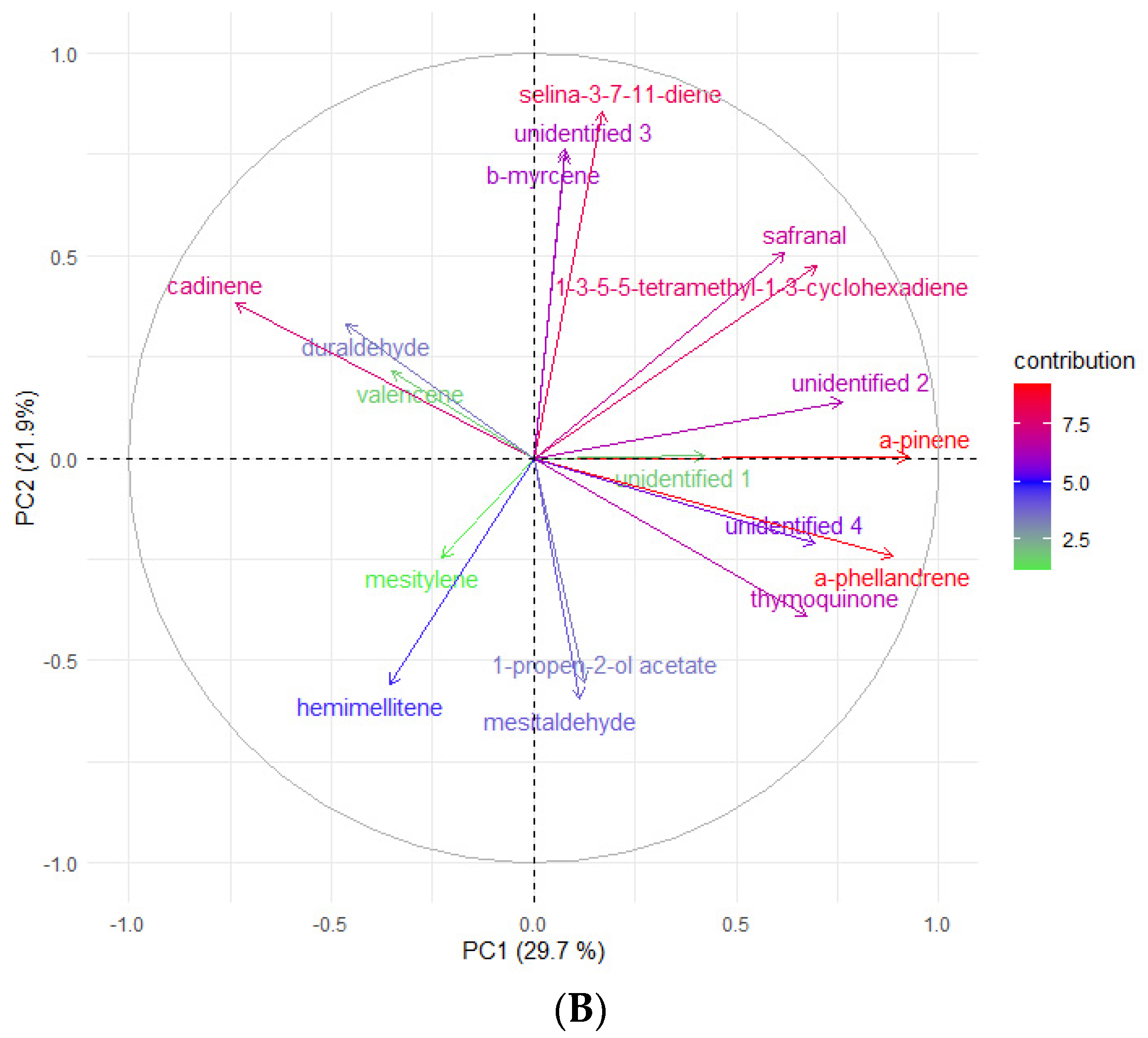
2.1. Essential Oil Quantification and Volatile Profile Characterization by GC-MS
2.2. Phytochemical Screening of E. maritimum Plant Extracts
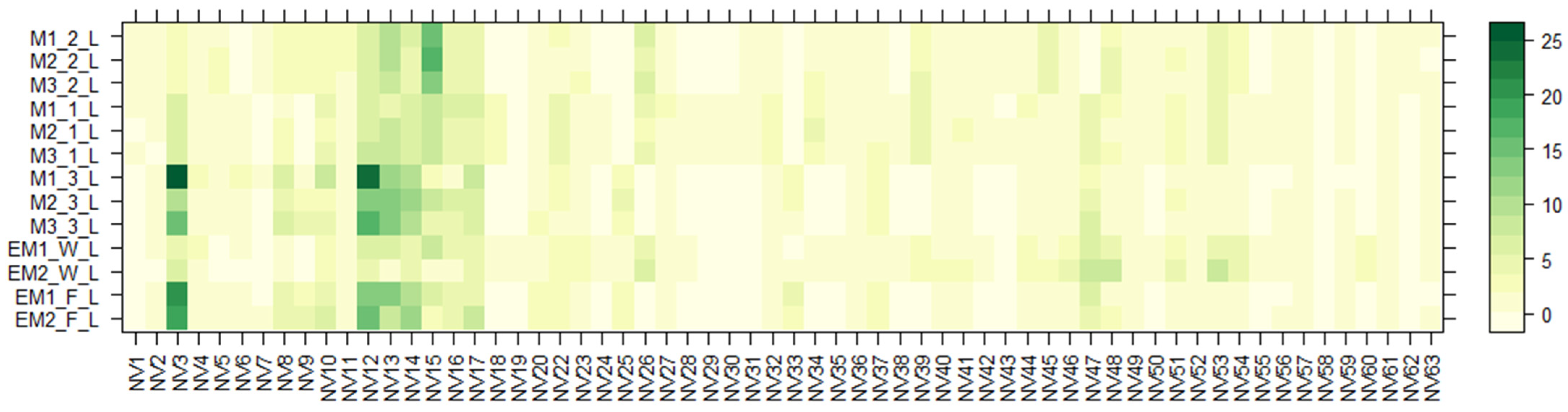
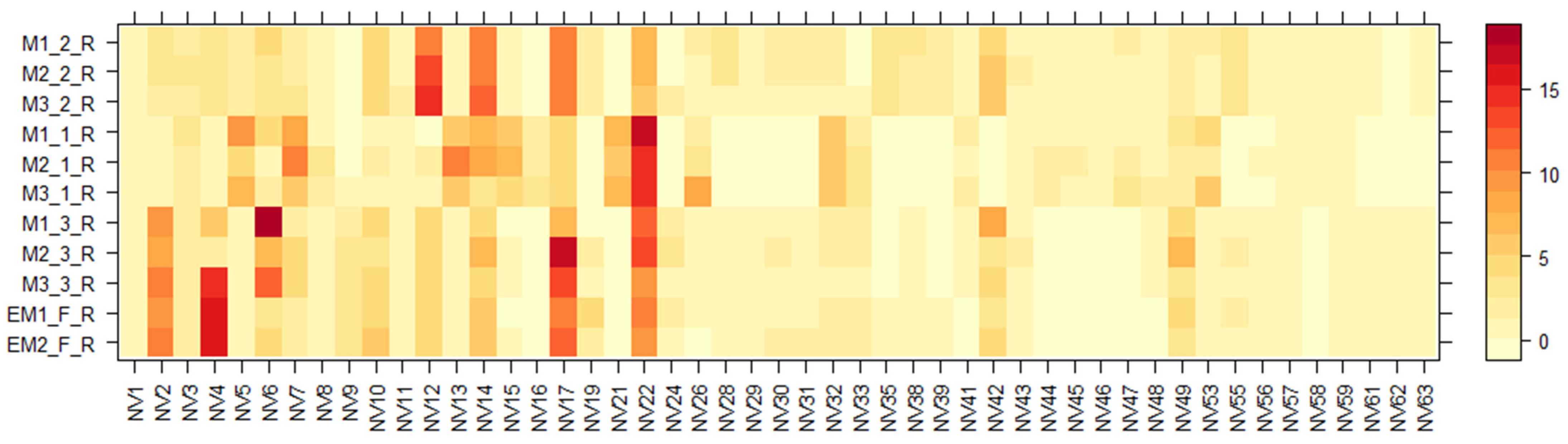
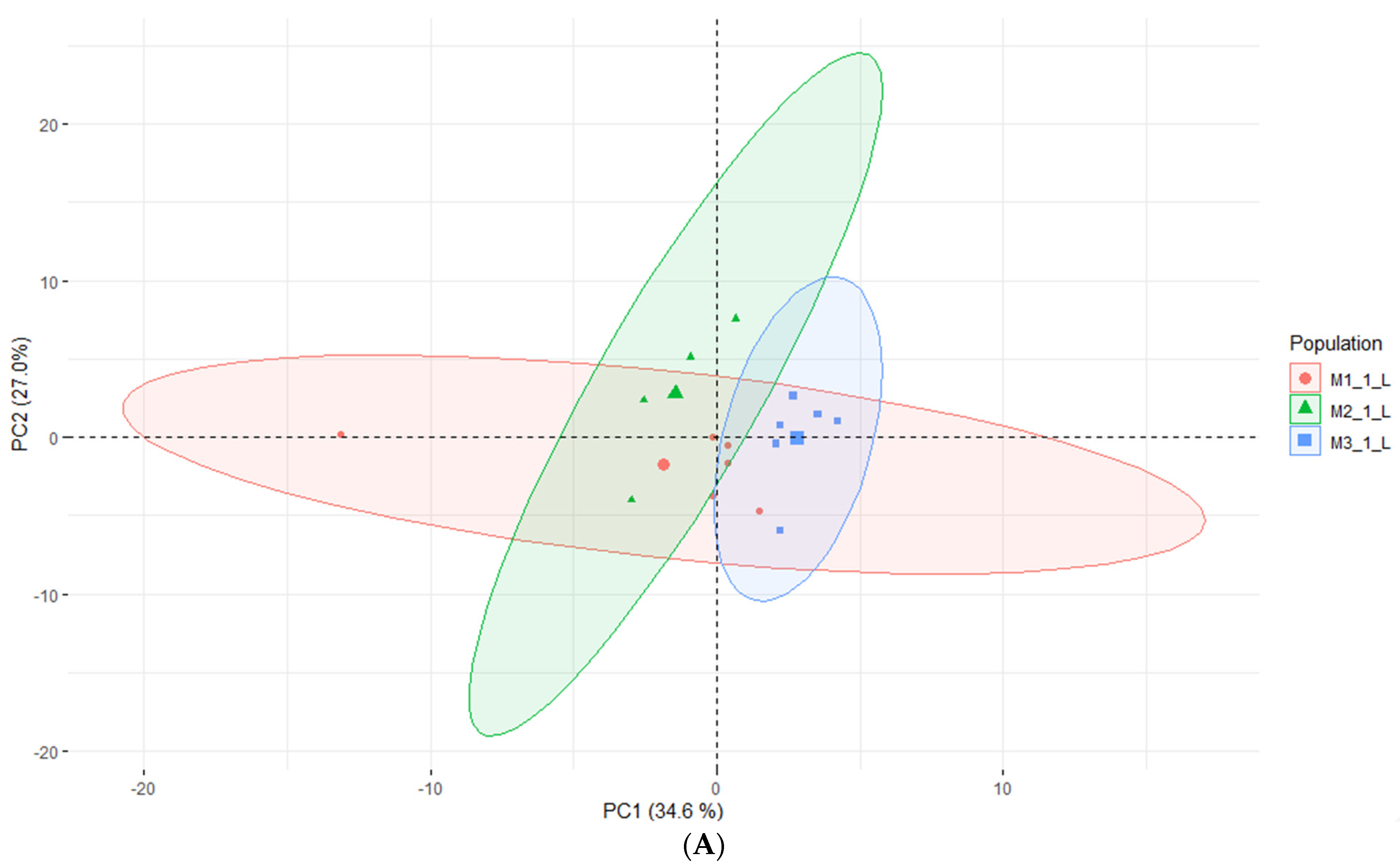
2.3. LC-MS of E. maritimum Plant Extracts
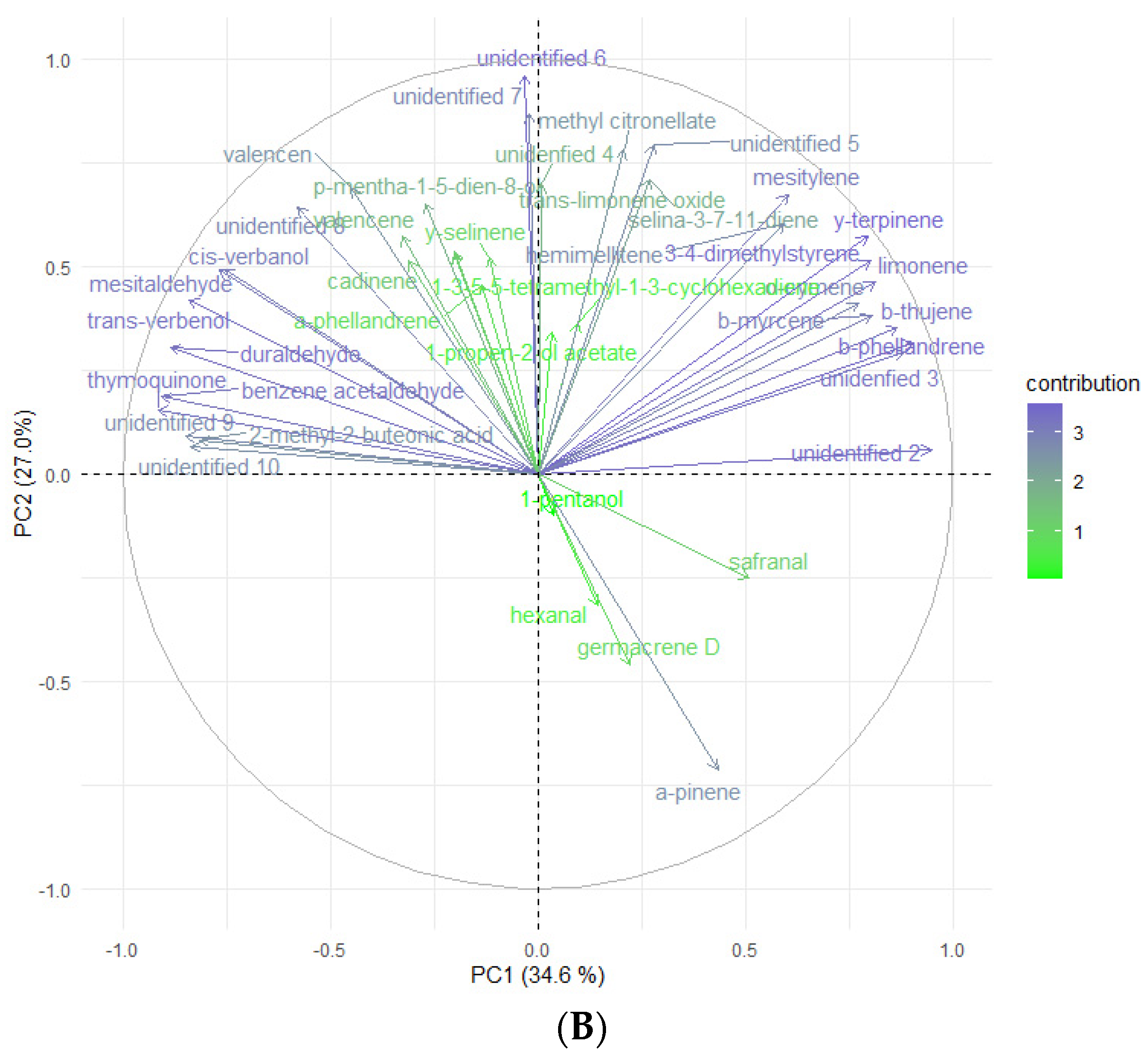
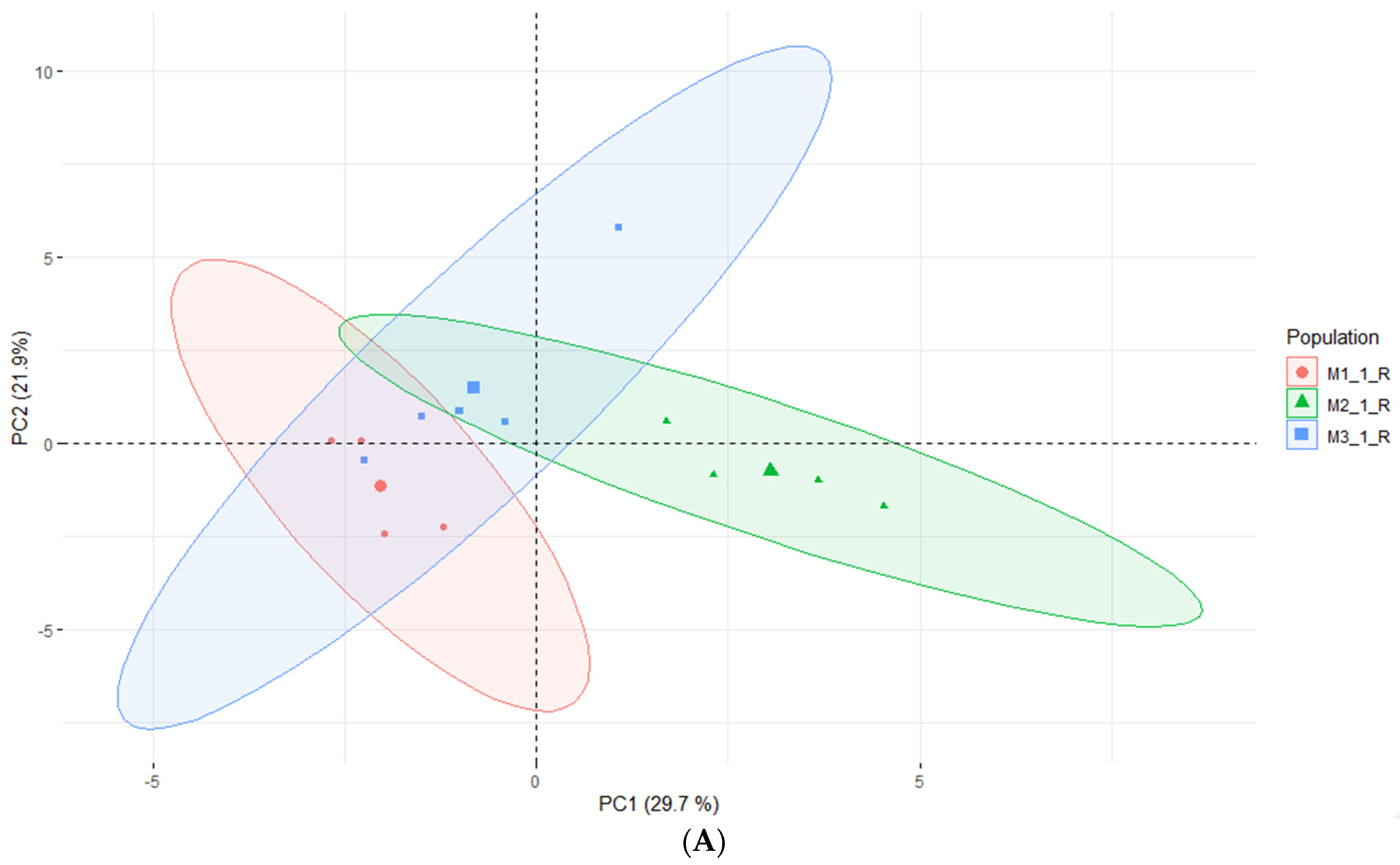
2.4. Headspace-GC-MS Analysis of E. maritimum In Vitro Micropropagated Plants
3. Materials and Methods
3.1. Plant Materials
3.2. Chemicals and Reagents
3.3. Essential Oil Quantification and Volatile Profile Characterization by GC-MS
3.4. The Identification and Quantification of Volatile Compounds by HS-GC-MS Analysis
3.5. Extract Preparation for UHPLC-HRMS Analysis and 96-Well Plate Assays
3.6. UHPLC-HRMS Analysis
3.7. 96-Well Plate Assays
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Clausing, G.; Vickers, K.; Kadereit, J.W. Historical Biogeography in a Linear System: Genetic Variation of Sea Rocket (Cakile maritima) and Sea Holly (Eryngium maritimum) along European Coasts. Mol. Ecol. 2000, 9, 1823–1833. [Google Scholar] [CrossRef] [PubMed]
- Necajeva, J.; Ievinsh, G. Seed Dormancy and Germination of an Endangered Coastal Plant Eryngium maritimum (Apiaceae). Est. J. Ecol. 2013, 62, 150–161. [Google Scholar] [CrossRef]
- Ingelög, T.; Andersson, R.; Tjernberg, M. Red Data Book of The BALTIC Region; Swedish Treatened Species Unit in cooperation with the Institute of Biology: Uppsala, Sweden, 1993. [Google Scholar]
- Isermann, M.; Rooney, P. Biological Flora of the British Isles: Eryngium maritimum. J. Ecol. 2014, 102, 789–821. [Google Scholar] [CrossRef]
- Kikowska, M.; Kalemba, D.; Dlugaszewska, J.; Thiem, B. Chemical Composition of Essential Oils from Rare and Endangered Species—Eryngium maritimum L. and E. alpinum L. Plants 2020, 9, 417. [Google Scholar] [CrossRef] [PubMed]
- Kikowska, M.; Thiem, B.; Sliwinska, E.; Rewers, M.; Kowalczyk, M.; Stochmal, A.; Oleszek, W. The Effect of Nutritional Factors and Plant Growth Regulators on Micropropagation and Production of Phenolic Acids and Saponins from Plantlets and Adventitious Root Cultures of Eryngium Maritimum L. J. Plant Growth Regul. 2014, 33, 809–819. [Google Scholar] [CrossRef]
- Kikowska, M.; Chanaj-Kaczmarek, J.; Derda, M.; Budzianowska, A.; Thiem, B.; Ekiert, H.; Szopa, A. The Evaluation of Phenolic Acids and Flavonoids Content and Antiprotozoal Activity of Eryngium Species Biomass Produced by Biotechnological Methods. Molecules 2022, 27, 363. [Google Scholar] [CrossRef] [PubMed]
- Sultana, K.W.; Das, S.; Chandra, I.; Roy, A. Efficient Micropropagation of Thunbergia coccinea Wall. and Genetic Homogeneity Assessment through RAPD and ISSR Markers. Sci. Rep. 2022, 12, 1683. [Google Scholar] [CrossRef]
- Ayuso, M.; García-Pérez, P.; Ramil-Rego, P.; Gallego, P.P.; Barreal, M.E. In Vitro Culture of the Endangered Plant Eryngium viviparum as Dual Strategy for Its Ex Situ Conservation and Source of Bioactive Compounds. Plant Cell Tissue Organ Cult. 2019, 138, 427–435. [Google Scholar] [CrossRef]
- Mežaka, I.; Kļaviņa, D.; Kaļāne, L.; Kronberga, A. Large-Scale In Vitro Propagation and Ex Vitro Adaptation of the Endangered Medicinal Plant Eryngium maritimum L. Horticulturae 2023, 9, 271. [Google Scholar] [CrossRef]
- Lajnef, H.B.; Ferioli, F.; Pasini, F.; Politowicz, J.; Khaldi, A.; D’Antuono, L.F.; Caboni, M.F.; Nasri, N. Chemical Composition and Antioxidant Activity of the Volatile Fraction Extracted from Air-Dried Fruits of Tunisian Eryngium maritimum L. Ecotypes. J. Sci. Food Agric. 2018, 98, 635–643. [Google Scholar] [CrossRef]
- Erdem, S.A.; Nabavi, S.F.; Orhan, I.E.; Daglia, M.; Izadi, M.; Nabavi, S.M. Blessings in disguise: A review of phytochemical composition and antimicrobial activity of plants belonging to the genus Eryngium. DARU J. Pharm. Sci. 2015, 23, 53. [Google Scholar] [CrossRef] [PubMed]
- Kholkhal, W.; Ilias, F.; Bekhechi, C.; Bekkara, F. Eryngium maritimum: A rich medicinal plant of polyphenols and flavonoids compounds with antioxidant, antibacterial and antifungal activities. Curr. Res. J. Biol. Sci. 2012, 4, 437–443. [Google Scholar]
- Mejri, H.; Tir, M.; Feriani, A.; Ghazouani, L.; Allagui, M.; Saidani-Tounsi, M. Does Eryngium maritimum seeds extract protect against CCl4 and cisplatin induced toxicity in rats: Preliminary phytochemical screening and assessment of its in vitro and in vivo antioxidant activity and antifibrotic effect. J. Funct. Foods 2017, 37, 363–372. [Google Scholar] [CrossRef]
- Pereira, C.G.; Locatelli, M.; Innosa, D.; Cacciagrano, F.; Polesná, L.; Santos, T.F.; Rodrigues, M.J.; Custódio, L. Unravelling the potential of the medicinal halophyte Eryngium maritimum L.: In vitro inhibition of diabetes-related enzymes, antioxidant potential, polyphenolic profile and mineral composition. S. Afr. J. Bot. 2019, 120, 204–212. [Google Scholar] [CrossRef]
- Benmerache, A.; Magid, A.A.; Berrehal, D.; Kabouche, A.; Voutquenne-Nazabadioko, L.; Messaili, S.; Abedini, A.; Harakat, D.; Kabouche, Z. Chemical composition, antibacterial, antioxidant and tyrosinase inhibitory activities of glycosides from aerial parts of Eryngium tricuspidatum L. Phytochem. Lett. 2016, 18, 23–28. [Google Scholar] [CrossRef]
- Traversier, M.; Gaslonde, T.; Lecso, M.; Michel, S.; Delannay, E. Comparison of extraction methods for chemical composition, antibacterial, depigmenting and antioxidant activities of Eryngium maritimum. Int. J. Cosmet. Sci. 2020, 42, 127–135. [Google Scholar] [CrossRef]
- Seppic: Unique Ingredients, Created for Products Used Every Day. Available online: https://www.seppic.com/en (accessed on 29 April 2023).
- Fabbrimarine la Cosmetica Marina. Available online: https://www.fabbricosmetica.com/en (accessed on 29 April 2023).
- Petropoulos, S.A.; Karkanis, A.; Martins, N.; Ferreira, I.C.F.R. Edible halophytes of the Mediterranean basin: Potential candidates for novel food products. Trends Food Sci. Technol. 2018, 74, 69–84. [Google Scholar] [CrossRef]
- Rodrigues, T.L.M.; Silva, M.E.P.; Gurgel, E.S.C.; Oliveira, M.S.; Lucas, F.C.A. Eryngium foetidum L. (Apiaceae): A Literature Review of Traditional Uses, Chemical Composition, and Pharmacological Activities. Evid. Based Complement. Alternat. Med. 2022, 14, 2896895. [Google Scholar] [CrossRef]
- The Local Food-Nutraceuticals Consortium. Understanding local Mediterranean diets: A multidisciplinary pharmacological and ethnobotanical approach. Pharmacol. Res. 2005, 52, 353–366. [Google Scholar] [CrossRef]
- Wang, P.; Su, Z.; Yuan, W.; Deng, G.; Li, S. Phytochemical Constituents and Pharmacological Activities of Eryngium L. (Apiaceae). Pharm. Crop. 2012, 3, 99–120. [Google Scholar] [CrossRef]
- Maggio, A.; Bruno, M.; Formisano, C.; Rigano, D.; Senatore, F. Chemical Composition of the Essential Oils of Three Species of Apiaceae Growing Wild in Sicily: Bonannia graeca, Eryngium maritimum and Opopanax chironium. Nat. Prod. Commun. 2013, 8, 841–844. [Google Scholar] [CrossRef]
- Darriet, F.; Andreani, S.; De Cian, M.-C.; Costa, J.; Muselli, A. Chemical variability and antioxidant activity of Eryngium maritimum L. essential oils from Corsica and Sardinia. Flavour Fragr. J. 2014, 29, 3–13. [Google Scholar] [CrossRef]
- Beeby, E.; Magalhães, M.; Poças, J.; Collins, T.; Lemos, M.F.L.; Barros, L.; Ferreira, I.C.F.R.; Cabral, C.; Pires, I.M. Secondary metabolites (essential oils) from sand-dune plants induce cytotoxic effects in cancer cells. J. Ethnopharmacol. 2020, 258, 112803. [Google Scholar] [CrossRef]
- Matejić, J.S.; Stojanović-Radić, Z.Z.; Krivošej, Z.D.; Zlatković, B.K.; Marin, P.D.; Džamić, A.M. Biological activity of extracts and essential oils of two Eryngium (Apiaceae) species from the Balkan peninsula. Acta Med. Median. 2019, 58, 24–31. [Google Scholar] [CrossRef]
- Darriet, F.; Bendahou, M.; Desjobert, J.M.; Costa, J.; Muselli, A. Bicyclo[4.4.0]decane oxygenated sesquiterpenes from Eryngium maritimum essential oil. Planta Med. 2012, 78, 386–389. [Google Scholar] [CrossRef] [PubMed]
- Meot-Duros, L.; Le Floch, G.; Magné, C. Radical scavenging, antioxidant and antimicrobial activities of halophytic species. J. Ethnopharmacol. 2008, 116, 258–262. [Google Scholar] [CrossRef] [PubMed]
- Rjeibi, I.; Saad, A.B.; Ncib, S.; Souid, S. Phenolic composition and antioxidant properties of Eryngium maritimum (sea holly). J. Coast. Life Med. 2017, 5, 212–215. [Google Scholar] [CrossRef]
- Kikowska, M.; Kruszka, D.; Derda, M.; Hadaś, E.; Thiem, B. Phytochemical Screening and Acanthamoebic Activity of Shoots from in Vitro Cultures and in Vivo Plants of Eryngium alpinum L.—The Endangered and Protected Species. Molecules 2020, 25, 1416. [Google Scholar] [CrossRef]
- Ali, A.; Bashmil, Y.M.; Cottrell, J.J.; Suleria, H.A.R.; Dunshea, F.R. LC-MS/MS-QTOF Screening and Identification of Phenolic Compounds from Australian Grown Herbs and Their Antioxidant Potential. Antioxidants 2021, 10, 1770. [Google Scholar] [CrossRef]
- Mugford, S.T.; Osbourn, A. Saponin Synthesis and Function. In Isoprenoid Synthesis in Plants and Microorganisms; Bach, T., Rohmer, M., Eds.; Springer: New York, NY, USA, 2012; Volume 28, pp. 405–424. [Google Scholar] [CrossRef]
- Kikowska, M.; Długaszewska, J.; Kubicka, M.M.; Kędziora, I.; Budzianowski, J.; Thiem, B. In vitro antimicrobial activity of extracts and their fractions from three Eryngium L. species. Herba Pol. 2016, 62, 67–77. [Google Scholar] [CrossRef]
- Kowalczyk, M.; Masullo, M.; Thiem, B.; Piacente, S.; Stochmal, A.; Oleszek, W. Three new triterpene saponins from roots of Eryngium planum. Nat. Prod. Res. 2014, 28, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Tembeni, B.; Oyedeji, O.O.; Manene, C.N.; Oyemitan, A.I.; Oyedeji, A.O. Anti-inflammatory, Analgesic Activity and Toxicity of Two Pelargonium inquinans Ait Essential Oils: Wild and Cultivated. J. Essent. Oil Bear. Plants 2019, 22, 1252–1264. [Google Scholar] [CrossRef]
- El Gendy, A.N.; Leonardi, M.; Mugnaini, L.; Bertelloni, F.; Ebani, V.V.; Nardoni, S.; Mancianti, F.; Hendawy, S.; Omer, E.; Pistelli, L. Chemical composition and antimicrobial activity of essential oil of wild and cultivated Origanum syriacum plants grown in Sinai, Egypt. Ind. Crop. Prod. 2015, 67, 201–207. [Google Scholar] [CrossRef]
- Sile, I.; Krizhanovska, V.; Nakurte, I.; Mezaka, I.; Kalane, L.; Filipovs, J.; Vecvanags, A.; Pugovics, O.; Grinberga, S.; Dambrova, M.; et al. Wild-Grown and Cultivated Glechoma hederacea L.: Chemical Composition and Potential for Cultivation in Organic Farming Conditions. Plants 2022, 11, 819. [Google Scholar] [CrossRef] [PubMed]
- Stanković, M.; Zlatić, N.; Mašković, J.; Mašković, P.; Jakovljević, D. Teucrium scordium L. and Mentha pulegium L. essential oil importance in adaptive response to salinity stress. Biochem. Syst. Ecol. 2022, 102, 104419. [Google Scholar] [CrossRef]
- Machado, M.; Santoto, G.; Sousa, M.C.; Salgueiro, L.; Cavaleiro, C. Activity of essential oils on the growth of Leishmania infantum promastigotes. Flavour Fragr. J. 2010, 25, 156–160. [Google Scholar] [CrossRef]
- Amessis-Ouchemoukh, N.; Madani, K.; Falé, P.L.V.; Serralheiro, M.L.; Araújo, M.E.M. Antioxidant capacity and phenolic contents of some Mediterranean medicinal plants and their potential role in the inhibition of cyclooxygenase-1 and acetylcholinesterase activities. Ind. Crop. Prod. 2014, 53, 6–15. [Google Scholar] [CrossRef]
- Cortés-Fernández, I.; Sureda, A.; Adrover, M.; Caprioli, G.; Maggi, F.; Gil-Vives, L.; Capó, X. Antioxidant and anti-inflammatory potential of rhizome aqueous extract of sea holly (Eryngium maritimum L.) on Jurkat cells. J. Ethnopharmacol. 2023, 305, 116120. [Google Scholar] [CrossRef]
- Küpeli, E.; Kartal, M.; Aslan, S.; Yesilada, E. Comparative evaluation of the anti-inflammatory and antinociceptive activity of Turkish Eryngium species. J. Ethnopharmacol. 2006, 107, 32–37. [Google Scholar] [CrossRef]
- Kwakman, P.H.S.; te Velde, A.A.; de Boer, L.; Vandenbroucke-Grauls, C.M.J.; Zaat, S.A.J. Two major medicinal honeys have different mechanisms of bactericidal activity. PLoS ONE 2011, 6, e17709. [Google Scholar] [CrossRef]
- Mizzi, L.; Maniscalco, D.; Gaspari, S.; Chatzitzika, C.; Gatt, R.; Valdramidis, V.P. Assessing the individual microbial inhibitory capacity of different sugars against pathogens commonly found in food systems. Lett. Appl. Microbiol. 2020, 71, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Nakurte, I.; Berga, M.; Pastare, L.; Kienkas, L.; Senkovs, M.; Boroduskis, M.; Ramata-Stunda, A. Valorization of Bioactive Compounds from By-Products of Matricaria recutita White Ray Florets. Plants 2023, 12, 396. [Google Scholar] [CrossRef] [PubMed]
- Trovato, M.; Funck, D.; Forlani, G.; Okumoto, S.; Amir, R. Amino Acids in Plants: Regulation and Functions in Development and Stress Defense. Front. Plant Sci. 2021, 12, 772810. [Google Scholar] [CrossRef]
- Taç, S.; Özcan, T. Fatty Acid and Amino Acid Profiles in Some Dune Vegetation Species from Istanbul. Süleyman Demirel Univ. J. Nat. Appl. Sci. 2019, 23, 892–903. [Google Scholar] [CrossRef]
- Trócsányi, E.; György, Z.; Zámboriné-Németh, É. New insights into rosmarinic acid biosynthesis based on molecular studies. Curr. Plant Biol. 2020, 23, 100162. [Google Scholar] [CrossRef]
- Guan, H.; Luo, W.; Bao, B.; Cao, Y.; Cheng, F.; Yu, S.; Fan, Q.; Zhang, L.; Wu, Q.; Shan, M. A Comprehensive Review of Rosmarinic Acid: From Phytochemistry to Pharmacology and Its New Insight. Molecules 2022, 27, 3292. [Google Scholar] [CrossRef] [PubMed]
- Magaña, A.A.; Kamimura, N.; Soumyanath, A.; Stevens, J.F.; Maier, C.S. Caffeoylquinic acids: Chemistry, biosynthesis, occurrence, analytical challenges, and bioactivity. Plant J. 2021, 107, 1299–1319. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, S.; Ownby, S.; Wang, P.; Yuan, W.; Zhang, W.; Scott Beasley, R. Phenolic compounds and rare polyhydroxylated triterpenoid saponins from Eryngium yuccifolium. Phytochemistry 2008, 69, 2070–2080. [Google Scholar] [CrossRef]
- Belliardo, F.F.; Bicchi, C.C.; Cordero, C.C.; Liberto, E.; Rubiolo, P.; Sgorbini, B. Headspace-Solid-Phase Microextraction in the Analysis of the Volatile Fraction of Aromatic and Medicinal Plants. J. Chromatogr. Sci. 2006, 44, 416–429. [Google Scholar] [CrossRef]
- Vador, N.; Vador, B.; Hole, R. Simple spectrophotometric methods for standardizing ayurvedic formulation. Indian J. Pharm. Sci. 2012, 74, 161–163. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | RI a | Compound b | Composition Range (Leaves) | Composition Range (Roots) |
|---|---|---|---|---|
| V1 | 921 | Cumene | 0.18–5.15 | n.d–2.39 |
| V2 | 937 | α-Pinene | 0.60–4.23 | n.d |
| V3 | 972 | Mesitylene | n.d | n.d–0.83 |
| V4 | 990 | Pseudocumene | n.d | n.d–0.41 |
| V5 | 1201 | Safranal | n.d | n.d–0.43 |
| V6 | 1283 | α-Terpinen-7-al | n.d | n.d–0.19 |
| V7 | 956 | Dehydrosabinene | n.d | n.d–0.28 |
| V8 | 1293 | Dihydroedulan | 0.61 | n.d |
| V9 | 1337 | Mesitaldehyde | n.d–1.57 | 16.15–27.30 |
| V10 | 1364 | Duraldehyde | 1.03–2.59 | 27.62–34.16 |
| V11 | 1376 | α-Copaene | n.d–1.09 | n.d |
| V12 | 1384 | α-Bourbonene | n.d–1.18 | n.d |
| V13 | 1448 | Isogermacrene D | n.d–0.72 | n.d |
| V14 | 1375 | Isoledene | n.d | 0.56–1.31 |
| V15 | 1464 | dehydro-Aromadendrene | n.d | n.d–0.42 |
| V16 | 1481 | Germacrene D | 17.92–75.05 | 0.39–2.55 |
| V17 | 1409 | α-Gurjunene | n.d–1.14 | 0.12–0.88 |
| V18 | 1492 | Valencene | n.d | n.d–1.05 |
| V19 | 1499 | α-Muurolene | n.d–4.85 | 29.64–46.83 |
| V20 | 1481 | Cadina-1(6),4-diene | 0.72–1.21 | n.d–1.10 |
| V21 | 1506 | α-Farnesene | n.d–1.67 | n.d |
| V22 | 1645 | δ-Cadinol | n.d–1.55 | n.d–1.04 |
| V23 | 1573 | 1,5-epoxysalvial-4(14)-ene | n.d–2.29 | n.d |
| V24 | 1576 | Spathulenol | n.d–8.02 | n.d |
| V25 | 1581 | Caryophyllene oxide | n.d–3.45 | n.d |
| V26 | 1595 | Mint ketone | n.d–3.09 | n.d |
| V27 | 1490 | β-Guaiene | n.d | n.d–0.45 |
| V28 | 1440 | Aromadendrane | n.d–0.91 | n.d |
| V29 | 1631 | Ledene oxide-(II) | n.d–2.28 | n.d |
| V30 | 1695 | 4βH-Muurol-9-en-15-al | n.d–33.79 | n.d |
| V31 | 1619 | Patchoulane | n.d–1.02 | n.d |
| V32 | 1635 | β-Vatirenene | n.d | 0.33–1.12 |
| V33 | 1778 | Isovalencenol | n.d–0.88 | n.d |
| V34 | 1648 | Alloaromadendrene oxide-1 | n.d–2.48 | n.d |
| V35 | 1640 | tau-Cadinol | n.d | n.d–0.36 |
| V36 | 1671 | Eudesma-4,7-diene-1β-ol | n.d–11.95 | n.d |
| V37 | 1712 | (E)-γ-Atlantone | n.d | 0.3–1.11 |
| V38 | 2040 | Falcarinol | n.d | 0.11–0.66 |
| V39 | 1742 | 4βH-Cadin-9-en-15-ol | n.d–5.14 | n.d |
| V40 | 1876 | Murolan-3,9(11)-diene-10-peroxy | n.d–1.32 | n.d |
| V41 | 1762 | Methyl farnesate | n.d–0.88 | n.d |
| V42 | 1777 | 15-hydroxy-α-muurolene | n.d–1.49 | n.d |
| V43 | 1837 | Neophytadiene | n.d–2.50 | n.d |
| V44 | 1844 | 6,10,14-trimethyl-2-Pentadecanone | n.d–0.59 | n.d |
| Yields, mL kg−1 | 1.52–5.43 | 1.42–2.85 | ||
| Sample Code | TPC a, g GAE/100 g | TSC b, g ESC/100 g | Sugars c, g GLE/100 g | ARA d, mg ASE/100 g | DPPH e Quenched, % | IC50, µg/mL |
|---|---|---|---|---|---|---|
| EM1 W_L | 0.64 ± 0.10 | 8.39 ± 0.21 | 5.91 ± 0.23 | 87.20 ± 1.26 | 17.5 ± 0.69 | 43.76 ± 1.51 |
| EM2 W_L | 0.46 ± 0.03 | 6.51 ± 0.16 | 2.49 ± 0.11 | 78.90 ± 1.14 | 15.8 ± 0.63 | 39.60 ± 1.38 |
| EM1 F_L | 0.83 ± 0.07 | 3.95 ± 0.19 | 2.63 ± 0.11 | 57.58 ± 0.83 | 11.6 ± 0.46 | 28.89 ± 0.99 |
| EM2 F_L | 0.77 ± 0.05 | 6.04 ± 0.25 | 2.22 ± 0.09 | 54.41 ± 0.78 | 10.9 ± 0.43 | 27.31 ± 0.94 |
| EM1 F_R | 0.22 ± 0.02 | 6.77 ± 0.30 | 12.76 ± 0.48 | 19.47 ± 0.52 | 3.9 ± 0.21 | 9.77 ± 0.36 |
| EM2 F_R | 0.19 ± 0.01 | 6.25 ± 0.20 | 12.68 ± 0.48 | 24.81 ± 0.60 | 5.0 ± 0.27 | 12.45 ± 0.46 |
| M1_1_L | 0.91 ± 0.12 | 6.64 ± 0.22 | 0.91 ± 0.05 | 115.20 ± 1.63 | 23.1 ± 0.88 | 57.81 ± 1.96 |
| M2_1_L | 0.85 ± 0.08 | 6.76 ± 0.32 | 0.85 ± 0.05 | 102.00 ± 1.47 | 20.5 ± 0.78 | 51.19 ± 1.74 |
| M3_1_L | 0.91 ± 0.09 | 8.27 ± 0.36 | 1.58 ± 0.07 | 117.30 ± 1.67 | 23.5 ± 0.90 | 58.87 ± 2.00 |
| M1_1_R | 0.97 ± 0.08 | 8.22 ± 0.48 | 16.56 ± 0.50 | 211.60 ± 2.99 | 42.5 ± 1.61 | 106.19 ± 3.51 |
| M2_1_R | 0.94 ± 0.07 | 10.27 ± 0.44 | 16.62 ± 0.47 | 202.90 ± 2.88 | 40.7 ± 1.54 | 101.83 ± 3.34 |
| M3_1_R | 0.80 ± 0.06 | 9.62 ± 0.34 | 15.48 ± 0.51 | 201.60 ± 2.73 | 40.5 ± 1.53 | 101.17 ± 3.32 |
| M1_2_L | 0.90 ± 0.07 | 8.64 ± 0.46 | 6.38 ± 0.24 | 179.18 ± 2.54 | 36.0 ± 1.37 | 89.92 ± 2.93 |
| M2_2_L | 1.02 ± 0.11 | 10.07 ± 0.42 | 6.93 ± 0.26 | 183.74 ± 2.61 | 36.9 ± 1.41 | 92.21 ± 3.01 |
| M3_2_L | 0.89 ± 0.05 | 9.39 ± 0.10 | 5.84 ± 0.22 | 170.33 ± 2.41 | 34.2 ± 1.29 | 85.48 ± 2.74 |
| M1_2_R | 0.08 ± 0.01 | 2.97 ± 0.12 | 21.85 ± 0.65 | 34.24 ± 0.49 | 6.9 ± 0.29 | 17.18 ± 0.64 |
| M2_2_R | 0.10 ± 0.02 | 3.26 ± 0.15 | 30.08 ± 0.78 | 38.12 ± 0.55 | 7.7 ± 0.33 | 19.13 ± 0.71 |
| M3_2_R | 0.11 ± 0.02 | 3.14 ± 0.22 | 30.36 ± 0.87 | 41.79 ± 0.60 | 8.4 ± 0.36 | 20.97 ± 0.78 |
| M1_3_L | 0.58 ± 0.07 | 6.94 ± 0.24 | 4.06 ± 0.34 | 64.79 ± 0.93 | 13.0 ± 0.53 | 32.52 ± 1.15 |
| M2_3_L | 0.59 ± 0.05 | 7.18 ± 0.27 | 6.04 ± 0.36 | 66.95 ± 0.96 | 13.4 ± 0.55 | 33.60 ± 1.21 |
| M3_3_L | 0.60 ± 0.03 | 6.98 ± 0.18 | 5.45 ± 0.27 | 69.81 ± 1.00 | 14.0 ± 0.59 | 35.04 ± 1.26 |
| M1_3_R | 0.31 ± 0.02 | 5.53 ± 0.20 | 27.67 ± 0.67 | 41.62 ± 0.60 | 8.4 ± 0.36 | 20.89 ± 0.78 |
| M2_3_R | 0.37 ± 0.04 | 6.12 ± 0.33 | 36.85 ± 0.54 | 50.15 ± 0.72 | 10.1 ± 0.43 | 25.17 ± 0.93 |
| M3_3_R | 0.27 ± 0.06 | 4.54 ± 0.25 | 26.02 ± 0.40 | 48.14 ± 0.70 | 9.7 ± 0.41 | 24.16 ± 0.89 |
| No. | RT (min) | Proposed Compound | Tentative Molecular Formula | Ion Mode | Theoretical (m/z) | Observed (m/z) | Mass Error (ppm) | Class a |
|---|---|---|---|---|---|---|---|---|
| NV1 | 1.34 | Histidine | C6H9N3O2 | [M+H]+ | 156.0768 | 156.0764 | 0.4 | AA |
| NV2 | 1.36 | Arginine | C6H14N4O2 | [M+H]+ | 175.119 | 175.1156 | 3.4 | AA |
| NV3 | 1.58 | Sucrose | C12H22O11 | [M+K]+ | 381.0794 | 381.0803 | 0.9 | O |
| NV4 | 1.61 | Proline | C5H9NO2 | [M+H]+ | 116.0702 | 116.0695 | 0.7 | AA |
| NV5 | 1.62 | Methionine | C5H11NO2S | [M+H]+ | 150.0583 | 150.0572 | 1.1 | AA |
| NV6 | 1.64 | Glutamine | C5H10N2O3 | [M+H]+ | 147.0764 | 147.0772 | 0.8 | AA |
| NV7 | 1.65 | Asparagine | C4H8N2O3 | [M+H]+ | 133.0608 | 133.059 | 1.8 | AA |
| NV8 | 1.72 | N-(1-deoxy-D-fructos-1-yl)-L-Valine | C11H21NO7 | [M+H]+ | 280.1391 | 280.1382 | 0.9 | AA |
| NV9 | 1.80 | Valine | C5H11NO2 | [M+H]+ | 118.0863 | 118.0848 | 1.5 | AA |
| NV10 | 1.82 | Isoleucine | C6H13NO2 | [M+H]+ | 132.1019 | 132.1021 | 0.2 | AA |
| NV11 | 2.18 | Tyrosine | C9H11NO3 | [M+H]+ | 182.0812 | 182.0797 | 1.5 | AA |
| NV12 | 2.29 | Leucine | C6H13NO2 | [M+H]+ | 132.1019 | 132.1006 | 1.3 | AA |
| NV13 | 2.42 | N-(1-deoxy-D-fructos-1-yl)-L-Leucine | C12H23NO7 | [M+H]+ | 294.1562 | 294.1573 | 1.1 | AA |
| NV14 | 3.84 | Phenylalanine | C9H11NO2 | [M+H]+ | 166.0863 | 166.085 | 1.3 | AA |
| NV15 | 3.96 | N-(1-deoxy-D-fructos-1-yl)-L-Phenylalanine | C15H21NO7 | [M+H]+ | 328.1503 | 328.1516 | 1.3 | AA |
| NV16 | 5.07 | N-(1-deoxy-D-fructos-1-yl)-L-Tryptophan | C17H22N2O7 | [M+H]+ | 367.1538 | 367.1521 | 1.7 | AA |
| NV17 | 6.35 | Tryptophan | C11H12N2O2 | [M+H]+ | 205.0972 | 205.0983 | 1.1 | AA |
| NV18 | 6.96 | 7-methoxycoumarin | C10H8O3 | [M+H]+ | 177.0546 | 177.0552 | 0.6 | C |
| NV19 | 7.00 | Methyl cinnamate | C10H10O2 | [M+H]+ | 163.0754 | 163.0756 | 0.2 | CA |
| NV20 | 7.63 | Caffeic acid 3-O-glucuronide | C15H16O10 | [M+K]+ | 379.0636 | 379.064 | 0.4 | CA |
| NV21 | 7.71 | Neochlorogenic acid | C16H18O9 | [M+H]+ | 355.1024 | 355.1052 | 2.8 | HC |
| NV22 | 7.88 | Chlorogenic acid | C16H18O9 | [M+H]+ | 355.1024 | 355.1056 | 3.2 | HC |
| NV23 | 7.99 | p-Coumaric acid 4-O-glucoside | C15H18O8 | [M+NH4]+ | 344.1340 | 344.1379 | 3.9 | PG |
| NV24 | 8.41 | alpha-L-Fucopyranosyl-(1->2)-beta-D-galactopyranosyl-(1->4)-D-glucose isomer I | C18H32O15 | [M+H]+ | 489.1814 | 489.1799 | 1.5 | O |
| NV25 | 8.56 | Unknown compound | - | - | - | 409.185 | - | - |
| NV26 | 9.19 | Kaempferol 3-O-(6″-malonyl-glucoside) | C24H22O14 | [M+H]+ | 535.1082 | 535.114 | 5.8 | F |
| NV27 | 9.25 | n-p-Coumaroylquinic acid | C16H18O8 | [M+H]+ | 339.1074 | 339.1085 | 1.1 | QA |
| NV28 | 9.49 | alpha-L-Fucopyranosyl-(1->2)-beta-D-galactopyranosyl-(1->4)-D-glucose isomer II | C18H32O15 | [M+H]+ | 489.1814 | 489.1794 | 2.0 | O |
| NV29 | 9.97 | Phenolic glycoside | C16H22O8 | [M+Na]+ | 365.1207 | 365.1215 | 0.8 | PG |
| NV30 | 9.98 | Phenolic glycoside I | C16H24O7 | [M+H]+ | 329.1595 | 329.1614 | 1.9 | PG |
| NV31 | 9.99 | Phenolic glycoside II | C16H24O7 | [M+H]+ | 329.1595 | 329.1617 | 2.2 | PG |
| NV32 | 9.56 | n-Feruloylquinic acid I | C17H20O9 | [M+H]+ | 369.118 | 369.1186 | 0.6 | QA |
| NV33 | 10.01 | n-Feruloylquinic acid II | C17H20O9 | [M+H]+ | 369.118 | 369.1188 | 0.8 | QA |
| NV34 | 10.10 | 8-Epiiridodial glucoside | C16H26O7 | [M+NH4]+ | 348.2017 | 348.2047 | 3.0 | OG |
| NV35 | 10.75 | Phenolic glycoside | C19H26O10 | [M+NH4]+ | 432.1864 | 432.1892 | 2.8 | QA |
| NV36 | 11.43 | Quercetin 3-O-galactoside 7-O-rhamnoside | C27H30O16 | [M+H]+ | 611.1607 | 611.162 | 1.3 | F |
| NV37 | 11.56 | alpha-L-Rhamnopyranosyl-(1->3)-alpha-D-galactopyranosyl-(1->3)-L-fucose | C18H32O14 | [M+H]+ | 495.1684 | 495.1872 | 18.8 | O |
| NV38 | 11.94 | Unknown compound | - | - | - | 575.179 | - | - |
| NV39 | 11.95 | Phenylethyl primeveroside | C19H28O10 | [M+Na]+ | 439.1575 | 439.1602 | 2.7 | OG |
| NV40 | 11.97 | Quercetin 3-O-glucoside | C21H20O12 | [M+H]+ | 465.1028 | 465.1044 | 1.6 | F |
| NV41 | 12.18 | Quercetin 3-O-(6″-malonyl-glucoside) | C24H22O15 | [M+H]+ | 551.1031 | 551.1047 | 1.6 | F |
| NV42 | 12.57 | Phenolic glycoside | C19H26O10 | [M+H]+ | 415.1599 | 415.1671 | 7.2 | PG |
| NV43 | 12.64 | Ferulol | C10H14O2 | [M+H]+ | 167.1067 | 167.1063 | 0.4 | HC |
| NV44 | 12.76 | Luteolin 7-O-rutinoside/Kaempferol 3-O-galactoside | C27H30O15 | [M+H]+ | 595.1657 | 595.1679 | 2.2 | F |
| NV45 | 12.77 | Luteolin 6-C-glucoside/Kaempferol 3-O-galactoside | C21H20O11 | [M+H]+ | 449.1078 | 449.1097 | 1.9 | F |
| NV46 | 13.18 | Isorhamnetin 3-O-glucoside 7-O-rhamnoside | C28H32O16 | [M+H]+ | 625.1763 | 625.1797 | 3.4 | F |
| NV47 | 13.37 | Luteolin 6-C-glucoside/Kaempferol 3-O-galactoside | C21H20O11 | [M+H]+ | 449.1078 | 449.1106 | 2.8 | F |
| NV48 | 13.80 | Isorhamnetin 3-O-glucoside | C22H22O12 | [M+H]+ | 479.1184 | 479.119 | 0.6 | F |
| NV49 | 13.84 | Rosmarinic acid | C18H16O8 | [M+Na]+ | 383.0737 | 383.0732 | 0.5 | HC |
| NV50 | 14.25 | Kaempferol 3-O-arabinoside | C20H18O10 | [M+H]+ | 419.0973 | 419.0993 | 2.0 | F |
| NV51 | 14.55 | 6″-Malonylastragalin | C24H22O14 | [M+H]+ | 535.1082 | 535.1089 | 0.7 | F |
| NV52 | 14.92 | Kaempferol 3-O-alpha-L-rhamnofuranoside | C21H20O10 | [M+H]+ | 433.1129 | 433.1144 | 1.5 | F |
| NV53 | 15.35 | Kaempferol 3-O-alpha-L-arabinopyranosyl-7-O-alpha-L-rhamnopyranoside | C26H28O14 | [M+H]+ | 565.1552 | 565.1537 | 1.5 | F |
| NV54 | 15.87 | Kaempferol 3-(6-acetylgalactoside) | C23H22O12 | [M+H]+ | 491.1184 | 491.121 | 2.6 | F |
| NV55 | 16.07 | Unknown compound | - | - | - | 341.1419 | - | - |
| NV56 | 21.28 | Eryngioside F/Eryngioside H/Eryngioside I | C52H82O21 | [M+Na]+ | 1065.5241 | 1065.5248 | 0.7 | TT |
| NV57 | 21.98 | Eryngioside J | C54H84O23 | [M+Na]+ | 1123.5296 | 1123.5297 | 0.1 | TT |
| NV58 | 22.42 | Eryngioside C | C54H88O24 | [M+Na]+ | 1143.5558 | 1143.5556 | 0.2 | TT |
| NV59 | 23.35 | Eryngioside F/Eryngioside H/Eryngioside I | C52H82O21 | [M+Na]+ | 1065.5241 | 1065.5244 | 0.3 | TT |
| NV60 | 24.09 | 6-Hydroxykaempferol 3,6,7-triglucoside or other kaempferol triglucoside | C33H40O22 | [M+H-H2O]+ | 771.1984 | 771.198 | 0.4 | F |
| NV61 | 24.67 | Eryngioside K/Eryngioside L | C54H84O22 | [M+Na]+ | 1107.5346 | 1107.537 | 2.4 | TT |
| NV62 | 24.67 | Eryngioside K/Eryngioside L | C54H84O22 | [M+Na]+ | 1107.5346 | 1107.5374 | 2.8 | TT |
| NV63 | 25.12 | Eryngioside F/Eryngioside H/Eryngioside I | C52H82O21 | [M+Na]+ | 1065.5241 | 1065.5243 | 0.2 | TT |
| Sample | Amino Acids (AA), mg/100 g DW | Chlorogenic Acid (CA), mg/100 g DW | Rosmarinic Acid (RA), mg/100 g DW |
|---|---|---|---|
| EM1 W_L | 53.3 ± 1.6 | 106.0 ± 1.7 | 2.8 ± 0.4 |
| EM2 W_L | 29.5 ± 0.9 | 78.0 ± 2.1 | 2.3 ± 0.3 |
| EM1 F_L | 49.5 ± 1.5 | 51.0 ± 1.1 | 0.9 ± 0.1 |
| EM2 F_L | 67.9 ± 2.1 | 29.0 ± 0.6 | 0.9 ± 0.1 |
| EM1 F_R | 36.5 ± 1.1 | 51.0 ± 1.2 | 4.0 ± 0.5 |
| EM2 F_R | 38.1 ± 1.2 | 40.0 ± 0.9 | 2.3 ± 0.3 |
| M1_1_L | 184.0 ± 4.3 | 120.0 ± 2.0 | 259.0 ± 3.2 |
| M2_1_L | 187.6 ± 4.4 | 72.0 ± 1.2 | 181.0 ± 2.3 |
| M3_1_L | 180.5 ± 4.3 | 87.0 ± 1.5 | 266.0 ± 3.3 |
| M1_1_R | 414.2 ± 9.8 | 2360.0 ± 2.9 | 674.0 ± 8.4 |
| M2_1_R | 529.4 ± 12.6 | 2560.0 ± 2.8 | 752.0 ± 9.4 |
| M3_1_R | 846.8 ± 20.1 | 5750.0 ± 2.7 | 1202.0 ± 15.9 |
| M1_2_L | 112.6 ± 2.7 | 159.0 ± 2.3 | 3.2 ± 0.4 |
| M2_2_L | 113.1 ± 2.7 | 180.0 ± 1.8 | 4.8 ± 0.6 |
| M3_2_L | 108.4 ± 2.6 | 151.0 ± 2.0 | 4.9 ± 0.5 |
| M1_2_R | 63.1 ± 1.5 | 61.0 ± 0.8 | 3.1 ± 0.4 |
| M2_2_R | 66.0 ± 1.6 | 75.0 ± 1.2 | 3.7 ± 0.5 |
| M3_2_R | 77.3 ± 1.8 | 66.0 ± 0.9 | 4.3 ± 0.6 |
| M1_3_L | 36.4 ± 0.9 | 10.0 ± 0.3 | 0.6 ± 0.04 |
| M2_3_L | 62.3 ± 1.6 | 25.0 ± 0.7 | 1.1 ± 0.09 |
| M3_3_L | 50.8 ± 1.2 | 15.0 ± 0.4 | 1.1 ± 0.09 |
| M1_3_R | 38.3 ± 0.9 | 61.0 ± 0.9 | 4.0 ± 0.5 |
| M2_3_R | 44.5 ± 1.1 | 77.0 ± 1.3 | 6.6 ± 0.8 |
| M3_3_R | 39.0 ± 0.9 | 38.0 ± 0.7 | 2.1 ± 0.2 |
| RI * | Compound | Composition Range (Shoots) | Composition Range (Roots) |
|---|---|---|---|
| 904 | 1-Propen-2-ol acetate | n.d.–1.45 | 1.94–10.27 |
| 979 | Pentanal | n.d.–0.48 | n.d. |
| 1028 | α-Pinene | 3.42–42.34 | n.d.–9.91 |
| 1083 | Hexanal | n.d.–0.52 | n.d. |
| 1117 | β-Thujene | 0.95–5.19 | n.d. |
| 1161 | β-Myrcene | 4.97–17.76 | n.d.–1.19 |
| 1167 | α -Phellandrene | n.d.–0.10 | n.d.–0.98 |
| 1202 | 1-Pentanol | n.d.–0.85 | n.d. |
| 1207 | Limonene | 2.1–7.87 | n.d. |
| 1211 | β-Phellandrene | n.d.–0.38 | n.d. |
| 1246 | γ-Terpinene | n.d.–0.43 | n.d. |
| 1249 | o-Cymene | n.d.–0.47 | n.d. |
| 1251 | Mesitylene | 3.82–15.43 | 4.1–11.6 |
| 1256 | 3,4-Dimethylstyrene | n.d.–0.84 | n.d. |
| 1340 | Hemimellitene | 0.41–2.81 | 0.82–2.71 |
| 1406 | 1,3,5,5-Tetramethyl-1,3-cyclohexadiene | 0.34–0.89 | n.d.–1.01 |
| 1462 | trans-Limonene oxide | n.d.–0.17 | n.d. |
| 1582 | Methyl citronellate | 0.05–0.31 | n.d. |
| 1616 | Safranal | 0.31–2.29 | n.d.–0.78 |
| 1640 | Benzeneacetaldehyde | n.d.–0.37 | n.d. |
| 1663 | cis-Verbanol | 0.15–0.75 | n.d. |
| 1685 | p-Mentha-1,5-dien-8-ol | n.d.–0.21 | n.d. |
| 1687 | trans-Verbenol | 0.56–3.16 | n.d. |
| 1689 | γ-Selinene | 0.25–0.51 | n.d. |
| 1695 | Cadinene | 2.68–5.91 | 3.98–15.58 |
| 1710 | Germacrene D | n.d.–3.28 | n.d. |
| 1729 | Valencene I | 9.47–19.85 | 2.25–21.67 |
| 1732 | Valencene II | n.d.–0.77 | n.d.–0.78 |
| 1792 | Selina-3,7(11)-diene | n.d.–0.16 | n.d.–0.51 |
| 1822 | 2-Methyl-2-buteonic acid | n.d.–7.20 | n.d.–2.12 |
| 1875 | Thymoquinone | n.d.–0.80 | n.d.–1.19 |
| 1896 | Duraldehyde | 1.38–8.99 | 11.16–29.61 |
| 1929 | Mesitaldehyde | 6.74–29.17 | 33.77–48.67 |
| Sum of unidentified | 0.89–10.45 | 0–27.15 |
| Sample Code | Plant Part | Growth Conditions | Population | Sample Type |
|---|---|---|---|---|
| EM1 W_L | Leaves | Wild | EM1 | Pooled sample of 20 individuals |
| EM2 W_L | Leaves | EM2 | ||
| EM1 F_L | Leaves | Field-grown, first vegetation season | EM1 | Pooled sample of 20 individuals |
| EM2 F_L | Leaves | EM2 | ||
| EM1 F_R | Roots | EM1 | ||
| EM2 F_R | Roots | EM2 | ||
| M1_1_L | Shoot | In vitro | EM2 | Sample of individual mericlones |
| M2_1_L | Shoot | |||
| M3_1_L | Shoot | |||
| M1_1_R | Roots | |||
| M2_1_R | Roots | |||
| M3_1_R | Roots | |||
| M1_2_L | Leaves | Field-grown, first vegetation season | EM2 | Sample of individual mericlones |
| M2_2_L | Leaves | |||
| M3_2_L | Leaves | |||
| M1_2_R | Roots | |||
| M2_2_R | Roots | |||
| M3_2_R | Roots | |||
| M1_3_L | Leaves | Field-grown, second vegetation season | EM2 | Sample of individual mericlones |
| M2_3_L | Leaves | |||
| M3_3_L | Leaves | |||
| M1_3_R | Roots | |||
| M2_3_R | Roots | |||
| M3_3_R | Roots |
| Compound | Class | Purity | Mass, mg | Volume, mL | Stock Solution, mg/mL | Calibration Range |
|---|---|---|---|---|---|---|
| Leucine | Amino acids | 99% | 4.5 | 10 mL water | 0.45 | 0.1–10 μg/mL R2 = 0.998 |
| Chlorogenic acid | Hydroxycinnamic acids | 98% | 10.1 | 10 mL methanol | 1.01 | 0.1–100 μg/mL R2 = 0.998 |
| Rosmarinic acid | Hydroxycinnamic acids | 98% | 9.8 | 10 mL methanol | 0.98 | 0.1–100 µg/mL R2 = 0.993 |
| Carried Test | Procedure | Wavelength | Standard and Concentration Range |
|---|---|---|---|
| Total phenolic content (TPC) | 25 µL of extract was mixed with 75 µL of H2O and 25 µL of Folin–Ciocalteu reagent (1:10) for 6 min. 100 µL of a 7% Na2CO3 solution was added; the plate was shaken for 30 s and left in a dark place at room temperature for 90 min. | 765 nm | 0.025–0.20 mg mL−1 gallic acid solutions |
| Total saponin content (TSC) | 20 µL of the extract was mixed with 20 µL of 0.5% anisaldehyde (diluted with ethyl acetate) and allowed to react for 10 min in a dark place. Afterwards, 200 µL of 72% sulfuric acid was added and allowed to react for color development at 60 °C for 10 min. | 560 nm | 0.10–0.40 mg mL−1 escin solutions |
| Total sugar content (Sugars) | 50 µL of extract was mixed with 150 µL of H2SO4 and 30 µL of 5% phenol reagent before being heated in an oven at 90 °C for 5 min. After heating, the plates were cooled. | 490 nm | 0.045–0.90 mg mL−1 glucose solutions |
| Antiradical activity/ DPPH radical scavenging (ARA/DPPH) | 20 µL of extract was mixed with 180 µL of 150 µM DPPH reagent. The plate was kept in the dark at room temperature for 60 min. Different concentrations of the extract were tested to find the IC50, which is the concentration at which the absorbance of DPPH dropped by 50%. | 517 nm | 0.018–0.22 μg mL−1 ascorbic acid solutions |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakurte, I.; Berga, M.; Mežaka, I. Phytochemical Diversity Comparison in Leaves and Roots of Wild and Micropropagated Latvian Sea Holly (Eryngium maritimum L.). Molecules 2023, 28, 3924. https://doi.org/10.3390/molecules28093924
Nakurte I, Berga M, Mežaka I. Phytochemical Diversity Comparison in Leaves and Roots of Wild and Micropropagated Latvian Sea Holly (Eryngium maritimum L.). Molecules. 2023; 28(9):3924. https://doi.org/10.3390/molecules28093924
Chicago/Turabian StyleNakurte, Ilva, Marta Berga, and Ieva Mežaka. 2023. "Phytochemical Diversity Comparison in Leaves and Roots of Wild and Micropropagated Latvian Sea Holly (Eryngium maritimum L.)" Molecules 28, no. 9: 3924. https://doi.org/10.3390/molecules28093924