
Rational Design of Disulfide Bridges in BbPETaseCD for Enhancing the Enzymatic Performance in PET Degradation

Abstract
:1. Introduction
2. Results and Discussion
2.1. Rational Design of Disulfide Bridges and Protein Expression
2.2. Activity and Stability of BbPETaseCD and Its Variants
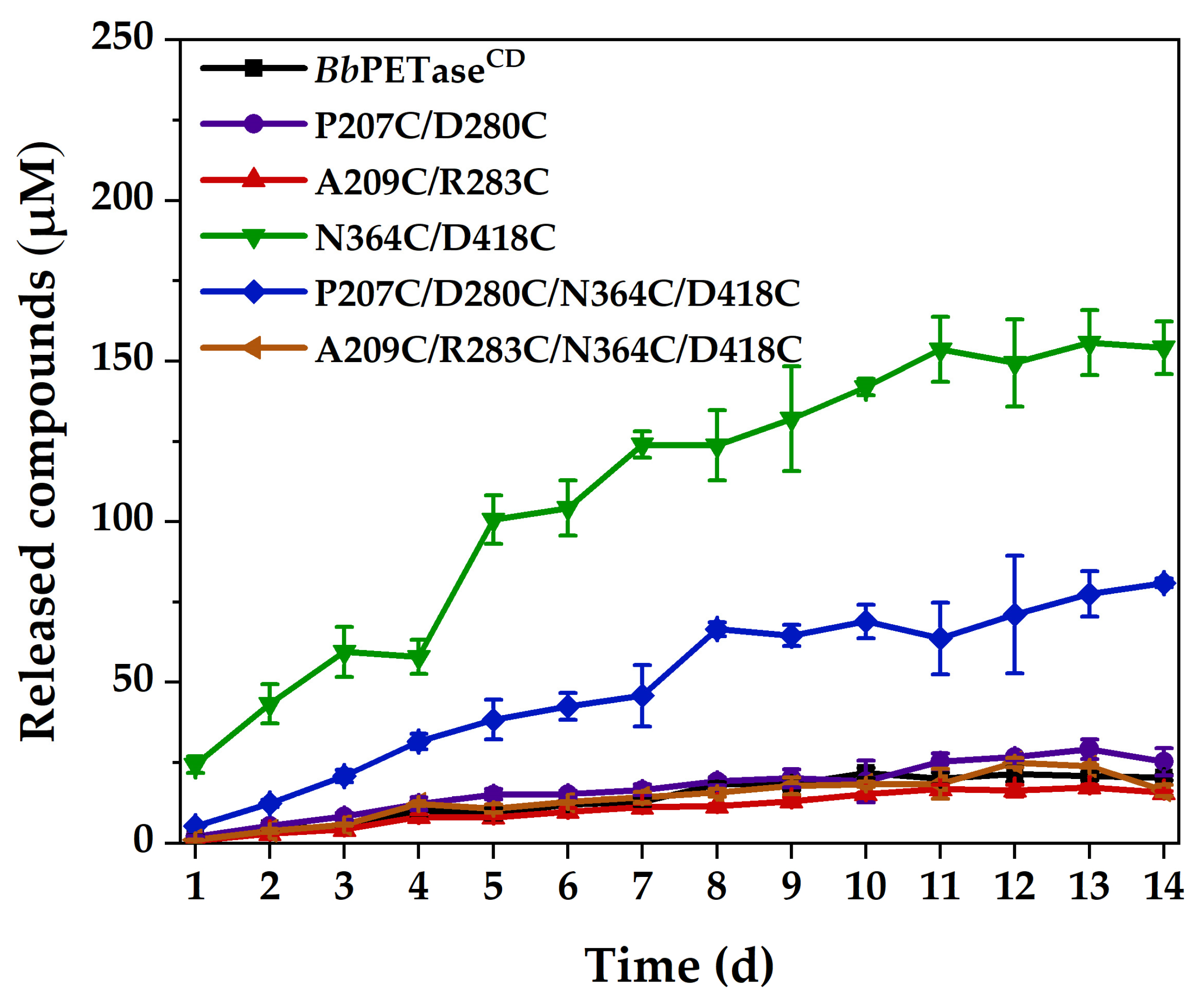
2.3. Activity of BbPETaseCD and Its Variants for PET Film Degradation
2.4. Circular Dichroism (CD) and Fluorescence Analysis
3. Materials and Methods
3.1. Bacterial Strains, Plasmid, and Materials
3.2. Rational Design of Disulfide Bridges
3.3. Site-Directed Mutagenesis
3.4. Expression and Purification
3.5. Analysis of Disulfide Bridge of Enzyme
3.6. Fluorescence and CD Spectrums
3.7. Enzyme Activity Using BHET as Substrate
3.8. Thermal Stability of the Enzyme
3.9. Enzyme Activity for PET Film Degradation
3.10. HPLC Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Suzuki, M.; Date, M.; Kashiwagi, T.; Suzuki, E.; Yokoyama, K. Rational design of a disulfide bridge increases the thermostability of microbial transglutaminase. Appl. Microbiol. Biotechnol. 2022, 106, 4553–4562. [Google Scholar] [CrossRef]
- Li, C.; Ban, X.; Zhang, Y.; Gu, Z.; Hong, Y.; Cheng, L.; Tang, X.; Li, Z. Rational design of disulfide bonds for enhancing the thermostability of the 1,4-α-glucan branching enzyme from Geobacillus thermoglucosidans STB02. J. Agric. Food Chem. 2020, 68, 13791–13797. [Google Scholar] [CrossRef] [PubMed]
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, use, and fate of all plastics ever made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef] [PubMed]
- Suaria, G.; Perold, V.; Lee, J.R.; Lebouard, F.; Aliani, S.; Ryan, P.G. Floating macro- and microplastics around the southern ocean: Results from the antarctic circumnavigation expedition. Environ. Int. 2020, 136, 105494. [Google Scholar] [CrossRef] [PubMed]
- Peeken, I.; Primpke, S.; Beyer, B.; Gütermann, J.; Katlein, C.; Krumpen, T.; Bergmann, M.; Hehemann, L.; Gerdts, G. Arctic sea ice is an important temporal sink and means of transport for microplastic. Nat. Commun. 2018, 9, 1505. [Google Scholar] [CrossRef]
- Singh, A.K.; Bedi, R.; Kaith, B.S. Composite materials based on recycled polyethylene terephthalate and their properties—A comprehensive review. Compos. Part B Eng. 2021, 219, 108928. [Google Scholar] [CrossRef]
- Velásquez, E.J.; Garrido, L.; Guarda, A.; Galotto, M.J.; López De Dicastillo, C. Increasing the incorporation of recycled PET on polymeric blends through the reinforcement with commercial nanoclays. Appl. Clay Sci. 2019, 180, 105185. [Google Scholar] [CrossRef]
- Liu, X.; Wen, Y.; Chen, X.; Tang, T.; Mijowska, E. Co-etching effect to convert waste polyethylene terephthalate into hierarchical porous carbon toward excellent capacitive energy storage. Sci. Total Environ. 2020, 723, 138055. [Google Scholar] [CrossRef]
- Mendiburu-Valor, E.; Mondragon, G.; González, N.; Kortaberria, G.; Martin, L.; Eceiza, A.; Peña-Rodriguez, C. Valorization of urban and marine PET waste by optimized chemical recycling. Resour. Conserv. Recycl. 2022, 184, 106413. [Google Scholar] [CrossRef]
- Fang, P.; Xia, S.; Lu, X. Rapid alcoholysis of PET enhanced by its swelling under high temperature. J. Environ. Chem. Eng. 2022, 10, 107823. [Google Scholar] [CrossRef]
- Shojaei, B.; Abtahi, M.; Najafi, M. Chemical recycling of PET: A stepping-stone toward sustainability. Polym. Adv. Technol. 2020, 31, 2912–2938. [Google Scholar] [CrossRef]
- Wei, R.; von Haugwitz, G.; Pfaff, L.; Mican, J.; Badenhorst, C.P.S.; Liu, W.; Weber, G.; Austin, H.P.; Bednar, D.; Damborsky, J.; et al. Mechanism-based design of efficient PET hydrolases. ACS Catal. 2022, 12, 3382–3396. [Google Scholar] [CrossRef]
- Sulaiman, S.; Yamato, S.; Kanaya, E.; Kim, J.; Koga, Y.; Takano, K.; Kanaya, S. Isolation of a novel cutinase homolog with polyethylene terephthalate-degrading activity from leaf-branch compost by using a metagenomic approach. Appl. Environ. Microbiol. 2012, 78, 1556–1562. [Google Scholar] [CrossRef]
- Kawai, F.; Oda, M.; Tamashiro, T.; Waku, T.; Tanaka, N.; Yamamoto, M.; Mizushima, H.; Miyakawa, T.; Tanokura, M. A novel Ca2+-activated, thermostabilized polyesterase capable of hydrolyzing polyethylene terephthalate from saccharomonospora viridis ahk190. Appl. Microbiol. Biotechnol. 2014, 98, 10053–10064. [Google Scholar] [CrossRef]
- Yoshida, S.; Hiraga, K.; Takehana, T.; Taniguchi, I.; Yamaji, H.; Maeda, Y.; Toyohara, K.; Miyamoto, K.; Kimura, Y.; Oda, K. A bacterium that degrades and assimilates poly(ethylene terephthalate). Science 2016, 351, 1196–1199. [Google Scholar] [CrossRef] [PubMed]
- Sagong, H.; Son, H.F.; Seo, H.; Hong, H.; Lee, D.; Kim, K. Implications for the PET decomposition mechanism through similarity and dissimilarity between PETases from Rhizobacter gummiphilus and Ideonella sakaiensis. J. Hazard. Mater. 2021, 416, 126075. [Google Scholar] [CrossRef] [PubMed]
- Sagong, H.; Kim, S.; Lee, D.; Hong, H.; Lee, S.H.; Seo, H.; Kim, K. Structural and functional characterization of an auxiliary domain-containing PET hydrolase from Burkholderiales bacterium. J. Hazard. Mater. 2022, 429, 128267. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, J.; Yu, X.; Hu, X.; Zhang, H. Engineering Leuconostoc mesenteroides dextransucrase by inserting disulfide bridges for enhanced thermotolerance. Enzym. Microb. Technol. 2020, 139, 109603. [Google Scholar] [CrossRef]
- Gihaz, S.; Bash, Y.; Rush, I.; Shahar, A.; Pazy, Y.; Fishman, A. Bridges to stability: Engineering disulfide bonds towards enhanced lipase biodiesel synthesis. Chemcatchem 2019, 12, 181–192. [Google Scholar] [CrossRef]
- Bashirova, A.; Pramanik, S.; Volkov, P.; Rozhkova, A.; Nemashkalov, V.; Zorov, I.; Gusakov, A.; Sinitsyn, A.; Schwaneberg, U.; Davari, M. Disulfide bond engineering of an endoglucanase from penicillium verruculosum to improve its thermostability. Int. J. Mol. Sci. 2019, 20, 1602. [Google Scholar] [CrossRef]
- Douglas, B.; Alan, D. Disulfide by design 2.0: A web-based tool for disulfide engineering in proteins. BMC Bioinform. 2013, 14, 346. [Google Scholar] [CrossRef]
- Sowdhamini, R.; Srinivasan, N.; Shoichet, B.; Santi, D.V.; Ramakrishnan, C.; Balaram, P. Stereochemical modeling of disulfide bridges. Criteria for introduction into proteins by site-directed mutagenesis. Protein Eng. 1989, 3, 95–103. [Google Scholar] [CrossRef]
- Vinayagam, A. Dsdbase: A consortium of native and modelled disulphide bonds in proteins. Nucleic Acids Res. 2004, 32, 200D–202D. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. Swiss-model: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Bayar, E.; Ren, Y.; Chen, Y.; Hu, Y.; Zhang, S.; Yu, X.; Fan, J. Construction, investigation and application of TEV protease variants with improved oxidative stability. J. Microbiol. Biotechnol. 2021, 31, 1732–1740. [Google Scholar] [CrossRef]
- Li, L.; Zhang, S.; Wu, W.; Guan, W.; Deng, Z.; Qiao, H. Enhancing thermostability of Yarrowia lipolytica lipase 2 through engineering multiple disulfide bonds and mitigating reduced lipase production associated with disulfide bonds. Enzym. Microb. Technol. 2019, 126, 41–49. [Google Scholar] [CrossRef]
- Yang, M.; Yang, S.; Liu, Z.; Li, N.; Li, L.; Mou, H. Rational design of alginate lyase from Microbulbifer sp. Q7 to improve thermal stability. Mar. Drugs. 2019, 17, 378. [Google Scholar] [CrossRef] [PubMed]
- Miles, A.J.; Ramalli, S.G.; Wallace, B.A. Dichroweb, a website for calculating protein secondary structure from circular dichroism spectroscopic data. Protein Sci. 2022, 31, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Sreerama, N.; Woody, R.W. Estimation of protein secondary structure from circular dichroism spectra: Comparison of contin, selcon, and cdsstr methods with an expanded reference set. Anal. Biochem. 2000, 287, 252–260. [Google Scholar] [CrossRef]
- Furukawa, M.; Kawakami, N.; Oda, K.; Miyamoto, K. Acceleration of enzymatic degradation of poly(ethylene terephthalate) by surface coating with anionic surfactants. Chemsuschem 2018, 11, 4018–4025. [Google Scholar] [CrossRef]
- Liu, Z.; Liang, Q.; Wang, P.; Kong, Q.; Fu, X.; Mou, H. Improving the kinetic stability of a hyperthermostable β-mannanase by a rationally combined strategy. Int. J. Biol. Macromol. 2021, 167, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhao, Y.; Mao, S.; Zhu, J.; Zhan, Y.; Cai, D.; Ma, X.; Wang, D.; Chen, S. Enhancing the activity of disulfide-bond-containing proteins via promoting disulfide bond formation in bacillus licheniformis. Int. J. Biol. Macromol. 2023, 233, 123468. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Sulfhydryl Content under Nonreducing Condition (μM) | Sulfhydryl Content under Reducing Condition (μM) | The Proportion of Disulfide Bridge (%) |
|---|---|---|---|
| BbPETaseCD | 1.2 | 20.1 | 93.8 |
| P207C/D280C | 4.7 | 26.5 | 82.2 |
| A209C/R283C | 6.2 | 26.2 | 76.3 |
| N364C/D418C | 3.3 | 30.6 | 89.3 |
| P207C/D280C/N364C/D418C | 6.1 | 37.7 | 83.8 |
| A209C/R283C/N364C/D418C | 5.5 | 39.4 | 86.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, D.; Zhang, L.; Sun, Y. Rational Design of Disulfide Bridges in BbPETaseCD for Enhancing the Enzymatic Performance in PET Degradation. Molecules 2023, 28, 3528. https://doi.org/10.3390/molecules28083528
Huang D, Zhang L, Sun Y. Rational Design of Disulfide Bridges in BbPETaseCD for Enhancing the Enzymatic Performance in PET Degradation. Molecules. 2023; 28(8):3528. https://doi.org/10.3390/molecules28083528
Chicago/Turabian StyleHuang, Dongjian, Lin Zhang, and Yan Sun. 2023. "Rational Design of Disulfide Bridges in BbPETaseCD for Enhancing the Enzymatic Performance in PET Degradation" Molecules 28, no. 8: 3528. https://doi.org/10.3390/molecules28083528