
Chitosan and Chitosan Nanoparticles: Parameters Enhancing Antifungal Activity

Abstract
:1. Introduction
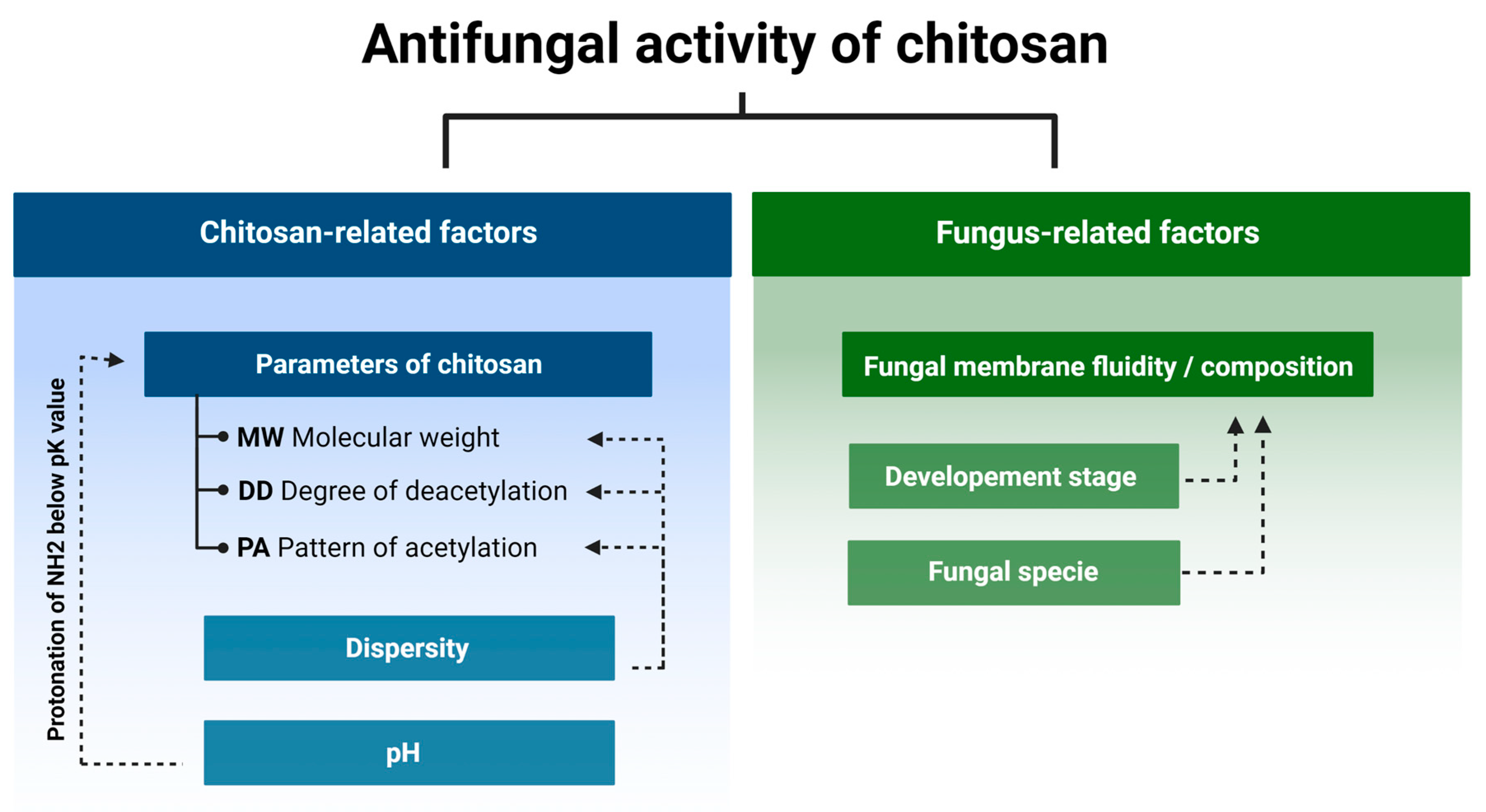
2. Determinants of Chitosan Antifungal Activity
2.1. Fungi Cell Membrane Composition and Chitosan Susceptibility
2.2. Fungus Developmental Stages and Chitosan Susceptibility
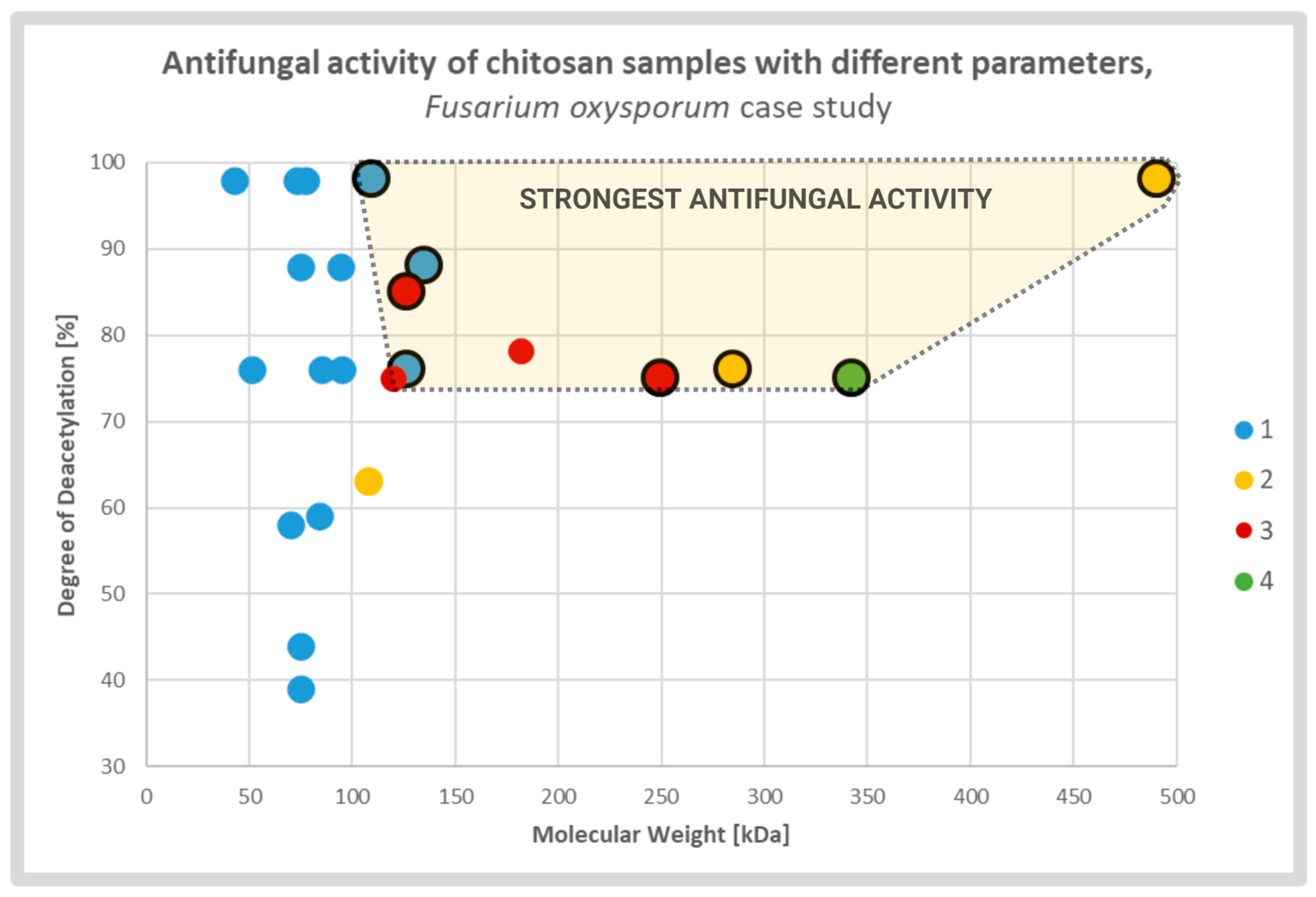
2.3. Physiochemical Attributes of Chitosan and Its Antifungal Activity
2.4. Chitosan Degree of Deacetylation and Antifungal Activity
2.5. Chitosan Pattern of Acetylation and Antifungal Activity
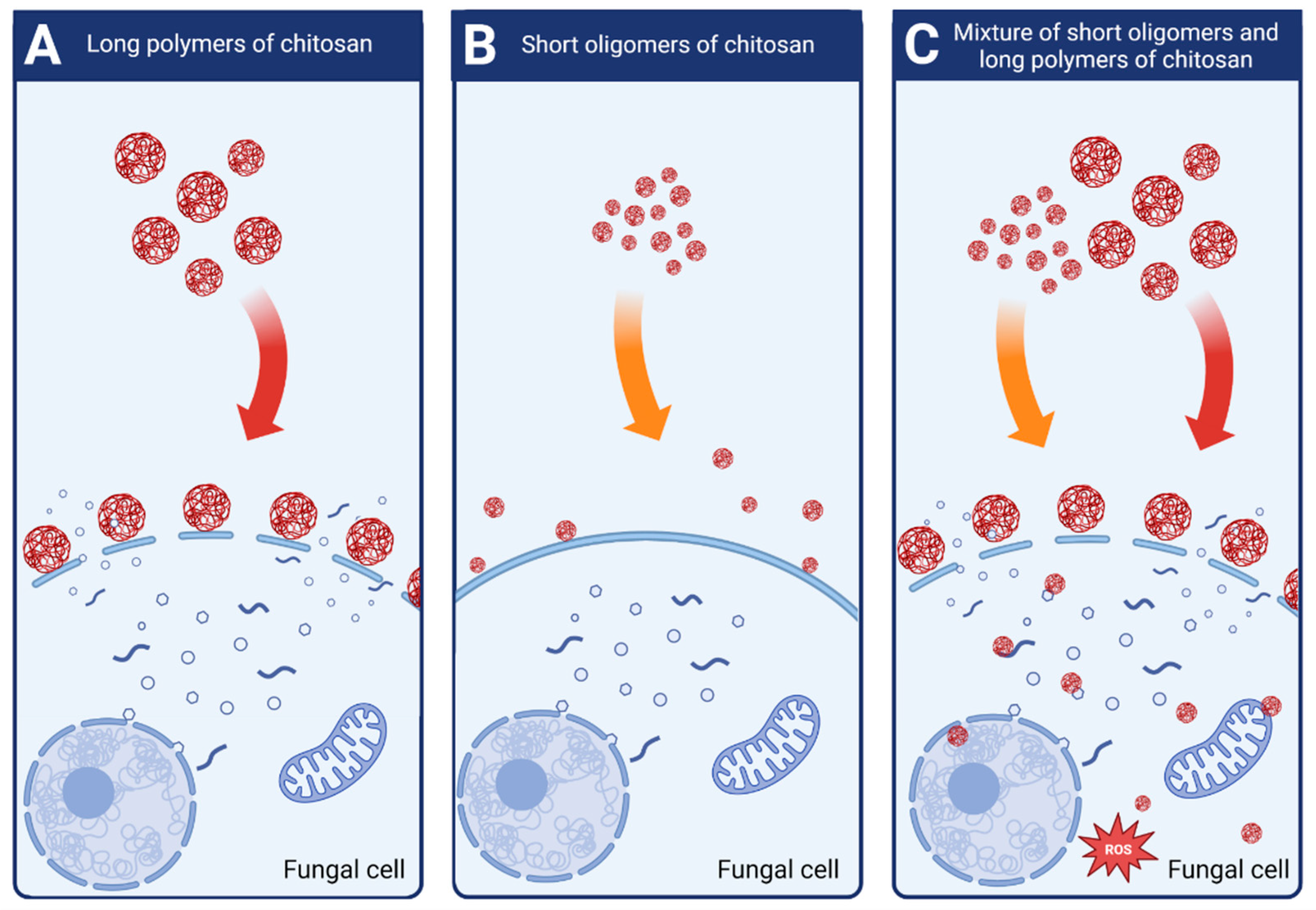
2.6. Dispersity of the Chitosan Sample Promotes Antifungal Activity
2.7. Effect of pH on Chitosan Antifungal Activity
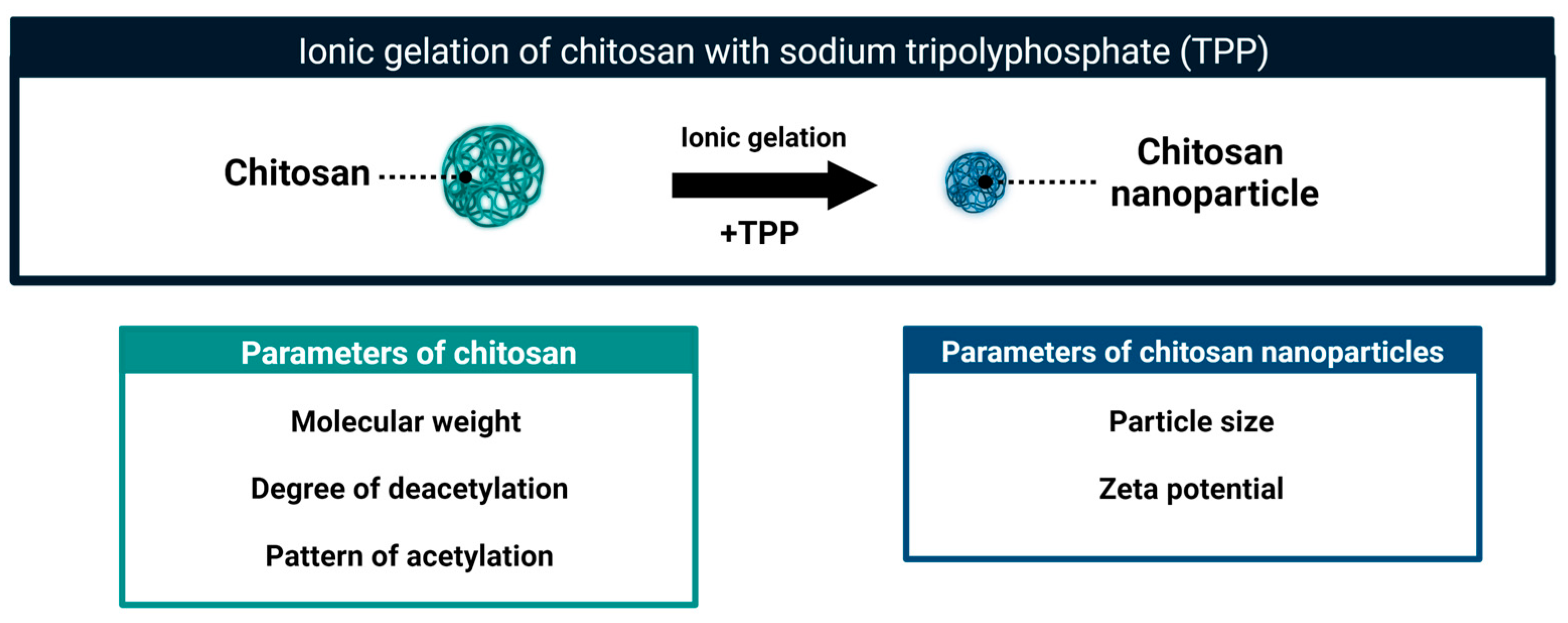
3. Chitosan Nanoparticles: The Way to Increase Antifungal Activity
Particle Size and Zeta Potential Define Antifungal Activity of Chitosan Nanoparticles
4. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Terkula Iber, B.; Azman Kasan, N.; Torsabo, D.; Wese Omuwa, J. A Review of Various Sources of Chitin and Chitosan in Nature. J. Renew. Mater. 2022, 10, 1097–1123. [Google Scholar] [CrossRef]
- Kean, T.; Thanou, M. Biodegradation, biodistribution and toxicity of chitosan. Adv. Drug Deliv. Rev. 2010, 62, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.R.; Muzzarelli, R.; Muzzarelli, C.; Sashiwa, H.; Domb, A.J. Chitosan Chemistry and Pharmaceutical Perspectives. Chem. Rev. 2004, 104, 6017–6084. [Google Scholar] [CrossRef] [PubMed]
- Kheiri, A.; Jorf, S.A.M.; Malihipour, A.; Saremi, H.; Nikkhah, M. Application of chitosan and chitosan nanoparticles for the control of Fusarium head blight of wheat (Fusarium graminearum) in vitro and greenhouse. Int. J. Biol. Macromol. 2016, 93, 1261–1272. [Google Scholar] [CrossRef]
- Long, L.T.; Tan, L.V.; Boi, V.N.; Trung, T.S. Antifungal activity of water-soluble chitosan against Colletotrichum toloni in postharvest chili pepper. J. Food Process. Preserv. 2018, 42, e13339. [Google Scholar] [CrossRef]
- Palma-Guerrero, J.; Jansson, H.B.; Salinas, J.; Lopez-Llorca, L.V. Effect of chitosan on hyphal growth and spore germination of plant pathogenic and biocontrol fungi. J. Appl. Microbiol. 2008, 104, 541–553. [Google Scholar] [CrossRef]
- Zavala-González, E.A.; Lopez-Moya, F.; Aranda-Martinez, A.; Cruz-Valerio, M.; Lopez-Llorca, L.V.; Ramírez-Lepe, M. Tolerance to chitosan by Trichoderma species is associated with low membrane fluidity. J. Basic Microbiol. 2016, 56, 792–800. [Google Scholar] [CrossRef]
- Verlee, A.; Mincke, S.; Stevens, C.V. Recent developments in antibacterial and antifungal chitosan and its derivatives. Carbohydr. Polym. 2017, 164, 268–283. [Google Scholar] [CrossRef]
- Li, K.; Xing, R.; Liu, S.; Li, P. Chitin and chitosan fragments responsible for plant elicitor and growth stimulator. J. Agric. Food Chem. 2020, 68, 12203–12211. [Google Scholar] [CrossRef]
- Brulé, D.; Villano, C.; Davies, L.J.; Trdá, L.; Claverie, J.; Héloir, M.C.; Chiltz, A.; Adrian, M.; Darblade, B.; Tornero, P. The grapevine (Vitis vinifera) LysM receptor kinases Vv LYK 1-1 and Vv LYK 1-2 mediate chitooligosaccharide-triggered immunity. Plant Biotechnol. J. 2019, 17, 812–825. [Google Scholar] [CrossRef] [Green Version]
- Shih, P.-Y.; Liao, Y.-T.; Tseng, Y.-K.; Deng, F.-S.; Lin, C.-H. A potential antifungal effect of chitosan against Candida albicans is mediated via the inhibition of SAGA complex component expression and the subsequent alteration of cell surface integrity. Front. Microbiol. 2019, 10, 602. [Google Scholar] [CrossRef] [Green Version]
- Peña, A.; Sánchez, N.S.; Calahorra, M. Effects of chitosan on Candida albicans: Conditions for its antifungal activity. BioMed. Res. Int. 2013, 2013, 527549. [Google Scholar] [CrossRef] [Green Version]
- Younes, I.; Sellimi, S.; Rinaudo, M.; Jellouli, K.; Nasri, M. Influence of acetylation degree and molecular weight of homogeneous chitosans on antibacterial and antifungal activities. Int. J. Food Microbiol. 2014, 185, 57–63. [Google Scholar] [CrossRef]
- Lei, J.; Yang, L.; Zhan, Y.; Wang, Y.; Ye, T.; Li, Y.; Deng, H.; Li, B. Plasma treated polyethylene terephthalate/polypropylene films assembled with chitosan and various preservatives for antimicrobial food packaging. Colloids Surf. B Biointerfaces 2014, 114, 60–66. [Google Scholar] [CrossRef]
- Meng, W.; Sun, H.; Mu, T.; Garcia-Vaquero, M. Pickering emulsions with chitosan and macroalgal polyphenols stabilized by layer-by-layer electrostatic deposition. Carbohydr. Polym. 2023, 300, 120256. [Google Scholar] [CrossRef]
- Apte, G.; Repanas, A.; Willems, C.; Mujtaba, A.; Schmelzer, C.E.H.; Raichur, A.; Syrowatka, F.; Groth, T. Effect of Different Crosslinking Strategies on Physical Properties and Biocompatibility of Freestanding Multilayer Films Made of Alginate and Chitosan. Macromol. Biosci. 2019, 19, e1900181. [Google Scholar] [CrossRef]
- Garavand, F.; Rouhi, M.; Razavi, S.H.; Cacciotti, I.; Mohammadi, R. Improving the integrity of natural biopolymer films used in food packaging by crosslinking approach: A review. Int. J. Biol. Macromol. 2017, 104 Pt A, 687–707. [Google Scholar] [CrossRef]
- Jones, R.S.; Gardner, R.; Pfeifer, C.L.H.; Morrill, S.D.I.; Ross, P. Chitin and Chitosan Final Registration Review Decision. Docket Number EPA-HQ-OPP-2007-0566; Chitin Case 6063. 2008. Available online: regulations.gov (accessed on 5 March 2023).
- Niaz, T.; Shabbir, S.; Noor, T.; Abbasi, R.; Raza, Z.A.; Imran, M. Polyelectrolyte multicomponent colloidosomes loaded with nisin Z for enhanced antimicrobial activity against foodborne resistant pathogens. Front. Microbiol. 2018, 8, 2700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonin, D.; Pochkaeva, E.; Zhuravskii, S.; Postnov, V.; Korolev, D.; Vasina, L.; Kostina, D.; Mukhametdinova, D.; Zelinskaya, I.; Skorik, Y.; et al. Biological Safety and Biodistribution of Chitosan Nanoparticles. Nanomaterials 2020, 10, 810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondéjar-López, M.; López-Jiménez, A.J.; Martínez, J.C.G.; Ahrazem, O.; Gómez-Gómez, L.; Niza, E. Thymoquinone-Loaded Chitosan Nanoparticles as Natural Preservative Agent in Cosmetic Products. Int. J. Mol. Sci. 2022, 23, 898. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, F.; Li, C.; An, H.; Wan, T.; Zhang, P. Application of chitosan and its derivative polymers in clinical medicine and agriculture. Polymers 2022, 14, 958. [Google Scholar] [CrossRef] [PubMed]
- Abdelfattah, I.; El-Shamy, A. Chitosan as potential de-coloring agent for synthetic and textile industrial wastewater. J. Environ. Account. Manag. 2022, 10, 305–319. [Google Scholar] [CrossRef]
- Hajji, S.; Younes, I.; Rinaudo, M.; Jellouli, K.; Nasri, M. Characterization and in vitro evaluation of cytotoxicity, antimicrobial and antioxidant activities of chitosans extracted from three different marine sources. Appl. Biochem. Biotechnol. 2015, 177, 18–35. [Google Scholar] [CrossRef] [PubMed]
- Hongpattarakere, T.; Riyaphan, O. Effect of deacetylation conditions on antimicrobial activity of chitosans prepared from carapace of black tiger shrimp. Songklanakarin J. Sci. Technol. 2008, 30, 1. [Google Scholar]
- Li, K.; Xing, R.; Liu, S.; Qin, Y.; Meng, X.; Li, P. Microwave-assisted degradation of chitosan for a possible use in inhibiting crop pathogenic fungi. Int. J. Biol. Macromol. 2012, 51, 767–773. [Google Scholar] [CrossRef]
- Tsai, G.-J.; Su, W.-H.; Chen, H.-C.; Pan, C.-L. Antimicrobial activity of shrimp chitin and chitosan from different treatments and applications of fish preservation. Fish. Sci. 2002, 68, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Zakrzewska, A.; Boorsma, A.; Delneri, D.; Brul, S.; Oliver, S.G.; Klis, F.M. Cellular processes and pathways that protect Saccharomyces cerevisiae cells against the plasma membrane-perturbing compound chitosan. Eukaryot. Cell 2007, 6, 600–608. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Lauzardo, A.N.; Bautista-Banos, S.; Velazquez-Del Valle, M.G.; Mendez-Montealvo, M.G.; Sanchez-Rivera, M.M.; Bello-Perez, L.A. Antifungal effects of chitosan with different molecular weights on in vitro development of Rhizopus tolonifera (Ehrenb.:Fr.) Vuill. Carbohydr. Polym. 2008, 73, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Ziani, K.; Fernández-Pan, I.; Royo, M.; Maté, J.I. Antifungal activity of films and solutions based on chitosan against typical seed fungi. Food Hydrocoll. 2009, 23, 2309–2314. [Google Scholar] [CrossRef]
- Li, R.; Zhu, L.; Liu, D.; Wang, W.; Zhang, C.; Jiao, S.; Wei, J.; Ren, L.; Zhang, Y.; Gou, X. High molecular weight chitosan oligosaccharide exhibited antifungal activity by misleading cell wall organization via targeting PHR transglucosidases. Carbohydr. Polym. 2022, 285, 119253. [Google Scholar] [CrossRef] [PubMed]
- Attjioui, M.; Gillet, D.; El Gueddari, N.E.; Moerschbacher, B.M. Synergistic Antimicrobial Effect of Chitosan Polymers and Oligomers. Mol. Plant Microbe. Interact. 2021, 34, 770–778. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Kim, M.-H.; Park, S.-C.; Cheong, H.; Jang, M.-K.; Nah, J.-W.; Hahm, K.-S. Investigation of the Antifungal Activity and Mechanism of Action of LMWS-Chitosan. J. Microbiol. Biotechnol. 2008, 18, 1729–1734. [Google Scholar] [PubMed]
- Cord-Landwehr, S.; Moerschbacher, B.M. Deciphering the ChitoCode: Fungal chitins and chitosans as functional biopolymers. Fungal Biol. Biotechnol. 2021, 8, 1–8. [Google Scholar] [CrossRef]
- Ing, L.Y.; Zin, N.M.; Sarwar, A.; Katas, H. Antifungal activity of chitosan nanoparticles and correlation with their physical properties. Int. J. Biomater. 2012, 2012, 632698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kheiri, A.; Moosawi Jorf, S.A.; Malihipour, A.; Saremi, H.; Nikkhah, M. Synthesis and characterization of chitosan nanoparticles and their effect on Fusarium head blight and oxidative activity in wheat. Int. J. Biol. Macromol. 2017, 102, 526–538. [Google Scholar] [CrossRef]
- Ma, Z.; Lim, L.Y. Uptake of Chitosan and Associated Insulin in Caco-2 Cell Monolayers: A Comparison Between Chitosan Molecules and Chitosan Nanoparticles. Pharm. Res. 2003, 20, 1812–1819. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Wang, X.; Guo, X.; Yang, P.; Yu, J.; Shui, Y.; Chen, C.; Li, X.; Xu, Q.; Xu, L.; et al. Comparison of Antimicrobial Activity of Chitosan Nanoparticles against Bacteria and Fungi. Coatings 2021, 11, 769. [Google Scholar] [CrossRef]
- Wang, J.J.; Zeng, Z.W.; Xiao, R.Z.; Xie, T.; Zhou, G.L.; Zhan, X.R.; Wang, S.L. Recent advances of chitosan nanoparticles as drug carriers. Int. J. Nanomed. 2011, 6, 765–774. [Google Scholar]
- Abdel-Hafez, S.M.; Hathout, R.M.; Sammour, O.A. Towards better modeling of chitosan nanoparticles production: Screening different factors and comparing two experimental designs. Int. J. Biol. Macromol. 2014, 64, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Palma-Guerrero, J.; Lopez-Jimenez, J.A.; Perez-Berna, A.J.; Huang, I.C.; Jansson, H.B.; Salinas, J.; Villalain, J.; Read, N.D.; Lopez-Llorca, L.V. Membrane fluidity determines sensitivity of filamentous fungi to chitosan. Mol. Microbiol. 2010, 75, 1021–1032. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Moya, F.; Lopez-Llorca, L.V. Omics for Investigating Chitosan as an Antifungal and Gene Modulator. J. Fungi 2016, 2, 11. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Moya, F.; Suarez-Fernandez, M.; Lopez-Llorca, L.V. Molecular Mechanisms of Chitosan Interactions with Fungi and Plants. Int. J. Mol. Sci. 2019, 20, 332. [Google Scholar] [CrossRef] [Green Version]
- El Ghaouth, A.; Arul, J.; Grenier, J.; Asselin, A. Antifungal activity of chitosan on two postharvest pathogens of strawberry fruits. Phytopathology 1992, 82, 398–402. [Google Scholar] [CrossRef]
- Laflamme, P.; Benhamou, N.; Bussières, G.; Dessureault, M. Differential effect of chitosan on root rot fungal pathogens in forest nurseries. Can. J. Bot. 2000, 77, 1460–1468. [Google Scholar] [CrossRef]
- Allan, C.R.; Hadwiger, L.A. The fungicidal effect of chitosan on fungi of varying cell wall composition. Exp. Mycol. 1979, 3, 285–287. [Google Scholar] [CrossRef]
- Klis, F.M.; Ram, A.F.; De Groot, P.W. A molecular and genomic view of the fungal cell wall. In The Mycota VIII: Biology of the Fungal Cell; Klis, R.Z., Gow, N.A.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 97–120. [Google Scholar]
- Liu, J.; Tian, S.P.; Meng, X.H.; Xu, Y. Effects of chitosan on control of postharvest diseases and physiological responses of tomato fruit. Postharvest. Biol. Tec. 2007, 44, 300–306. [Google Scholar] [CrossRef]
- Palma-Guerrero, J.; Huang, I.-C.; Jansson, H.-B.; Salinas, J.; Lopez-Llorca, L.; Read, N. Chitosan permeabilizes the plasma membrane and kills cells of Neurospora crassa in an energy dependent manner. Fungal Genet. Biol. 2009, 46, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.H.; Hjeljord, L.G.; Aam, B.B.; Sorlie, M.; Tronsmo, A. Antifungal effect of chito-oligosaccharides with different degrees of polymerization. Eur. J. Plant Pathol. 2015, 141, 147–158. [Google Scholar] [CrossRef]
- Luan, J.; Wei, X.; Li, Z.; Tang, W.; Yang, F.; Yu, Z.; Li, X. Inhibition of Chitosan with Different Molecular Weights on Barley-Borne Fusarium graminearum during Barley Malting Process for Improving Malt Quality. Foods 2022, 11, 3058. [Google Scholar] [CrossRef] [PubMed]
- Loron, A.; Wang, Y.; Atanasova, V.; Richard-Forget, F.; Gardrat, C.; Coma, V. Chitosan for eco-friendly control of mycotoxinogenic Fusarium graminearum. Food Hydrocoll. 2023, 134, 108067. [Google Scholar] [CrossRef]
- Li, X.-F.; Feng, X.-Q.; Yang, S.; Wang, T.-P.; Su, Z.-X. Effects of MolecularWeight and Concentration of Chitosan on Antifungal Activity Against Aspergillus Niger. Iran. Polym. J. 2008, 17, 843–852. [Google Scholar]
- Onaran, A.; Bayar, Y.; Karakurt, T.; Tokatlı, K.; Bayram, M.; Yanar, Y. Antifungal activity of chitosan against soil-borne plant pathogens in cucumber and a molecular docking study. J. Taibah Univ. Sci. 2021, 15, 852–860. [Google Scholar] [CrossRef]
- Carmona, S.L.; Villarreal-Navarrete, A.; Burbano-David, D.; Gómez-Marroquín, M.; Torres-Rojas, E.; Soto-Suárez, M. Protection of tomato plants against Fusarium oxysporum f. sp. Lycopersici induced by chitosan. Rev. Colomb. De Cienc. Hortícolas 2021, 15, e12822. [Google Scholar] [CrossRef]
- Cord-Landwehr, S.; Richter, C.; Wattjes, J.; Sreekumar, S.; Singh, R.; Basa, S.; El Gueddari, N.E.; Moerschbacher, B.M. Patterns matter part 2: Chitosan oligomers with defined patterns of acetylation. React. Funct. Polym. 2020, 151, 104577. [Google Scholar] [CrossRef]
- Wattjes, J.; Sreekumar, S.; Richter, C.; Cord-Landwehr, S.; Singh, R.; El Gueddari, N.E.; Moerschbacher, B.M. Patterns matter part 1: Chitosan polymers with non-random patterns of acetylation. React. Funct. Polym. 2020, 151, 104583. [Google Scholar] [CrossRef]
- Basa, S.; Nampally, M.; Honorato, T.; Das, S.N.; Podile, A.R.; El Gueddari, N.E.; Moerschbacher, B.M. The pattern of acetylation defines the priming activity of chitosan tetramers. J. Am. Chem. Soc. 2020, 142, 1975–1986. [Google Scholar] [CrossRef] [PubMed]
- Sreekumar, S.; Wattjes, J.; Niehues, A.; Mengoni, T.; Mendes, A.C.; Morris, E.R.; Goycoolea, F.M.; Moerschbacher, B.M. Biotechnologically produced chitosans with nonrandom acetylation patterns differ from conventional chitosans in properties and activities. Nat. Commun. 2022, 13, 7125. [Google Scholar] [CrossRef] [PubMed]
- Lemke, P.; Junemann, L.; Moerschbacher, B.M. Synergistic Antimicrobial Activities of Chitosan Mixtures and Chitosan-Copper Combinations. Int. J. Mol. Sci. 2022, 23, 3345. [Google Scholar] [CrossRef]
- Wang, Q.Z.; Chen, X.G.; Liu, N.; Wang, S.X.; Liu, C.S.; Meng, X.H.; Liu, C.G. Protonation constants of chitosan with different molecular weight and degree of deacetylation. Carbohydr. Polym. 2006, 65, 194–201. [Google Scholar] [CrossRef]
- Alburquenque, C.; Bucarey, S.A.; Neira-Carrillo, A.; Urzua, B.; Hermosilla, G.; Tapia, C.V. Antifungal activity of low molecular weight chitosan against clinical isolates of Candida spp. Med. Mycol. 2010, 48, 1018–1023. [Google Scholar] [CrossRef] [Green Version]
- Philippova, O.E.; Korchagina, E.V.; Volkov, E.V.; Smirnov, V.A.; Khokhlov, A.R.; Rinaudo, M. Aggregation of some water-soluble derivatives of chitin in aqueous solutions: Role of the degree of acetylation and effect of hydrogen bond breaker. Carbohydr. Polym. 2012, 87, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Kulikov, S.; Tikhonov, V.; Blagodatskikh, I.; Bezrodnykh, E.; Lopatin, S.; Khairullin, R.; Philippova, Y.; Abramchuk, S. Molecular weight and pH aspects of the efficacy of oligochitosan against methicillin-resistant Staphylococcus aureus (MRSA). Carbohydr. Polym. 2012, 87, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.Y.; Hsu, C.H.; Huang, J.R.; Tsai, M.L.; Chen, R.H. Effect of the ionic strength of the media on the aggregation behaviors of high molecule weight chitosan. J. Polym. Res. 2011, 18, 1385–1395. [Google Scholar] [CrossRef]
- Mohammadpour Dounighi, N.; Eskandari, R.; Avadi, M.R.; Zolfagharian, H.; Mir Mohammad Sadeghi, A.; Rezayat, M. Preparation and in vitro characterization of chitosan nanoparticles containing Mesobuthus eupeus scorpion venom as an antigen delivery system. J. Venom. Anim. Toxins Incl. Trop. Dis. 2012, 18, 44–52. [Google Scholar] [CrossRef] [Green Version]
- Calvo, P.; Remuñan-López, C.; Vila-Jato, J.L.; Alonso, M.J. Chitosan and chitosan/ethylene oxide-propylene oxide block copolymer nanoparticles as novel carriers for proteins and vaccines. Pharm. Res. 1997, 14, 1431–1436. [Google Scholar] [CrossRef]
- Jameela, S.; Kumary, T.; Lal, A.; Jayakrishnan, A. Progesterone-loaded chitosan microspheres: A long acting biodegradable controlled delivery system. J. Control Release 1998, 52, 17–24. [Google Scholar] [CrossRef]
- Davis, S.; Illum, L. Sustained release chitosan microspheres prepared by novel spray drying methods. J. Microencapsul. 1999, 16, 343–355. [Google Scholar] [CrossRef]
- Tokumitsu, H.; Ichikawa, H.; Fukumori, Y. Chitosan-gadopentetic acid complex nanoparticles for gadolinium neutron-capture therapy of cancer: Preparation by novel emulsion-droplet coalescence technique and characterization. Pharm. Res. 1999, 16, 1830–1835. [Google Scholar] [CrossRef]
- Brunel, F.; Véron, L.; David, L.; Domard, A.; Delair, T. A novel synthesis of chitosan nanoparticles in reverse emulsion. Langmuir 2008, 24, 11370–11377. [Google Scholar] [CrossRef]
- Jonassen, H.; Kjøniksen, A.-L.; Hiorth, M. Stability of chitosan nanoparticles cross-linked with tripolyphosphate. Biomacromolecules 2012, 13, 3747–3756. [Google Scholar] [CrossRef] [PubMed]
- Sreekumar, S.; Goycoolea, F.M.; Moerschbacher, B.M.; Rivera-Rodriguez, G.R. Parameters influencing the size of chitosan-TPP nano-and microparticles. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Leon, T.; Carvalho, E.L.; Seijo, B.; Ortega-Vinuesa, J.L.; Bastos-Gonzalez, D. Physicochemical characterization of chitosan nanoparticles: Electrokinetic and stability behavior. J. Colloid Interface Sci. 2005, 283, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Cai, Y.H.; Lapitsky, Y. Factors affecting the stability of chitosan/tripolyphosphate micro- and nanogels: Resolving the opposing findings. J. Mater. Chem. B 2015, 3, 5957–5970. [Google Scholar] [CrossRef]
- Zhu, Y.; Marin, L.M.; Xiao, Y.; Gillies, E.R.; Siqueira, W.L. Ph-sensitive chitosan nanoparticles for salivary protein delivery. Nanomaterials 2021, 11, 1028. [Google Scholar] [CrossRef] [PubMed]
- Hassan, E.O.; Shoala, T.; Attia, A.M.; Badr, O.A.; Mahmoud, S.Y.; Farrag, E.S.; El-Fiki, I.A. Chitosan and nano-chitosan for management of Harpophora maydis: Approaches for investigating antifungal activity, pathogenicity, maize-resistant lines, and molecular diagnosis of plant infection. J. Fungi 2022, 8, 509. [Google Scholar] [CrossRef] [PubMed]
- Saharan, V.; Mehrotra, A.; Khatik, R.; Rawal, P.; Sharma, S.S.; Pal, A. Synthesis of chitosan based nanoparticles and their in vitro evaluation against phytopathogenic fungi. Int. J. Biol. Macromol. 2013, 62, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Gan, Q.; Wang, T.; Cochrane, C.; McCarron, P. Modulation of surface charge, particle size and morphological properties of chitosan-TPP nanoparticles intended for gene delivery. Colloids Surf. B Biointerfaces 2005, 44, 65–73. [Google Scholar] [CrossRef]
- El-Mohamedya, R.S.R.; Abd El-Aziz, M.E.; Kamel, S. Antifungal activity of chitosan nanoparticles against some plant pathogenic fungi in vitro. Agric. Eng. Int. CIGR J. 2019, 21, 201–209. [Google Scholar]
- Abdel-Aliem, H.A.; Gibriel, A.Y.; Rasmy, N.M.H.; Sahab, A.F.; El-Nekeety, A.A.; Abdel-Wahhab, M.A. Antifungal efficacy of chitosan nanoparticles against phytopathogenic fungi and inhibition of zearalenone production by Fusarium graminearum. Comun. Sci. 2019, 10, 338–345. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Rahman, F.A.; Monir, G.A.; Hassan, M.S.S.; Ahmed, Y.; Refaat, M.H.; Ismail, I.A.; El-Garhy, H.A.S. Exogenously Applied Chitosan and Chitosan Nanoparticles Improved Apple Fruit Resistance to Blue Mold, Upregulated Defense-Related Genes Expression, and Maintained Fruit Quality. Horticulturae 2021, 7, 224. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chitosan | Chitosan Nanoparticles | ||||||
|---|---|---|---|---|---|---|---|
| Molecular Weight [kDa] | Deacetylation [%] | Particle Size [nm] | Zeta Potential [mV] | Fungi Species | Concentration of CSNPs | The Rate of Growth Inhibition | References |
| 70 kDa | 75–85% | 174 nm | 39 mV | C. albicans | 0.25 [mg/mL] | 90% | [35] |
| F. solani | 1.0 [mg/mL] | 90% | |||||
| A. niger | - | 0% | |||||
| 233 nm | 38 mV | C. albicans | 0.85 [mg/mL] | 90% | |||
| F. solani | 0.85 [mg/mL] | 90% | |||||
| A. niger | - | 0% | |||||
| 255 nm | 48 mV | C. albicans | 0.60 [mg/mL] | 90% | |||
| F. solani | 1.21 [mg/mL] | 90% | |||||
| A. niger | - | 0% | |||||
| 310 kDa | 75% | 210 nm | 40 mV | C. albicans | 1.0 [mg/mL] | 90% | |
| F. solani | 0.5 [mg/mL] | 90% | |||||
| A. niger | - | 0% | |||||
| 263 nm | 52 mV | C. albicans | 0.85 [mg/mL] | 90% | |||
| F. solani | 0.85 [mg/mL] | 90% | |||||
| A. niger | 1.71 [mg/mL] | 90% | |||||
| 301 nm | 54 mV | C. albicans | 0.60 [mg/mL] | 90% | |||
| F. solani | 0.60 [mg/mL] | 90% | |||||
| A. niger | 2.42 [mg/mL] | 90% | |||||
| - | 85.61% | - | - | P.steckii | 5 [mg/mL] | 100% | [38] |
| A.oryzae | >5 [mg/mL] | 100% | |||||
| “Low molecular weight chitosan” | 40–70 nm | 48 mV | F. ox f. radicis lycopersici | 0.0125–0.1% | 48–100% | [80] | |
| F. oxysporum | 50–100% | ||||||
| F. solani | 50–100% | ||||||
| F. semibaticum | 46–100% | ||||||
| A. solani | 46–100% | ||||||
| P. infestance | 40–100% | ||||||
| R. solani | 51–100% | ||||||
| S. rolfsii | 32–100% | ||||||
| S. sclerotinum | 34–100% | ||||||
| B. cinerea | 39–100% | ||||||
| M. phaseolina | 43–100% | ||||||
| “High molecular weight chitosan” | 40–70 nm | 48 mV | F. ox f. radicis lycopersici | 0.0125–0.1% | 55–100% | ||
| F. oxysporum | 55–100% | ||||||
| F. solani | 58–100% | ||||||
| F. semibaticum | 52–100% | ||||||
| A. solani | 52–100% | ||||||
| P. infestance | 50–100% | ||||||
| R. solani | 55–100% | ||||||
| S. rolfsii | 39–100% | ||||||
| S. sclerotinum | 46–100% | ||||||
| B. cinerea | 43–100% | ||||||
| M. phaseolina | 53–100% | ||||||
| 244 kDa | 86.9% | 180 nm | - | A. tenuis | 0.01–0.08% | 20–68% | [81] |
| A. niger | 21–63% | ||||||
| A. terreus | 23–75% | ||||||
| B. bassiana | 74–76% | ||||||
| F. graminearum | 43–60% | ||||||
| F. oxysporum | 32–67% | ||||||
| S. rolfsii | 0–37% | ||||||
| - | 80% | 192.2 nm | 45.33 mV | A. alternata | 0.001–0.1% | 12–82% | [78] |
| M. phaseolina | 62–88% | ||||||
| R. solani | 13–34% | ||||||
| “Low molecular weight chitosan” | 180.9 nm | 45.6 mV | F. graminearum | 0.001–0.5% | 27 –75% | [36] | |
| 225.7 nm | 33.4 mV | 17–78% | |||||
| “Medium molecular weight chitosan” | 309.9 nm | 33.2 mV | 7–52% | ||||
| 301.5 nm | 20.2 mV | 7–53% | |||||
| “High molecular weight chitosan” | 339.4 nm | 21.7 mV | 7–52% | ||||
| 595.7 nm | 16 mV | 5–51% | |||||
| - | - | 100–160 nm | - | P. expansum | 0.2–0.4 [g/L] | 32–44% | [82] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poznanski, P.; Hameed, A.; Orczyk, W. Chitosan and Chitosan Nanoparticles: Parameters Enhancing Antifungal Activity. Molecules 2023, 28, 2996. https://doi.org/10.3390/molecules28072996
Poznanski P, Hameed A, Orczyk W. Chitosan and Chitosan Nanoparticles: Parameters Enhancing Antifungal Activity. Molecules. 2023; 28(7):2996. https://doi.org/10.3390/molecules28072996
Chicago/Turabian StylePoznanski, Pawel, Amir Hameed, and Waclaw Orczyk. 2023. "Chitosan and Chitosan Nanoparticles: Parameters Enhancing Antifungal Activity" Molecules 28, no. 7: 2996. https://doi.org/10.3390/molecules28072996