

Characterization of the Key Compounds of Bell Pepper by Spectrophotometry and Gas Chromatography on the Effects of Induced Stress on the Concentration of Secondary Metabolite
 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Metabolomic Profiling of Sweet Pepper Red
2.2. The Phenolic Profile of Sweet Pepper
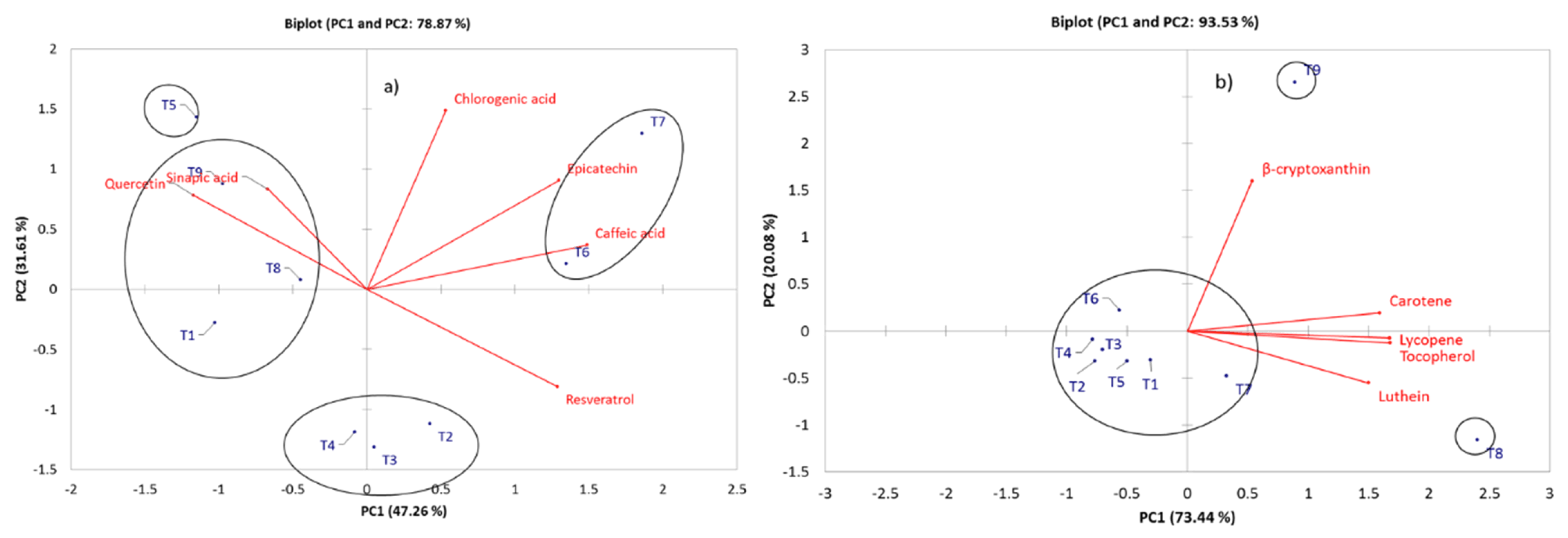
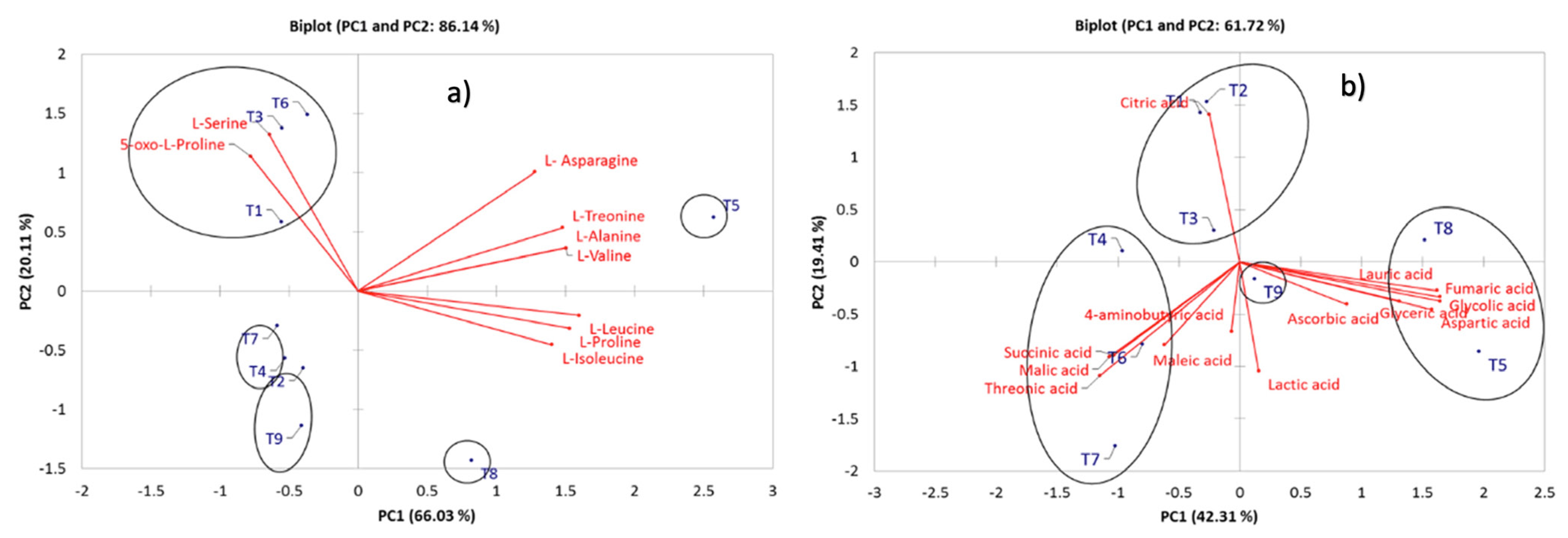
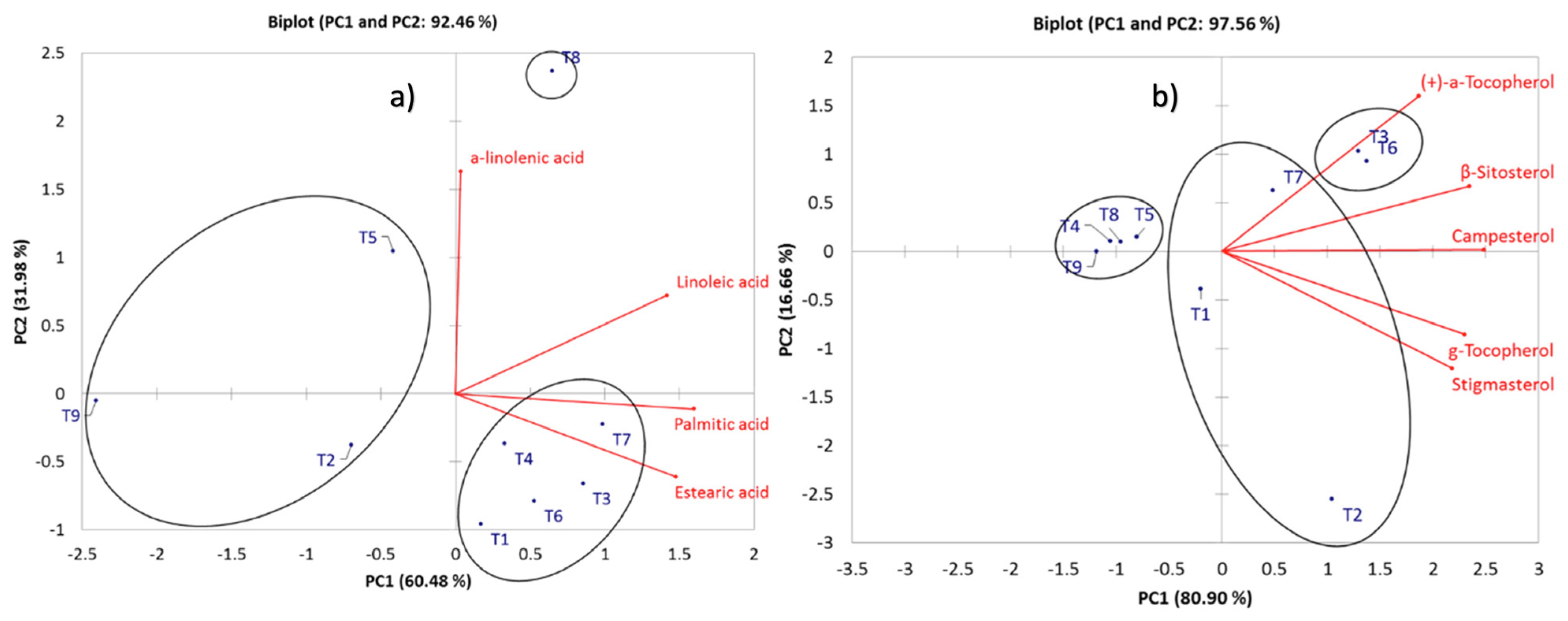
2.3. Comparative Profiling of Metabolites in the Different Treatments under Elicitor Application and Electric Conductivity Levels by PCA and HCA
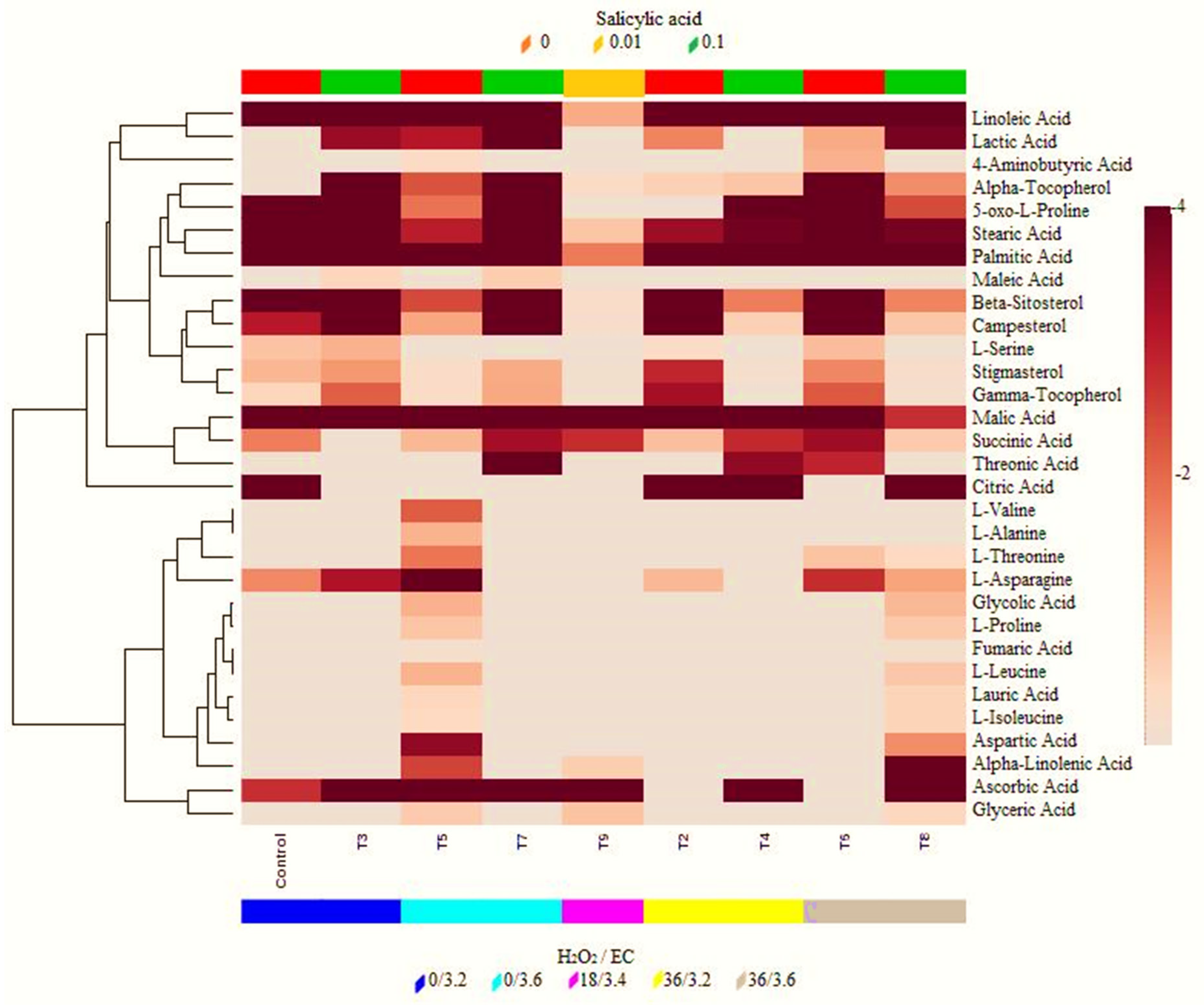
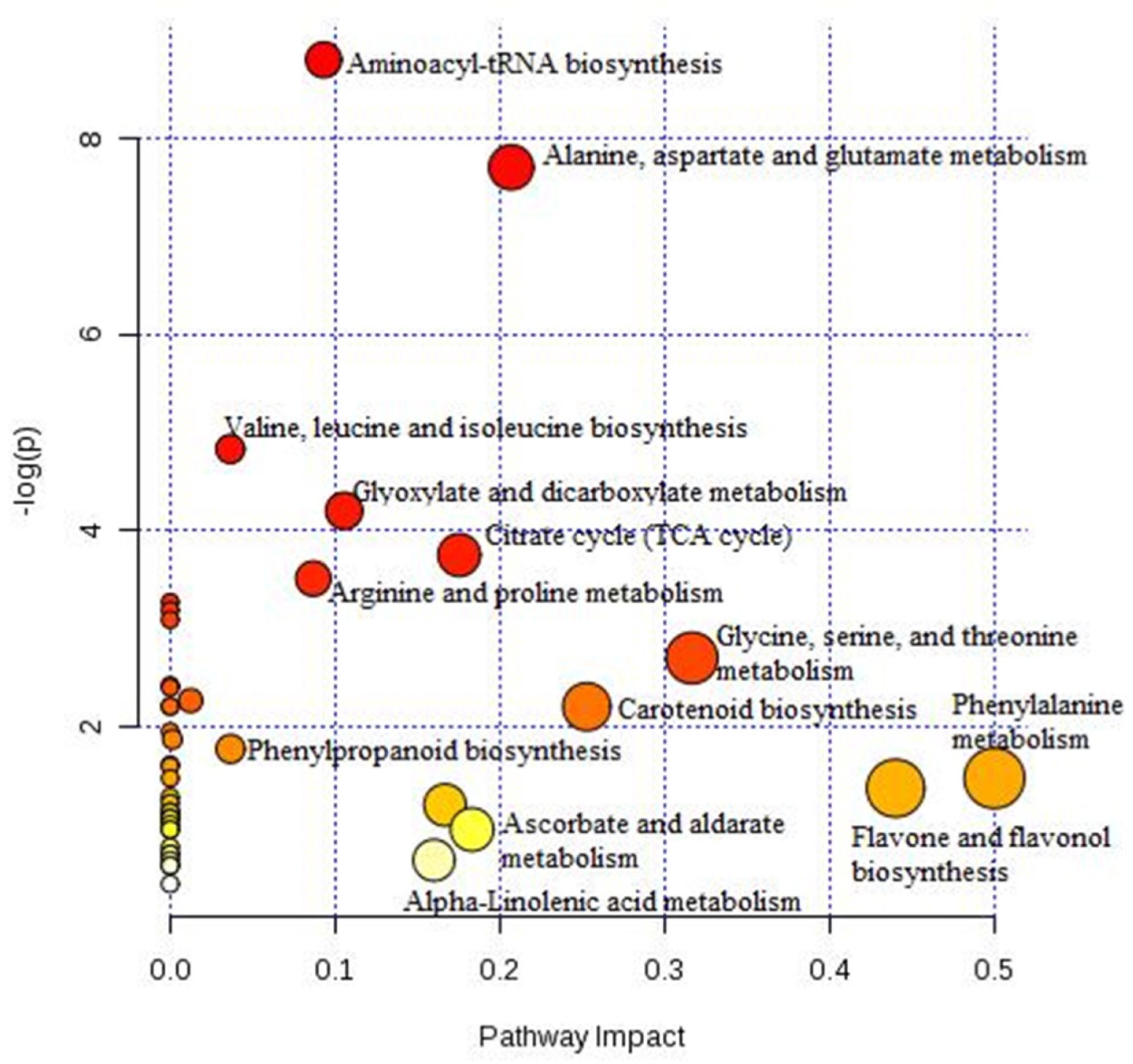
2.4. Metabolic Profiles of Sweet Pepper (C. annum L.) Stress-Induced
3. Discussion
4. Materials and Methods
4.1. Greenhouse Climatic Conditions
4.2. Experimental Design
4.3. Fruit Material and Preparation
4.4. Identification of Phenolic Compounds
4.5. Identification of Carotenoids
4.6. Identification of Ascorbic Acid
4.7. Gas-Chromatography-Mass Spectrometry (GC-MS) Analysis
4.8. Statistical Analysis
4.9. Hierarchical Clustering and Pathway Enrichment Analysis
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Forest Europe; UNECE FAO; State of Europe’s Forests. Status and Trends in Sustainable Forest Management in Europe. In Proceedings of the Ministerial Conference on the Protection of Forests in Europe, Oslo, Norway, 14–16 June 2011; p. 344. [Google Scholar]
- Howard, L.R.; Wildman, R.E. Antioxidant vitamin and phytochemical content of fresh and processed pepper fruit (Capsicum annuum). In Nutraceuticals and Funtional Foods, 2nd ed.; Wildman, R.E.C., Ed.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA; New York, NY, USA, 2007; pp. 165–192. [Google Scholar]
- Rybak, K.; Samborska, K.; Jedlinska, A.; Parniakov, O.; Nowacka, M.; Witrowa-Rajchert, D.; Wiktor, A. The impact of pulsed electric field pretreatment of bell pepper on the selected properties of spray dried juice. Innov. Food Sci. Emerg. Technol. 2020, 65, 102446. [Google Scholar] [CrossRef]
- Wahyuni, Y.; Ballester, A.-R.; Sudarmonowati, E.; Bino, R.J.; Bovy, A.G. Metabolite biodiversity in pepper (Capsicum) fruits of thirty-two diverse accessions: Variation in health-related compounds and implications for breeding. Phytochemistry 2011, 72, 1358–1370. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.D.; Kennedy, J.A. Plant metabolism and the environment: Implications for managing phenolics. Crit. Rev. Food Sci. Nutr. 2010, 50, 620–643. [Google Scholar] [CrossRef]
- Sadef, Y.; Javed, T.; Javed, R.; Mahmood, A.; Alwahibi, M.S.; Elshikh, M.S.; AbdelGawwa, M.R.; Alhaji, J.H.; Rasheed, R.A. Nutritional status, antioxidant activity and total phenolic content of different fruits and vegetables’ peels. PLoS ONE 2022, 17, e0265566. [Google Scholar] [CrossRef]
- Martínez-Ispizua, E.; Martínez-Cuenca, M.-R.; Marsal, J.I.; Díez, M.J.; Soler, S.; Valcárcel, J.V.; Calatayud, Á. Bioactive compounds and antioxidant capacity of valencian pepper landraces. Molecules 2021, 26, 1031. [Google Scholar] [CrossRef] [PubMed]
- Edreva, A.; Velikova, V.; Tsonev, T.; Dagnon, S.; Gürel, A.; Aktaş, L.; Gesheva, E. Stress-protective role of secondary metabolites: Diversity of functions and mechanisms. Gen. Appl. Plant Physiol. 2008, 34, 67–78. [Google Scholar]
- Ferrari, S. Biological elicitors of plant secondary metabolites: Mode of action and use in the production of nutraceutics. In Bio-Farms for Nutraceuticals; Springer: Berlin/Heidelberg, Germany, 2010; pp. 152–166. [Google Scholar]
- Tang, Y.; Zhang, G.; Yang, T.; Yang, S.; Aisimutuola, P.; Wang, B.; Li, N.; Wang, J.; Yu, Q. Biochemical variances through metabolomic profile analysis of Jacq. during fruit development. Folia Hortic. 2021, 33, 17–26. [Google Scholar] [CrossRef]
- Batiha, G.E.-S.; Alqahtani, A.; Ojo, O.A.; Shaheen, H.M.; Wasef, L.; Elzeiny, M.; Ismail, M.; Shalaby, M.; Murata, T.; Zaragoza-Bastida, A. Biological properties, bioactive constituents, and pharmacokinetics of some Capsicum spp. and capsaicinoids. Int. J. Mol. Sci. 2020, 21, 5179. [Google Scholar] [CrossRef]
- Raskin, I. Role of salicylic acid in plants. Annu. Rev. Plant Biol. 1992, 43, 439–463. [Google Scholar] [CrossRef]
- Barrajon-Catalan, E.; Álvarez-Martínez, F.J.; Borras, F.; Perez, D.; Herrero, N.; Ruiz, J.J.; Micol, V. Metabolomic analysis of the effects of a commercial complex biostimulant on pepper crops. Food Chem. 2020, 310, 125818. [Google Scholar] [CrossRef]
- Klessig, D.F.; Malamy, J. The salicylic acid signal in plants. In Signals and Signal Transduction Pathways in Plants; Springer: Berlin/Heidelberg, Germany, 1994; pp. 203–222. [Google Scholar]
- Cheeseman, J.M. Hydrogen peroxide and plant stress: A challenging relationship. Plant Stress 2007, 1, 4–15. [Google Scholar]
- Hung, S.-H.; Yu, C.-W.; Lin, C.H. Hydrogen peroxide functions as a stress signal in plants. Bot. Bull. Acad. Sin. 2005, 46, 1–10. [Google Scholar]
- Dorai, M.; Papadopoulos, A.; Gosselin, A. Influence of electric conductivity management on greenhouse tomato yield and fruit quality. Agronomie 2001, 21, 367–383. [Google Scholar] [CrossRef]
- Al-Rubeai, M.; Fussenegger, M. Systems Biology; Springer: Berlin/Heidelberg, Germany, 2007; Volume 5. [Google Scholar]
- Liu, D.; He, X.-Q.; Wu, D.-T.; Li, H.-B.; Feng, Y.-B.; Zou, L.; Gan, R.-Y. Elderberry (Sambucus nigra L.): Bioactive Compounds, Health Functions, and Applications. J. Agric. Food Chem. 2022, 70, 4202–4220. [Google Scholar] [CrossRef] [PubMed]
- Arbona, V.; Manzi, M.; Ollas, C.d.; Gómez-Cadenas, A. Metabolomics as a tool to investigate abiotic stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 4885–4911. [Google Scholar] [CrossRef]
- Teixeira, A.; Martins, V.; Noronha, H.; Eiras-Dias, J.; Gerós, H. The First Insight into the Metabolite Profiling of Grapes from Three Vitis vinifera L. Cultivars of Two Controlled Appellation (DOC) Regions. Int. J. Mol. Sci. 2014, 15, 4237–4254. [Google Scholar] [CrossRef]
- Ananthan, R.; Subhash, K.; Longvah, T. Capsaicinoids, amino acid and fatty acid profiles in different fruit components of the world hottest Naga king chilli (Capsicum chinense Jacq). Food Chem. 2018, 238, 51–57. [Google Scholar] [CrossRef]
- Guzman, I.; Coon, D.; Vargas, K.; Bosland, P.W. NuMex LotaLutein, a Lutein-rich Serrano Pepper. HortScience 2020, 55, 2052–2055. [Google Scholar] [CrossRef]
- Fernie, A.R.; Schauer, N. Metabolomics-assisted breeding: A viable option for crop improvement? Trends Genet. 2009, 25, 39–48. [Google Scholar] [CrossRef]
- Florentino-Ramos, E.; Villa-Ruano, N.; Hidalgo-Martínez, D.; Ramírez-Meraz, M.; Méndez-Aguilar, R.; Velásquez-Valle, R.; Zepeda-Vallejo, L.G.; Pérez-Hernández, N.; Becerra-Martínez, E. 1H NMR-based fingerprinting of eleven Mexican Capsicum annuum cultivars. Food Res. Int. 2019, 121, 12–19. [Google Scholar] [CrossRef]
- Feng, X.; Yu, Q.; Li, B.; Kan, J. Comparative analysis of carotenoids and metabolite characteristics in discolored red pepper and normal red pepper based on non-targeted metabolomics. LWT 2022, 153, 112398. [Google Scholar] [CrossRef]
- Rhouma, A.; Ben Daoud, H.; Ghanmi, S.; Ben Salah, H.; Romdhane, M.; Demak, M. Antimicrobial activities of leaf extracts of Pistacia and Schinus species against some plant pathogenic fungi and bacteria. J. Plant Pathol. 2009, 91, 339–345. [Google Scholar]
- Iorizzi, M.; Lanzotti, V.; De Marino, S.; Zollo, F.; Blanco-Molina, M.; Macho, A.; Muñoz, E. New glycosides from Capsicum annuum L. var. acuminatum. Isolation, structure determination, and biological activity. J. Agric. Food Chem. 2001, 49, 2022–2029. [Google Scholar] [CrossRef] [PubMed]
- Campos, Â.D.; Ferreira, A.G.; Hampe, M.M.V.; Antunes, I.F.; Brancão, N.; Silveira, E.P.; Silva, J.B.d.; Osório, V.A. Induction of chalcone synthase and phenylalanine ammonia-lyase by salicylic acid and Colletotrichum lindemuthianum in common bean. Braz. J. Plant Physiol. 2003, 15, 129–134. [Google Scholar] [CrossRef]
- Sakurai, N. Recent applications of metabolomics in plant breeding. Breed. Sci. 2022, 72, 56–65. [Google Scholar] [CrossRef]
- Jeong, W.Y.; Jin, J.S.; Cho, Y.A.; Lee, J.H.; Park, S.; Jeong, S.W.; Kim, Y.H.; Lim, C.S.; El-Aty, A.A.; Kim, G.S. Determination of polyphenols in three Capsicum annuum L.(bell pepper) varieties using high-performance liquid chromatography-tandem mass spectrometry: Their contribution to overall antioxidant and anticancer activity. J. Sep. Sci. 2011, 34, 2967–2974. [Google Scholar] [CrossRef]
- Silva, C.L.; Haesen, N.; Câmara, J.S. A new and improved strategy combining a dispersive-solid phase extraction-based multiclass method with ultra high pressure liquid chromatography for analysis of low molecular weight polyphenols in vegetables. J. Chromatogr. 2012, 1260, 154–163. [Google Scholar] [CrossRef]
- Mokhtar, M.; Soukup, J.; Donato, P.; Cacciola, F.; Dugo, P.; Riazi, A.; Jandera, P.; Mondello, L. Determination of the polyphenolic content of a Capsicum annuum L. extract by liquid chromatography coupled to photodiode array and mass spectrometry detection and evaluation of its biological activity. J. Sep. Sci. 2015, 38, 171–178. [Google Scholar] [CrossRef]
- Guzman, I.; Vargas, K.; Chacon, F.; McKenzie, C.; Bosland, P.W. Health-promoting carotenoids and phenolics in 31 Capsicum accessions. HortScience 2021, 56, 36–41. [Google Scholar] [CrossRef]
- Medina-Juárez, L.; Molina-Quijada, D.M.; Del Toro-Sánchez, C.L.; González-Aguilar, G.A.; Gámez-Meza, N. Antioxidant activity of peppers (Capsicum annuum L.) extracts and characterization of their phenolic constituents. Interciencia 2012, 37, 588–593. [Google Scholar]
- Biacs, P.A.; Daood, H.G.; Pavisa, A.; Hajdu, F. Studies on the carotenoid pigments of paprika (Capsicum annuum L. var Sz-20). J. Agric. Food Chem. 1989, 37, 350–353. [Google Scholar] [CrossRef]
- Deli, J.; Matus, Z.; Szabolcs, J. Carotenoid composition in the fruits of black paprika (Capsicum annuum variety longum nigrum) during ripening. J. Agric. Food Chem. 1992, 40, 2072–2076. [Google Scholar] [CrossRef]
- Machlin, L.J.; Bendich, A. Free radical tissue damage: Protective role of antioxidant nutrients. FASEB J. 1987, 1, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Bendich, A.; Olson, J.A. Biological actions of carotenoids. FASEB J. 1989, 3, 1927–1932. [Google Scholar] [CrossRef]
- Lozada, D.N.; Bhatta, M.; Coon, D.; Bosland, P.W. Single nucleotide polymorphisms reveal genetic diversity in New Mexican chile peppers (Capsicum spp.). BMC Genom. 2021, 22, 1–12. [Google Scholar] [CrossRef]
- Howard, L.; Smith, R.; Wagner, A.; Villalon, B.; Burns, E. Provitamin A and ascorbic acid content of fresh pepper cultivars (Capsicum annuum) and processed jalapenos. J. Food Sci. 1994, 59, 362–365. [Google Scholar] [CrossRef]
- Simonne, A.; Simonne, E.; Eitenmiller, R.; Mills, H.; Green, N. Ascorbic Acid and Provitamin A Contents in Unusually Colored Bell Peppers (Capsicum annuum L.). J. Food Compos. Anal. 1997, 10, 299–311. [Google Scholar] [CrossRef]
- Storey, R.; Davies, H. Tuber quality. In The Potato Crop; Harris, P., Ed.; Springer: Berlin/Heidelberg, Germany; Champam & Hall: London, UK, 1992; pp. 507–569. [Google Scholar]
- Zhang, D.; Hamauzu, Y. Phenolics, ascorbic acid, carotenoids and antioxidant activity of broccoli and their changes during conventional and microwave cooking. Food Chem. 2004, 88, 503–509. [Google Scholar] [CrossRef]
- Hancock, R.E.; Sahl, H.-G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef]
- Foyer, C.H.; Lopez-Delgado, H.; Dat, J.F.; Scott, I.M. Hydrogen peroxide and glutathione associated mechanisms of acclimatory stress tolerance and signalling. Physiol. Plant. 1997, 100, 241–254. [Google Scholar] [CrossRef]
- Rivas-San Vicente, M.; Plasencia, J. Salicylic acid beyond defence: Its role in plant growth and development. J. Exp. Bot. 2011, 62, 3321–3338. [Google Scholar] [CrossRef] [PubMed]
- Ghasemzadeh, A.; Omidvar, V.; Jaafar, H.Z. Polyphenolic content and their antioxidant activity in leaf extract of sweet potato (Ipomoea batatas). J. Med. Plants Res. 2012, 6, 2971–2976. [Google Scholar] [CrossRef]
- Ibrahim, M.H.; Jaafar, H.Z.; Rahmat, A.; Rahman, Z.A. Effects of nitrogen fertilization on synthesis of primary and secondary metabolites in three varieties of kacip fatimah (Labisia Pumila Blume). Int. J. Mol. Sci. 2011, 12, 5238–5254. [Google Scholar] [CrossRef] [PubMed]
- Espichán, F.; Rojas, R.; Quispe, F.; Cabanac, G.; Marti, G. Metabolomic characterization of 5 native Peruvian chili peppers (Capsicum spp.) as a tool for species discrimination. Food Chem. 2022, 386, 132704. [Google Scholar] [CrossRef] [PubMed]
- Kaewnaree, P.; Vichitphan, S.; Klanrit, P.; Siri, B.; Vichitphan, K. Effect of accelerated aging process on seed quality and biochemical changes in sweet pepper (Capsicum annuum Linn.) seeds. Biotechnology 2011, 10, 175–182. [Google Scholar] [CrossRef]
- Rahimi, A.R.; Rokhzadi, A.; Amini, S.; Karami, E. Effect of salicylic acid and methyl jasmonate on growth and secondary metabolites in Cuminum cyminum L. J. Biodivers. Environ. Sci. 2013, 3, 140–149. [Google Scholar]
- Dučaiová, Z.; Petruľová, V.; Repčák, M. Salicylic acid regulates secondary metabolites content in leaves of Matricaria chamomilla. Biologia 2013, 68, 904–909. [Google Scholar] [CrossRef]
- Pech, R.; Volná, A.; Hunt, L.; Bartas, M.; Červeň, J.; Pečinka, P.; Špunda, V.; Nezval, J. Regulation of Phenolic Compound Production by Light Varying in Spectral Quality and Total Irradiance. Int. J. Mol. Sci. 2022, 23, 6533. [Google Scholar] [CrossRef]
- Hao, W.; Guo, H.; Zhang, J.; Hu, G.; Yao, Y.; Dong, J. Hydrogen Peroxide Is Involved in Salicylic Acid-Elicited Rosmarinic Acid Production in Salvia miltiorrhiza Cell Cultures. Sci. World J. 2014, 2014, 843764. [Google Scholar] [CrossRef]
- Desikan, R.; Cheung, M.K.; Bright, J.; Henson, D.; Hancock, J.T.; Neill, S.J. ABA, hydrogen peroxide and nitric oxide signalling in stomatal guard cells. J. Exp. Bot. 2004, 55, 205–212. [Google Scholar] [CrossRef]
- Wendehenne, D.; Durner, J.; Klessig, D.F. Nitric oxide: A new player in plant signalling and defence responses. Curr. Opin. Plant Biol. 2004, 7, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Leete, E.; Louden, M.C. Biosynthesis of capsaicin and dihydrocapsaicin in Capsicum frutescens. J. Am. Chem. Soc. 1968, 90, 6837–6841. [Google Scholar] [CrossRef] [PubMed]
- Kopp, B.; Jurenitsch, J. Biosynthesis of Capsaicinoids in Capsicum annuum L. var. annuum. Planta Med. 1981, 43, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085. [Google Scholar] [CrossRef]
- Reddy, U.K.; Almeida, A.; Abburi, V.L.; Alaparthi, S.B.; Unselt, D.; Hankins, G.; Park, M.; Choi, D.; Nimmakayala, P. Identification of gene-specific polymorphisms and association with capsaicin pathway metabolites in Capsicum annuum L. collections. PLoS ONE 2014, 9, e86393. [Google Scholar] [CrossRef]
- Zeinali, A.; Esmaeili, M.; Heidarzade, A. Salicylic acid and abiotic stress influence allelochemicals and inhibitory potential of root exudates of two rice (Oryza sativa) cultivars against Barnyardgrass (Echinochloa crus-galli L.). Int. J. Farm. Allied Sci. 2013, 2, 779–784. [Google Scholar]
- Torres-Schumann, S.; Godoy, J.A.; Pintor-Toro, J.A. A probable lipid transfer protein gene is induced by NaCl in stems of tomato plants. Plant Mol. Biol. 1992, 18, 749–757. [Google Scholar] [CrossRef]
- Molina, A.; Segura, A.; García-Olmedo, F. Lipid transfer proteins (nsLTPs) from barley and maize leaves are potent inhibitors of bacterial and fungal plant pathogens. FEBS Lett. 1993, 316, 119–122. [Google Scholar] [CrossRef]
- Treviño, M.B.; O’Connell, M.A. Three Drought-Responsive Members of the Nonspecific Lipid-Transfer Protein Gene Family in Lycopersicon pennellii Show Different Developmental Patterns of Expression. Plant Physiol. 1998, 116, 1461–1468. [Google Scholar] [CrossRef]
- Terras, F.R.; Goderis, I.J.; Van Leuven, F.; Vanderleyden, J.; Cammue, B.P.; Broekaert, W.F. In vitro antifungal activity of a radish (Raphanus sativus L.) seed protein homologous to nonspecific lipid transfer proteins. Plant Physiol. 1992, 100, 1055–1058. [Google Scholar] [CrossRef]
- Jung, H.W.; Kim, W.; Hwang, B.K. Three pathogen-inducible genes encoding lipid transfer protein from pepper are differentially activated by pathogens, abiotic, and environmental stresses*. Plant Cell. Environ. 2003, 26, 915–928. [Google Scholar] [CrossRef] [PubMed]
- Angmo, P.; Dolma, T.; Phuntsog, N.; Chaurasia, O.; Stobdan, T. Effect of shading and high temperature amplitude on yield and phenolic contents of greenhouse capsicum (Capsicum annuum L.). J. Biol. Pharm. 2021, 4, 30–39. [Google Scholar]
- Blée, E. Impact of phyto-oxylipins in plant defense. Trends Plant Sci. 2002, 7, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Upchurch, R.G. Fatty acid unsaturation, mobilization, and regulation in the response of plants to stress. Biotechnol. Lett. 2008, 30, 967–977. [Google Scholar] [CrossRef]
- Xue, H.; Upchurch, R.; Kwanyuen, P. Ergosterol as a Quantifiable Biomass Marker for Diaporthe haseolorum and Cercospora kikuchi. Plant Dis. 2006, 90, 1395–1398. [Google Scholar] [CrossRef] [PubMed]
- Wahyuni, Y.; Stahl-Hermes, V.; Ballester, A.-R.; de Vos, R.C.; Voorrips, R.E.; Maharijaya, A.; Molthoff, J.; Zamora, M.V.; Sudarmonowati, E.; Arisi, A.C.M. Genetic mapping of semi-polar metabolites in pepper fruits (Capsicum sp.): Towards unravelling the molecular regulation of flavonoid quantitative trait loci. Mol. Breed. 2014, 33, 503–518. [Google Scholar] [CrossRef]
- Gzik, A. Accumulation of proline and pattern of α-amino acids in sugar beet plants in response to osmotic, water and salt stress. Environ. Exp. Bot. 1996, 36, 29–38. [Google Scholar] [CrossRef]
- Castillejo, N.; Martínez-Zamora, L.; Artés-Hernández, F. Postharvest UV radiation enhanced biosynthesis of flavonoids and carotenes in bell peppers. Postharvest Biol. Technol. 2022, 184, 111774. [Google Scholar] [CrossRef]
- Huang, G.-T.; Ma, S.-L.; Bai, L.-P.; Zhang, L.; Ma, H.; Jia, P.; Liu, J.; Zhong, M.; Guo, Z.-F. Signal transduction during cold, salt, and drought stresses in plants. Mol. Biol. Rep. 2012, 39, 969–987. [Google Scholar] [CrossRef]
- Prasch, C.M.; Sonnewald, U. Signaling events in plants: Stress factors in combination change the picture. Environ. Exp. Bot. 2015, 114, 4–14. [Google Scholar] [CrossRef]
- Mendoza Robles, J.; Laird, R.; Turrent Fernandez, A.; Trinidad Santos, A. [Generation of recommendations on the fertilization of rainfed maize in middle South, Morelos region]. Agrociencia 1985, 60, 75–86. [Google Scholar]
- Cortés, M.; Herrera, E.; Rodríguez, E. Optimización experimental del proceso de liofilización de uchuva adicionada con componentes activos por impregnación al vacío. Vitae 2015, 22, 47–56. [Google Scholar] [CrossRef]
- Cardador-Martínez, A.; Loarca-Piña, G.; Oomah, B.D. Antioxidant activity in common beans (Phaseolus vulgaris L.). J. Agric. Food Chem. 2002, 50, 6975–6980. [Google Scholar] [CrossRef]
- Ruiz-Cruz, S.; Alvarez-Parrilla, E.; Laura, A.; Martinez-Gonzalez, A.I.; Ornelas-Paz, J.D.; Mendoza-Wilson, A.M.; Gonzalez-Aguilar, G.A. Effect of different sanitizers on microbial, sensory and nutritional quality of fresh-cut jalapeno peppers. Am. J. Agric. Biol. Sci. 2010, 5, 331. [Google Scholar] [CrossRef]
- Ni, Y.; Su, M.; Lin, J.; Wang, X.; Qiu, Y.; Zhao, A.; Chen, T.; Jia, W. Metabolic profiling reveals disorder of amino acid metabolism in four brain regions from a rat model of chronic unpredictable mild stress. FEBS Lett. 2008, 582, 2627–2636. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Compound (Groups) | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | F (F-Ratio) | p (p-Values) |
| Phenolic compounds | |||||||||||
| Chlorogenic acid | - | - | - | - | 2746.79 a | 2414.41 a | 4229.29 b | 2376.57 a | 2538.03 a | 214.88 | 0.000 |
| Epicatechin | - | - | - | - | 577.30 a | 3989.41 a | 8896.08 b | - | 1052.83 a | 17.69 | 0.000 |
| Caffeic acid | - | 717.21ab | - | - | - | 1353.34 b | 1239.91 b | - | - | 10.439 | 0.001 |
| Sinapic acid | - | - | - | - | 98.59 a | - | - | - | - | 2.326 | 0.115 |
| Quercetin | 175.02 a | 71.09 a | 73.53 a | 77.32 a | 159.95 a | 61.04 a | 82.79 a | 85.45 a | 229.73 a | 2.772 | 0.075 |
| Resveratrol | - | 180.42 b | 197.42 b | 161.74 ab | - | 158.13 ab | 177.85 b | - | - | 8.474 | 0.002 |
| Carotenoids | |||||||||||
| Luthein | 0.0017 ab | 0.0015 ab | 0.0015 ab | 0.0013 ab | 0.0016 ab | 0.0009 a | 0.0019 b | 0.0029 c | 0.0017 ab | 7.965 | 0.000 |
| Tocopherol | 0.0075 b | 0.0072 b | 0.0074 b | 0.0074 b | 0.0082 b | 0.0081 b | 0.0097 b | 0.0173 a | 0.0114 b | 7.214 | 0.000 |
| β-cryptoxanthin | 0.0012 a | 0.0012 a | 0.0012 a | 0.0012 a | 0.0012 a | 0.0012 a | 0.0012 a | 0.0012 a | 0.0015 a | 1.074 | 0.398 |
| Lycopene | 0.0060 a | 0.0057a | 0.0018 a | 0.0027 a | 0.0048 a | 0.0057 a | 0.0115 ab | 0.0227 b | 0.0128 ab | 5.178 | 0.000 |
| Carotene | 0.0149 ab | 0.0102a | 0.0133 ab | 0.0129 a | 0.0127 a | 0.0160 ab | 0.0164 ab | 0.0230 b | 0.0190 ab | 3.256 | 0.005 |
| Vitamins | |||||||||||
| Ascorbic acid | 0.84 a | 1.12 b | 1.09 b | 0.85 a | 1.13 b | 0.96 ab | 1.12 b | 1.31 c | 0.98 ab | 0.731 | 0.664 |
| Aminoacids | |||||||||||
| L-Alanine | - | - | - | - | 1.087 a | - | - | - | 0.000 | 0.000 | |
| L-Valine | - | - | - | - | 2.167 a | - | - | - | 0.000 | 0.000 | |
| L-Leucine | - | - | - | - | 1.07 a | - | - | 0.771 a | - | 0.021 | 0.281 |
| L-Isoleucine | - | - | - | - | 0.441 a | - | - | 0.553 a | - | 0.008 | 0.000 |
| L-Proline | - | - | - | - | 0.796 a | - | - | 0.731 a | - | 0.724 | 0.001 |
| L-Serine | 0.84 b | 0.317 a | 1.094 c | - | 0.908 b | - | 0.963 | 0.000 | |||
| L-Treonine | - | - | - | - | 1.845 c | 0.834 b | 0.454 a | - | 0.384 | 0.000 | |
| 5-oxo-L-Proline | 10.996 b | 16.181 bc | 10.517 b | 1.887 a | 18.465 c | 15.526 bc | 2.399 a | - | 0.551 | 0.391 | |
| L- Asparagine | 1.63 c | 0.982 bc | 3.181 a | 6.612 b | 2.757 a | 1.325 a | - | 1.522 | 0.000 | ||
| Organic acids | |||||||||||
| Lactic acid | - | 1.709 a | 3.423 ab | 3.083 ab | 1.207 a | 7.663 b | 3.843 a | - | 1.083 | 0.000 | |
| Glycolic acid | - | - | - | - | 1.103 a | - | - | 0.977 a | - | 0.774 | 0.000 |
| Succinic acid | 1.751 b | 0.893 ab | 2.827 a | 0.988 ab | 3.366 a | 3.284 a | 0.691 a | 2.753 a | 1.691 | 0.000 | |
| Glyceric acid | - | - | - | - | 0.696 a | - | - | 0.487 a | 0.838 a | 0.478 | 0.000 |
| Fumaric acid | - | - | - | - | 0.184 a | - | - | 0.136 a | - | 0.096 | 0.291 |
| Maleic acid | - | - | 0.529 a | - | - | - | 0.671 a | - | - | 0.4523 | 0.000 |
| Malic acid | 4.103 a | 4.6 a | 4.59 a | 13.577 b | 5.627 a | 9.011 ab | 10.592 ab | 2.722 a | 8.48 ab | 1.527 | 0.002 |
| Aspartic acid | - | - | - | - | 3.528 a | - | - | 1.588 a | - | 0.988 | 0.000 |
| 4-aminobutyric acid | - | - | - | - | 0.393 a | 1.122 a | - | - | - | 0.193 | 0.000 |
| Threonic acid | - | - | - | 3.512 a | - | 2.879 a | 4.364 a | - | - | 01364 | 0.145 |
| Lauric acid | - | - | - | - | 0.488 a | - | - | 0.548 a | - | 0.001 | 0.000 |
| Citric acid | 46.515 ab | 56.955 ab | - | 42.347 ab | - | - | - | 27.936 a | - | 10.955 | 0.005 |
| Ascorbic acid | 2.745 a | 10.005 ab | 4.495 a | 43.774 bc | - | 8.855 ab | 24.466 b | 82.472 c | 4.954 | 0.000 | |
| Fatty acids | |||||||||||
| Palmitic acid | 7.131 ab | 4.657 a | 9.961 b | 9.068 b | 6.166 ab | 8.678 ab | 8.458 ab | 8.165 ab | 1.811 a | 1.911 | 0.000 |
| Linoleic acid | 4.56 a | 5.195 a | 6.556 a | 6.473 a | 5.993 a | 5.629 a | 9.204 ab | 9.397 ab | 1.203 b | 1.032 | 0.230 |
| a-linolenic acid | - | - | - | - | 2.492 a | - | - | 4.541 a | 0.675 a | 0.025 | 0.050 |
| Estearic acid | 5.652 a | 3.405 a | 5.312 a | 3.846 a | 2.985 a | 5.415 a | 5.226 a | 3.834 a | 0.811 a | 0.862 | 0.000 |
| Phytosterols | 0.000 | ||||||||||
| γ-Tocopherol | 0.531 a | 3.274 b | 2.132 ab | - | 0.316 a | 2.241 ab | 1.238 ab | 0.172 a | 0.082 a | 0.351 | 0.002 |
| (+)-a-Tocopherol | - | 0.609 ab | 12.41 c | 0.799 ab | 2.298 b | 13.129 c | 8.75 bc | 1.576 ab | 0.335 a | 0.309 | 0.000 |
| Campesterol | 3.037 ab | 5.165 ab | 5.971 ab | 0.624 a | 1.264 a | 5.771 ab | 4.196 ab | 0.753 a | 0.195 a | 0.357 | 0.000 |
| Stigmasterol | 1.031 ab | 2.845 b | 1.438 ab | 0.157 a | 0.306 a | 1.68 ab | 1.209 ab | 0.235 a | 0.049 | 0.249 | 0.000 |
| β-Sitosterol | 9.478 ab | 8.739 ab | 17.216 b | 1.758 a | 2.412 a | 16.887 b | 10.763 ab | 1.7 a | 0.296 a | 2.587 | 0.810 |
| Treatments |
SA (mM) | H2O2 (mM) | EC (dS/m) |
|---|---|---|---|
| 1 | 0 | 0 | 3.2 |
| 2 | 0 | 36 | 3.2 |
| 3 | 0.1 | 0 | 3.2 |
| 4 | 0.1 | 36 | 3.2 |
| 5 | 0 | 0 | 3.6 |
| 6 | 0 | 36 | 3.6 |
| 7 | 0.1 | 0 | 3.6 |
| 8 | 0.1 | 36 | 3.6 |
| 9 | 0.01 | 18 | 3.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jimenez-García, S.N.; Garcia-Mier, L.; Ramirez-Gomez, X.S.; Guevara-Gonzalez, R.G.; Aguirre-Becerra, H.; Escobar-Ortiz, A.; Contreras-Medina, L.M.; Garcia-Trejo, J.F.; Vazquez-Cruz, M.A.; Feregrino-Perez, A.A. Characterization of the Key Compounds of Bell Pepper by Spectrophotometry and Gas Chromatography on the Effects of Induced Stress on the Concentration of Secondary Metabolite. Molecules 2023, 28, 3830. https://doi.org/10.3390/molecules28093830
Jimenez-García SN, Garcia-Mier L, Ramirez-Gomez XS, Guevara-Gonzalez RG, Aguirre-Becerra H, Escobar-Ortiz A, Contreras-Medina LM, Garcia-Trejo JF, Vazquez-Cruz MA, Feregrino-Perez AA. Characterization of the Key Compounds of Bell Pepper by Spectrophotometry and Gas Chromatography on the Effects of Induced Stress on the Concentration of Secondary Metabolite. Molecules. 2023; 28(9):3830. https://doi.org/10.3390/molecules28093830
Chicago/Turabian StyleJimenez-García, Sandra N., Lina Garcia-Mier, Xóchitl S. Ramirez-Gomez, Ramon G. Guevara-Gonzalez, Humberto Aguirre-Becerra, Alexandro Escobar-Ortiz, Luis M. Contreras-Medina, Juan F. Garcia-Trejo, Moises A. Vazquez-Cruz, and Ana A. Feregrino-Perez. 2023. "Characterization of the Key Compounds of Bell Pepper by Spectrophotometry and Gas Chromatography on the Effects of Induced Stress on the Concentration of Secondary Metabolite" Molecules 28, no. 9: 3830. https://doi.org/10.3390/molecules28093830