

Effects of Temperature, Metal Ions and Biosurfactants on Interaction Mechanism between Caffeic Acid Phenethyl Ester and Hemoglobin

Abstract
:
1. Introduction
2. Results and Discussion
2.1. Effect of Temperature on the Binding Interaction of CAPE–Hb
2.1.1. Quenching Mechanism and Thermodynamic Analysis
2.1.2. Time-Resolved Fluorescence Spectrometry
2.1.3. UV−Vis Spectrum Analysis
2.1.4. Secondary Structure Analysis
2.1.5. Particle Size Analysis
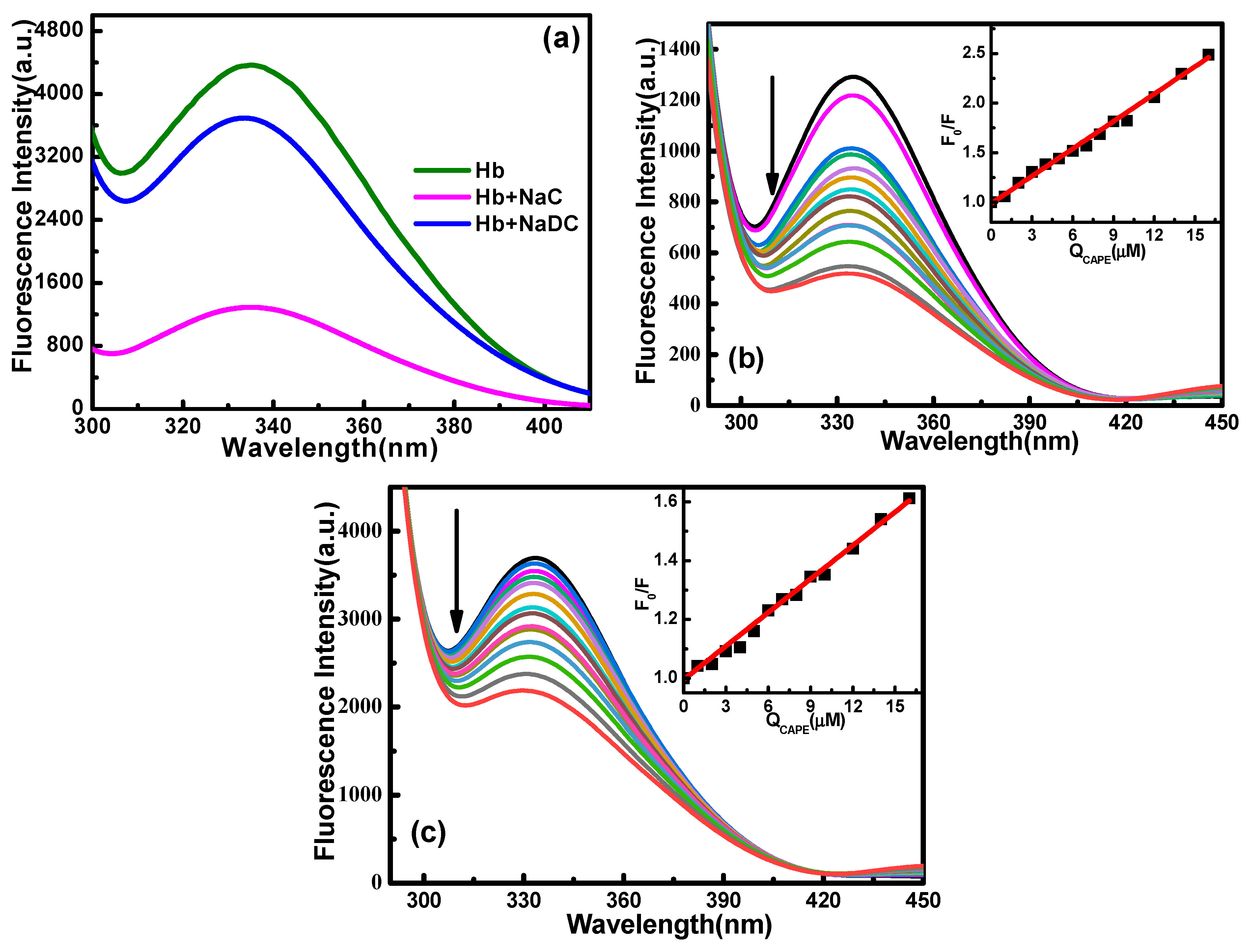
2.2. Effect of Biosurfactants (NaC, NaDC) on the Binding Interaction of CAPE–Hb
2.3. Effect of Metal Ions on Binding Interaction of CAPE–Hb
2.4. Molecular Docking
3. Materials and Methods
3.1. Materials
3.2. Preparation of Sample
3.3. Fluorescence Quenching Spectroscopy
3.4. Time-Resolved Fluorescence Spectrometry
3.5. UV−Vis Adsorption Spectroscopy (UV−Vis)
3.6. Circular Dichroism (CD)
3.7. Dynamic Light Scattering (DLS)
3.8. Molecular Docking
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Tolba, M.F.; Azab, S.S.; Khalifa, A.E.; Abdel-Rahman, S.Z.; Abdel-Naim, A.B. Caffeic acid phenethyl ester, a promising component of propolis with a plethora of biological activities: A review on its anti-inflammatory, neuroprotective, hepatoprotective, and cardioprotective effects. IUBMB Life 2013, 65, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.X.; Tang, Y.P.; Li, N.G.; Zhu, Y.; Duan, J.A. Bioactivity and chemical synthesis of caffeic acid phenethyl ester and its derivatives. Molecules 2014, 19, 16458–16476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rzepecka-Stojko, A.; Kabała-Dzik, A.; Moździerz, A.; Kubina, R.; Wojtyczka, R.D.; Stojko, R.; Dziedzic, A.; Jastrzębska-Stojko, Ż.; Jurzak, M.; Buszman, E.; et al. Caffeic acid phenethyl ester and ethanol extract of propolis induce the complementary cytotoxic effect on triple-negative breast cancer cell lines. Molecules 2015, 20, 9242–9262. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.P.; Lin, C.Y.; Liu, C.C.; Su, L.C.; Huo, C.H.; Kuo, Y.Y.; Tseng, J.C.; Hsu, J.M.; Chen, C.K.; Chuu, C.P. Caffeic acid phenethyl ester as a potential treatment for advanced prostate cancer targeting akt signaling. Int. J. Mol. Sci. 2013, 14, 5264–5283. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.; Castellano, A.; Uzoaru, C.; Lim, J.; Carvis, N.; Wieczorek, P.; Barsotti, R.; Young, L.H.; Chen, Q. The effects of Caffeic Acid Phenethyl Ester (CAPE) on oxidative stress and hypoxia-induced cell damage. FASEB J. 2017, 31, 1070.9. [Google Scholar]
- Hassan, N.A.; El-Bassossy, H.M.; Mahmoud, M.F.; Fahmy, A. Caffeic acid phenethyl ester, a 5-lipoxygenase enzyme inhibitor, alleviates diabetic atherosclerotic manifestations: Effect on vascular reactivity and stiffness. Chem. Biol. Interact. 2014, 213, 28–36. [Google Scholar] [CrossRef]
- Kuo, Y.Y.; Jim, W.T.; Su, L.C.; Chung, C.J.; Lin, C.Y.; Huo, C.; Tseng, J.C.; Huang, S.H.; Lai, C.J.; Chen, B.C.; et al. Caffeic Acid phenethyl ester is a potential therapeutic agent for oral cancer. Int. J. Mol. Sci. 2015, 12, 10748–10766. [Google Scholar] [CrossRef] [Green Version]
- Paeng, S.H.; Jung, W.K.; Park, W.S.; Lee, D.S.; Kim, G.Y.; Choi, Y.H.; Seo, S.K.; Jang, W.H.; Choi, J.S.; Lee, Y.M. Caffeic acid phenethyl ester reduces the secretion of vascular endothelial growth factor through the inhibition of the ROS, PI3K and HIF-1α signaling pathways in human retinal pigment epithelial cells under hypoxic conditions. Int. J. Mol. Med. 2015, 35, 1419–1426. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Kaur, D.; Bansal, N. Caffeic Acid Phenethyl Ester (CAPE) Prevents Development of STZ-ICV Induced dementia in Rats. Pharmacogn. Mag. 2017, 13, S10–S15. [Google Scholar]
- Armutcu, F.; Akyol, S.; Ustunsoy, S.; Turan, F.F. Therapeutic potential of caffeic acid phenethyl ester and its anti-inflammatory and immunomodulatory effects (Review). Exp. Ther. Med. 2015, 9, 1582–1588. [Google Scholar] [CrossRef] [Green Version]
- Nai, X.; Chen, Y.R.; Hao, S.Y.; Liu, M.; Zhang, Q.; Liu, J.; Li, M.Y.; Kong, J. Temperature, pH and additives effects on the binding of Caffeic acid phenethyl ester to the native state of bovine serum albumin. J. Chem. Thermodyn. 2022, 168, 106724. [Google Scholar] [CrossRef]
- Nai, X.; Chen, Y.R.; Zhang, Q.; Hao, S.; Xuan, H.Z.; Liu, J. Interaction between caffeic acid phenethyl ester and protease: Monitoring by spectroscopic and molecular docking approaches. Luminescence 2022, 37, 1025–1036. [Google Scholar] [CrossRef] [PubMed]
- Bi, S.Y.; Sun, Y.T.; Qiao, C.Y.; Zhang, H.Q.; Liu, C.M. Binding of several anti-tumor drugs to bovine serum albumin: Fluorescence study. J. Lumin. 2009, 129, 541–547. [Google Scholar] [CrossRef]
- Gull, N.; Kumar, S.; Ahmad, B.; Khan, R.H.; Din, K. Influence of urea additives on micellar morphology/protein conformation. Colloids Surf. B Biointerfaces 2006, 51, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, B.; Kamal, M.Z.; Khan, R.H. Alkali-Induced Conformational transition in different domains of bovine serum albumin. Protein Pept. Lett. 2004, 11, 307–315. [Google Scholar] [CrossRef]
- Khan, R.H.; Rasheedi, S.; Haq, S.K. Effect of pH, temperature and alcohols on the stability of glycosylated and deglycosylated stem bromelain. J. Biosci. 2003, 28, 709–714. [Google Scholar] [CrossRef]
- Patel, R.; Maurya, N.; Parray, M.U.D.; Farooq, N.; Siddique, A.; Verma, K.L.; Dohare, N. Esterase activity and conformational changes of bovine serum albumin toward interaction with mephedrone: Spectroscopic and computational studies. J. Mol. Recognit. 2018, 31, e2734. [Google Scholar] [CrossRef]
- Bhat, I.A.; Roy, B.; Hazra, P.; Kabir-ud-Din. Conformational and solution dynamics of hemoglobin (Hb) in presence of a cleavable gemini surfactant: Insights from spectroscopy, atomic force microscopy, molecular docking and density functional theory. J. Colloid. Interf. Sci. 2019, 538, 489–498. [Google Scholar] [CrossRef]
- Wu, Q.; Zhao, H.; Chen, X.; Cai, Z.W. Interaction of bisphenol A 3, 4-quinone metabolite with human hemoglobin, human serum albumin and cytochrome in vitro. Chemosphere. 2019, 220, 930–936. [Google Scholar] [CrossRef]
- Xu, L.; Liu, Z.; Liao, T.; Tuo, X. Probing the interaction between levamlodipine and hemoglobin based on spectroscopic and molecular docking methods. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2019, 223, 117306. [Google Scholar] [CrossRef]
- Dong, J.W.; Li, X.L.; Zhou, Y.Q.; Lu, Y.H.; Lv, Y.P.; Chi, Y.L.; He, Q. Interactions of Gallic Acid with Porcine Hemoglobin: Effect on the Redox State and Structure of Hemoglobin. J. Agric. Food Chem. 2021, 69, 397–403. [Google Scholar] [CrossRef]
- Malarkani, K.; Sarkar, I.; Selvam, S. Denaturation studies on bovine serum albumin–bile salt system: Bile salt stabilizes bovine serum albumin through hydrophobicity. J. Pharm. Anal. 2018, 8, 27–36. [Google Scholar] [CrossRef]
- Eftink, M.; Ghiron, C. Fluorescence quenching of indole and model micelle systems. J. Phys. Chem. C 1976, 80, 486–493. [Google Scholar] [CrossRef]
- Trnková, L.; Bousova, I.; Kubicek, V. Binding of naturally occurring hydroxycinnamic acids to bovine serum albumin. Nat. Sci. 2010, 2, 563–570. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wu, F.; Tan, J.; Wang, K.; Zhang, C.P.; Zheng, H.Q.; Hu, F.L. Caffeic acid phenethyl ester exhibiting distinctive binding interaction with human serum albumin implies the pharmacokinetic basis of propolis bioactive components. J. Pharmaceut. Biomed. 2016, 122, 21–28. [Google Scholar] [CrossRef]
- Lehrer, S.S. Solute perturbation of protein fluorescence. Quenching of the tryptophyl fluorescence of model compounds and of lysozyme by iodide ion. Biochemistry 1971, 10, 3254–3263. [Google Scholar] [CrossRef]
- Roy, A.S.; Dinda, A.K.; Chaudhury, S. Binding of antioxidant flavonol morin to the native state of bovine serum albumin: Effects of urea and metal ions on the binding. J. Lumin. 2014, 145, 741–751. [Google Scholar]
- Ross, P.D.; Subramanian, S. Thermodynamics of protein association reactions: Forces contributing to stability. Biochemistry 1981, 20, 3096–3102. [Google Scholar] [CrossRef]
- Tiwari, A.; Bhatia, P.; Randhawa, J.K. Systematic spectroscopic investigation of structural changes and corona formation of bovine serum albumin over magneto-fluorescent nanoparticles. Dalton Trans. 2020, 49, 12380–12389. [Google Scholar] [CrossRef]
- Das, S.; Santra, S.; Rohman, M.A.; Ray, M.; Jana, M.; Roy, A.S. An insight into the binding of 6-hydroxyflavone with hen egg white lysozyme: A combined approach of multi-spectroscopic and computational studies. J. Biomol. Struct. Dyn. 2018, 37, 4019–4034. [Google Scholar] [CrossRef]
- Dohare, N.; Siddiquee, M.A.; Parray, M.D.; Kumar, A.; Patel, R. Esterase activity and interaction of human hemoglobin with diclofenac sodium: A spectroscopic and molecular docking study. J. Mol. Recognit. 2020, 33, e2841. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yang, G.; Mei, Z. Spectroscopic and dynamic light scattering studies of the interaction between pterodontic acid and bovine serum albumin. Acta Pharm. Sin. B. 2012, 2, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Nobbmann, U.; Connah, M.; Fish, B.; Varley, P.; Gee, C.; Mulot, S.; Chen, J.; Zhou, L.; Lu, Y.L.; Shen, F.; et al. Dynamic light scattering as a relative tool for assessing the molecular integrity and stability of monoclonal antibodies. Biotechnol. Genet. Eng. 2007, 24, 117–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maity, S.; Acharjee, A.; Saha, B. Chapter 15—Application of biosurfactant as biocontrol agents against soil-borne and root-borne plant pathogens. In Applications of Biosurfactant in Agriculture; Academic Press: Cambridge, MA, USA, 2022; pp. 283–302. [Google Scholar]
- Pal, A.; Mondal, M.H.; Adhikari, A.; Bhattarai, A.; Saha, B. Scientific information about sugar-based emulsifiers: A comprehensive review. RSC Adv. 2021, 11, 33004–33016. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.S.; Dinda, A.K.; Pandey, N.K.; Dasgupta, S. Effects of urea, metal ions and surfactants on the binding of baicalein with bovine serum albumin. J. Pharm. Anal. 2016, 6, 256–267. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Huang, L.; Li, R.J.; Zhang, Z.W.; Chen, J.; Tang, H.J. Multispectroscopic and computational evaluation of the binding of flavonoids with bovine serum albumin in the presence of Cu2+. Food Chem. 2022, 385, 132656. [Google Scholar] [CrossRef]
- Li, D.J.; Zhu, M.; Xu, C.; Ji, B.M. Characterization of the baicaleinebovine serum albumin complex without or with Cu2+ or Fe3+ by spectroscopic approaches. Eur. J. Med. Chem. 2011, 46, 588–599. [Google Scholar] [CrossRef]
- Yu, X.Y.; Liu, R.H.; Yang, F.X.; Ji, D.H.; Li, X.F.; Chen, J.; Huang, H.W.; Yi, P.G. Study on the interaction between dihydromyricetin and bovine serum albumin by spectroscopic techniques. J. Mol. Struct. 2011, 985, 407–412. [Google Scholar] [CrossRef]
- Liu, B.S.; Yan, X.N.; Cao, S.N.; Chong, B.H.; Yang, C.; Lü, Y.K. Interaction of Salicylic Acid with Bovine Hemoglobin and Effect of the Coexistent Metal Ion on the Reaction. Spectrosc. Lett. 2013, 46, 165–174. [Google Scholar] [CrossRef]
- Liu, B.S.; Yang, C.; Yan, X.N.; Wang, J.; Lv, Y.K. Study on the conjugation mechanism of colistin sulfate with bovine serum albumin and effect of the metal ions on the reaction. J. Lumin. 2012, 132, 1133–1138. [Google Scholar] [CrossRef]
- Gu, J.; Zheng, S.; Zhao, H.; Sun, T. Investigation on the interaction between triclosan and bovine serum albumin by spectroscopic methods. J. Environ. Sci. Health B 2020, 55, 52–59. [Google Scholar] [CrossRef]
- Li, S.; Huang, K.; Zhong, M.; Guo, J.; Wang, W.Z.; Zhu, R.H. Comparative studies on the interaction of caffeic acid, chlorogenic acid and ferulic acid with bovine serum albumin. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2010, 77, 680–686. [Google Scholar] [CrossRef]
- Liu, T.; Liu, M.; Guo, Q.; Liu, Y.L.; Zhao, Y.N.; Wu, Y.S.; Sun, B.; Wang, Q.P.; Liu, J.; Han, J. Investigation of binary and ternary systems of human serum albumin with oxyresveratrol/piceatannol and/or mitoxantrone by multipectroscopy, molecular docking and cytotoxicity evaluation. J. Mol. Liq. 2020, 311, 113364. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T | Ksv (104, M−1) | Kq (1012, M−1·s−1) | Ka (103, M−1) | n | ∆Gm (kJ/mol) | ∆Hm (kJ/mol) | ∆Sm [J/(mol·K)] |
|---|---|---|---|---|---|---|---|
| 298.2 | 6.41 ± 0.008 | 6.41 ± 0.008 | 5.89 ± 0.013 | 0.78 ± 0.009 | −21.5 ± 0.054 | −40.8 ± 0.05 | −65.1 ± 0.02 |
| 302.2 | 6.00 ± 0.002 | 6.00 ± 0.002 | 4.27 ± 0.009 | 0.76 ± 0.008 | −21.0 ± 0.052 | ||
| 306.2 | 5.81 ± 0.005 | 5.81 ± 0.005 | 3.52 ± 0.008 | 0.75 ± 0.014 | −20.8 ± 0.057 | ||
| 310.2 | 5.45 ± 0.003 | 5.45 ± 0.003 | 3.13 ± 0.007 | 0.74 ± 0.019 | −20.7 ± 0.050 |
| Molar Ratio of Hb to CAPE | (ns) | (ns) | (ns) | ||
|---|---|---|---|---|---|
| 1:0 | 0.98 | 0.89 | 8.74 | 0.11 | 1.83 |
| 1:1 | 0.85 | 0.84 | 6.91 | 0.15 | 1.81 |
| 1:2 | 0.86 | 0.83 | 6.64 | 0.17 | 1.84 |
| 1:3 | 0.85 | 0.83 | 6.52 | 0.17 | 1.82 |
| SAA | C (mM) | Ksv (104, M−1) | Kq (1012, M−1·s−1) | Ka (104, M−1) | n |
|---|---|---|---|---|---|
| NaC | 1.0 | 9.17 ± 0.014 | 9.17 ± 0.014 | 0.84 ± 0.018 | 0.78 ± 0.039 |
| 3.3 | 7.68 ± 0.001 | 7.68 ± 0.001 | 4.47 ± 0.027 | 1.09 ± 0.053 | |
| 10.0 | 6.60 ± 0.018 | 6.60 ± 0.018 | 1.32 ± 0.015 | 0.84 ± 0.036 | |
| 20.0 | 4.51 ± 0.009 | 4.51 ± 0.009 | 0.45 ± 0.003 | 0.78 ± 0.036 | |
| NaDC | 1.0 | 8.92 ± 0.016 | 8.92 ± 0.016 | 9.72 ± 0.015 | 0.90 ± 0.034 |
| 2.5 | 7.26 ± 0.001 | 7.26 ± 0.001 | 15.84 ± 0.016 | 0.94 ± 0.031 | |
| 8.0 | 5.93 ± 0.005 | 5.93 ± 0.005 | 36.47 ± 0.065 | 1.19 ± 0.036 | |
| 20.0 | 5.39 ± 0.008 | 5.39 ± 0.008 | 10.6 ± 0.025 | 1.06 ± 0.039 |
| SAA | T (K) | Ksv (104, M−1) | Ka (104, M−1) | n | ∆Gm (kJ/mol) | ∆Hm (kJ/mol) | ∆Sm [J/(mol·K)] |
|---|---|---|---|---|---|---|---|
| NaC | 298.2 | 7.68 ± 0.001 | 4.47 ± 0.027 | 1.09 ± 0.053 | −26.5 ± 0.008 | −90.1 ± 0.499 | −213 ± 0.165 |
| 302.2 | 7.52 ± 0.001 | 3.89 ± 0.023 | 1.03 ± 0.044 | −26.5 ± 0.008 | |||
| 306.2 | 7.45 ± 0.001 | 1.62 ± 0.021 | 1.08 ± 0.051 | −24.7 ± 0.014 | |||
| NaDC | 298.2 | 7.26 ± 0.001 | 15.84 ± 0.016 | 0.94 ± 0.031 | −29.7 ± 0.004 | −73.5 ± 0.244 | −148 ± 0.809 |
| 302.2 | 7.17 ± 0.002 | 8.51 ± 0.013 | 0.97 ± 0.025 | −28.5 ± 0.006 | |||
| 306.2 | 6.59 ± 0.002 | 7.08 ± 0.091 | 0.86 ± 0.017 | −28.4 ± 0.007 |
| Metal Ions | Ka (M−1) | R2 |
|---|---|---|
| without | 5.89 × 103 | 0.95 |
| Ca2+ | 9.42 × 100 | 0.99 |
| Cu2+ | 5.88 × 104 | 0.98 |
| K+ | 1.46 × 103 | 0.97 |
| Mn2+ | 2.14 × 103 | 0.98 |
| Ni2+ | 7.24 × 103 | 0.98 |
| Chemical | Source | CAS Number | Storage Method | Mass Fraction Purity a |
|---|---|---|---|---|
| CAPE | Ark Pharm, Inc., Shanghai, China | 104594-70-9 | Keep in dark place, Sealed in dry, Store in freezer, under 20 °C | ≥0.980 |
| Hb | Aladdin Biochemical Technology Co., Ltd., Shanghai, China | 9008-02-0 | Store in the freezer at 2 to 8 °C | ≥0.980 |
| NaC | InnoChem Science & Technology Co., Ltd., Beijing, China | 361-09-1 | Stored in desiccator | ≥0.990 |
| NaDC | 302-95-4 | ≥0.990 | ||
| Na2HPO4 | Damao Chemical Reagent Co., Ltd., Tianjin, China | 7558-79-4 | ≥0.990 | |
| NaH2PO4 | 7558-80-7 | ≥0.990 | ||
| CaCl2 | 10043-52-4 | ≥0.996 | ||
| KCl | 7447-40-7 | ≥0.990 | ||
| CuCl2·2H2O | 1344-67-8 | ≥0.990 | ||
| MnCl2·4H2O | 13446-34-9 | ≥0.990 | ||
| NiCl2·6H2O | 7791-20-0 | ≥0.980 | ||
| C2H5OH | 64-17-5 | Stored in dark place, sealed in dry | ≥0.997 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhao, Z.; Nai, X.; Li, M.; Kong, J.; Chen, Y.; Liu, M.; Zhang, Q.; Liu, J.; Yan, H. Effects of Temperature, Metal Ions and Biosurfactants on Interaction Mechanism between Caffeic Acid Phenethyl Ester and Hemoglobin. Molecules 2023, 28, 3440. https://doi.org/10.3390/molecules28083440
Li Y, Zhao Z, Nai X, Li M, Kong J, Chen Y, Liu M, Zhang Q, Liu J, Yan H. Effects of Temperature, Metal Ions and Biosurfactants on Interaction Mechanism between Caffeic Acid Phenethyl Ester and Hemoglobin. Molecules. 2023; 28(8):3440. https://doi.org/10.3390/molecules28083440
Chicago/Turabian StyleLi, Yutong, Zhen Zhao, Xiao Nai, Mingyuan Li, Jing Kong, Yanrong Chen, Min Liu, Qian Zhang, Jie Liu, and Hui Yan. 2023. "Effects of Temperature, Metal Ions and Biosurfactants on Interaction Mechanism between Caffeic Acid Phenethyl Ester and Hemoglobin" Molecules 28, no. 8: 3440. https://doi.org/10.3390/molecules28083440