
Rhodococcus Strains from the Specialized Collection of Alkanotrophs for Biodegradation of Aromatic Compounds
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Toxicity of Aromatic Compounds for Rhodococcus
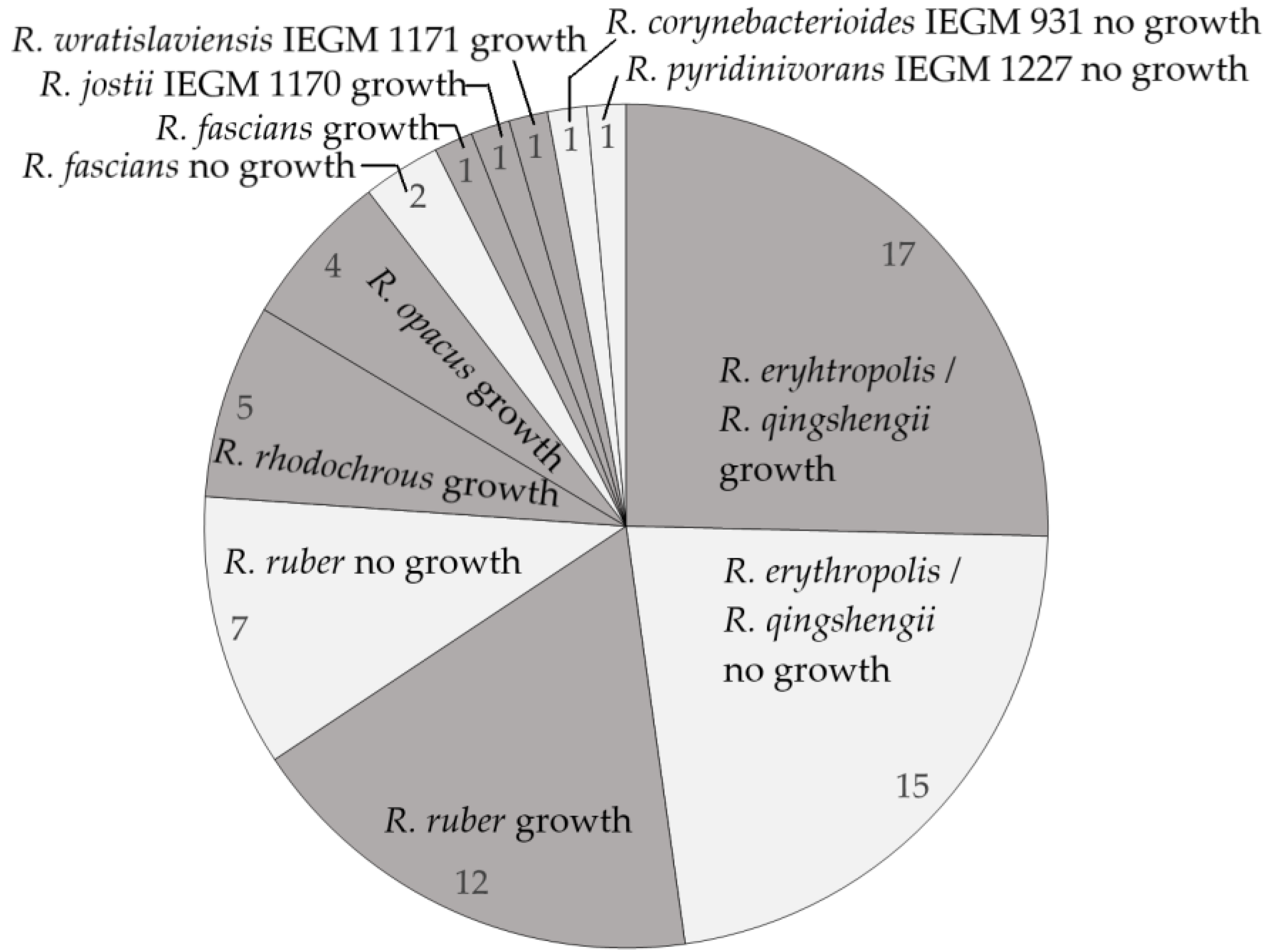
2.2. Degradation of Monoaromatic Hydrocarbons by Rhodococcus
2.3. Degradation of PAHs by Rhodococcus
2.4. Degradation of Substituted Aromatic Hydrocarbons and Aromatic Heterocycles by Rhodococcus
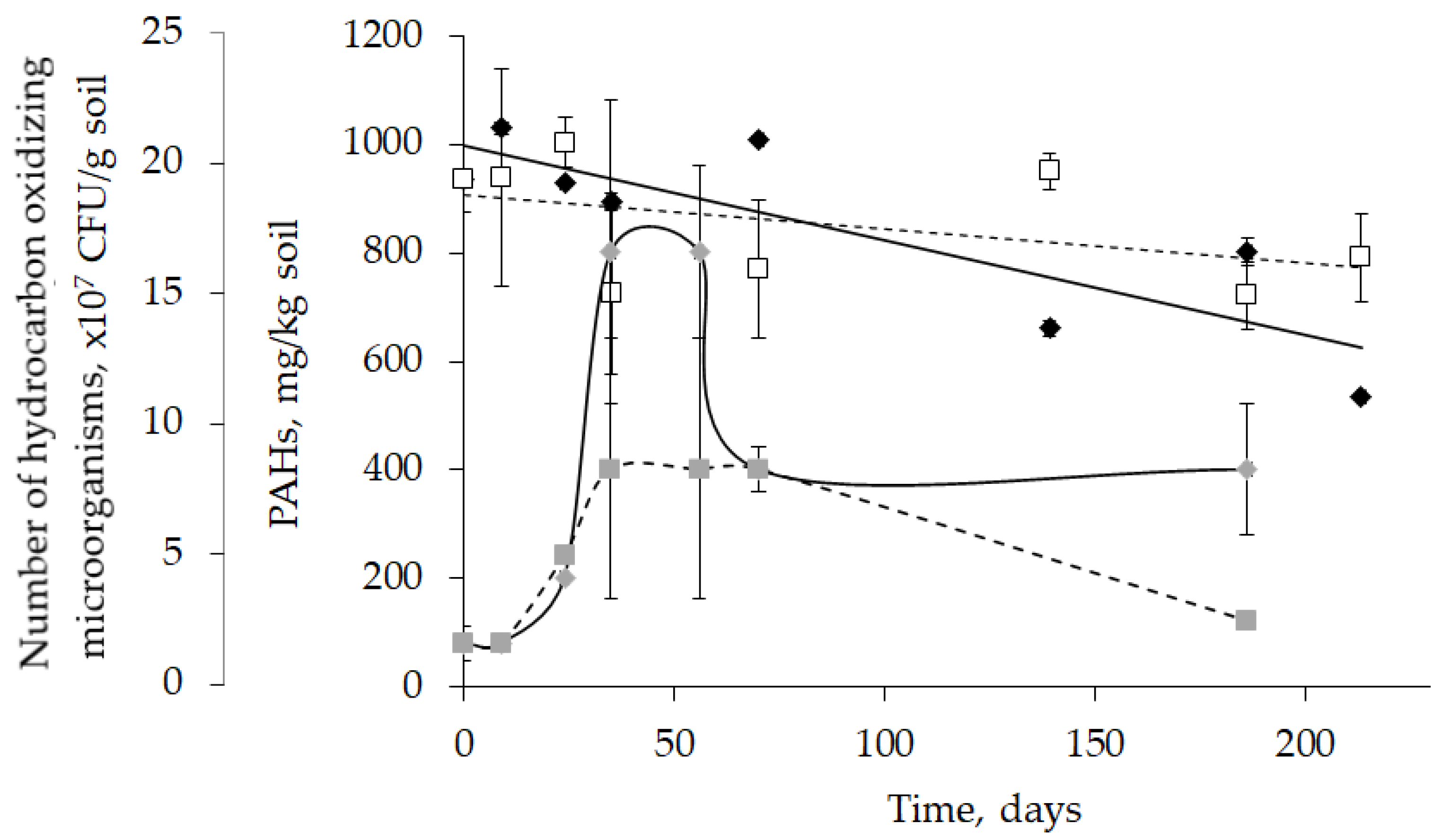
2.5. Biodegradation of Toxic Aromatic Compounds by Rhodococcus in Model Soil
2.6. Analysis of Rhodococcus Genes Involved in the Biodegradation of Aromatic Compounds
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Tested Aromatic Compounds
4.3. Toxicity Tests
4.4. Growth Experiments
4.5. Model Soil Experiments
4.6. Bioinformatical and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guo, H.; Yao, J.; Chen, H.; Wang, J.; Masakorala, K.; Jin, Y.; Richnowd, H.H.; Blake, R.E. Substrate interactions during biodegradation of benzene/alkylbenzene mixtures by Rhodococcus sp. ustb-1. Int. Biodeterior. Biodegrad. 2012, 75, 124–130. [Google Scholar] [CrossRef]
- You, Y.; Shim, J.; Cho, C.-H.; Ryu, M.-H.; Shea, P.J.; Kamala-Kannan, S.; Chae, J.-C.; Oh, B.-T. Biodegradation of BTEX mixture by Pseudomonas putida YNS1 isolated from oil-contaminated soil. J. Basic Microbiol. 2013, 53, 469–475. [Google Scholar] [CrossRef]
- Li, H.; Meng, F.; Duan, W.; Lin, Y.; Zheng, Y. Biodegradation of phenol in saline or hypersaline environments by bacteria: A review. Exotoxicol. Environ. Saf. 2019, 184, 109658. [Google Scholar] [CrossRef] [PubMed]
- Hlisníková, H.; Petrovičová, I.; Kolena, B.; Šidlovská, M.; Sirotkin, A. Effects and mechanisms of phthalates’ action on reproductive processes and reproductive health: A literature review. Int. J. Environ. Res. Public Health 2020, 17, 6811. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, M.; Mekala, L.P.; Chintalapati, S.; Chintalapati, V.R. New insights into aniline toxicity: Aniline exposure triggers envelope stress and extracellular polymeric substance formation in Rubrivivax benzoatilyticus JA2. J. Hazard. Mater. 2020, 385, 121571. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Gong, L.; Zhang, Y.; Tong, Y.; Zhang, H.; Zhu, D.; Huang, X.; Yang, H. Bioaugmentation potential evaluation of a bacterial consortium composed of isolated Pseudomonas and Rhodococcus for degrading benzene, toluene and styrene in sludge and sewage. Bioresour. Technol. 2021, 320, 124329. [Google Scholar] [CrossRef]
- Görmez, Ö.; Akay, S.; Gözmen, B.; Kayan, B.; Kalderis, D. Degradation of emerging contaminant coumarin based on anodic oxidation, electro-Fenton and subcritical water oxidation processes. Environ. Res. 2022, 208, 112736. [Google Scholar] [CrossRef] [PubMed]
- Tao, N.; Zhang, W.; Si, L.; Zhang, R.; Wang, D.; Guo, C. Effects of aniline on growth, oxidative and DNA damage of rice (Oryza sativa L.) seedlings. Environ. Technol. Innov. 2022, 28, 102583. [Google Scholar] [CrossRef]
- Zheng, P.; Li, Y.; Chi, Q.; Cheng, Y.; Jiang, X.; Chen, D.; Mu, Y.; Shen, J. Structural characteristics and microbial function of biofilm in membrane-aerated biofilm reactor for the biodegradation of volatile pyridine. J. Hazard. Mater. 2022, 437, 129370. [Google Scholar] [CrossRef]
- Zhihan, Z.; Bofeng, Z.; Dong, W.; Qi, Z.; Cong, T.; Nanwen, Z.; Guobiao, L. A Yb3+/Er3+ co-doped Bi1.95Yb0.04Er0.01V2O8 efficient upconversion glass-ceramic photocatalyst for antibiotic degradation driven by UV–Vis-NIR broad spectrum light. Appl. Surf. Sci. 2022, 583, 152565. [Google Scholar] [CrossRef]
- Ali, M.; Song, X.; Wang, Q.; Zhang, Z.; Che, J.; Chen, X.; Tang, Z.; Liu, X. Mechanisms of biostimulant-enhanced biodegradation of PAHs and BTEX mixed contaminants in soil by native microbial consortium. Environ. Pollut. 2023, 318, 120831. [Google Scholar] [CrossRef] [PubMed]
- Balasubramaniyan, B.; Usha Mary, T.M.; Swaminathan, M. Microbial degradation of quinoline by immobilized Bacillus subtilis. Biocatal. Agric. Biotechnol. 2023, 47, 102604. [Google Scholar] [CrossRef]
- Bird, M.G.; Greim, H.; Kaden, D.A.; Rice, J.M.; Snyder, R. BENZENE 2009—Health effects and mechanisms of bone marrow toxicity: Implications for t-AML and the mode of action framework. Chem. Biol. Interact. 2010, 184, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Azubuike, C.C.; Chikere, C.B.; Okpokwasili, G.C. Bioremediation techniques-classification based on site of application: Principles, advantages, limitations and prospects. World J. Microbiol. Biotechnol. 2016, 32, 180. [Google Scholar] [CrossRef] [Green Version]
- Krivoruchko, A.V.; Kuyukina, M.S.; Ivshina, I.B. Advanced Rhodococcus biocatalysts for environmental biotechnologies. Catalysts 2019, 9, 236. [Google Scholar] [CrossRef] [Green Version]
- Kuyukina, M.S.; Ivshina, I.B. Bioremediation of contaminated environments using Rhodococcus. In Biology of Rhodococcus, 2nd ed.; Alvarez, H.M., Ed.; Microbiology Monographs; Springer Nature Switzerland AG: Cham, Switzerland, 2019; Volume 16, pp. 231–270. [Google Scholar] [CrossRef]
- Mohapatra, B.; Dhamale, T.; Saha, B.K.; Phale, P.S. Microbial degradation of aromatic pollutants: Metabolic routes, pathway diversity, and strategies for bioremediation. In Microbial Biodegradation and Bioremediation: Techniques and Case Studies for Environmental Pollution, 2nd ed.; Das, S., Dash, H.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 365–394. [Google Scholar] [CrossRef]
- Pérez-Pantoja, D.; Donoso, R.; Agulló, L.; Córdova, M.; Seeger, M.; Pieper, D.H.; González, B. Genomic analysis of the potential for aromatic compounds biodegradation in Burkholderiales. Environ. Microbiol. 2012, 14, 1091–1117. [Google Scholar] [CrossRef]
- Lee, Y.; Lee, Y.; Jeon, C.O. Biodegradation of naphthalene, BTEX, and aliphatic hydrocarbons by Paraburkholderia aromaticivorans BN5 isolated from petroleum-contaminated soil. Sci. Rep. 2019, 9, 860. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Hu, Q.; Qiu, Z.; Yu, C. Biodegradation characteristics and mechanism of quinoline by Ochrobactrum sp. strain C2. Water Sci. Technol. 2022, 86, 1284–1298. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Qi, H.; Zhang, X.M. Co-biodegradation of anthracene and naphthalene by the bacterium Acinetobacter johnsonii. J. Environ. Sci. Health A Tox Hazard. Subst. Environ. Eng. 2018, 53, 448–456. [Google Scholar] [CrossRef]
- Parab, V.; Phadke, M. Co-biodegradation studies of naphthalene and phenanthrene using bacterial consortium. J. Environ. Sci. Health A Tox Hazard. Subst. Environ. Eng. 2020, 55, 912–924. [Google Scholar] [CrossRef]
- Luo, Y.; Yue, X.; Wei, P.; Zhou, A.; Kong, X.; Alimzhanova, S. A state-of-the-art review of quinoline degradation and technical bottlenecks. Sci. Total Environ. 2020, 747, 141136. [Google Scholar] [CrossRef]
- Zhu, G.; Xing, F.; Tao, J.; Chen, S.; Li, K.; Cao, L.; Yan, N.; Zhang, Y.; Rittmann, B.E. Synergy of strains that accelerate biodegradation of pyridine and quinoline. J. Environ. Manag. 2021, 285, 112119. [Google Scholar] [CrossRef]
- Huang, S.; Wang, M.; Mao, D.; Rasool, A.; Jia, C.; Yang, P.; Han, L.; Yan, M. Isolation, identification and characterization of growth parameters of Pseudomonas putida HSM-C2 with coumarin-degrading bacteria. Molecules 2022, 27, 6007. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, F.; Xu, S.; Yang, P.; Wang, X.; Zhang, X.; Hong, Q.; Qiu, J.; Chu, C.; He, J. Biodegradation of quinoline by a newly isolated salt-tolerating bacterium Rhodococcus gordoniae strain JH145. Microorganisms 2022, 10, 797. [Google Scholar] [CrossRef]
- Pátek, M.; Grulich, M.; Nešvera, J. Stress response in Rhodococcus strains. Biotechnol. Adv. 2021, 53, 107698. [Google Scholar] [CrossRef]
- Ivshina, I.B.; Kuyukina, M.S.; Krivoruchko, A.V.; Tyumina, E.A. Responses to ecopollutants and pathogenization risks of saprotrophic Rhodococcus species. Pathogens 2021, 10, 974. [Google Scholar] [CrossRef]
- Ivshina, I.; Bazhutin, G.; Tyumina, E. Rhodococcus strains as a good biotool for neutralizing pharmaceutical pollutants and obtaining therapeutically valuable products: Through the past into the future. Front. Microbiol. 2022, 13, 967127. [Google Scholar] [CrossRef] [PubMed]
- Nazari, M.T.; Simon, V.; Machado, B.S.; Crestani, L.; Marchezi, G.; Concolato, G.; Ferrari, V.; Colla, L.M.; Piccin, J.S. Rhodococcus: A promising genus of actinomycetes for the bioremediation of organic and inorganic contaminants. J. Environ. Manag. 2022, 323, 116220. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Niu, C.; Lu, Z. Individual or synchronous biodegradation of di-n-butyl phthalate and phenol by Rhodococcus ruber strain DP-2. J. Hazard. Mater. 2014, 273, 104–109. [Google Scholar] [CrossRef]
- Choi, K.Y.; Kang, B.S.; Nam, M.H.; Sul, W.J.; Kim, E. Functional identification of OphR, an IclR family transcriptional regulator involved in the regulation of the phthalate catabolic operon in Rhodococcus sp. strain DK17. Indian J. Microbiol. 2015, 55, 313–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Dong, W.; Wang, H.; Zhao, Y.; Zhao, Z.; Huang, J.; Zhou, T.; Wu, Z.; Li, W. Enhanced bioremediation of sediment contaminated with polycyclic aromatic hydrocarbons by combined stimulation with sodium acetate/phthalic acid. Chemosphere 2022, 291, 132770. [Google Scholar] [CrossRef] [PubMed]
- Ivshina, I.B.; Krivoruchko, A.V.; Kuyukina, M.S.; Peshkur, T.A.; Cunningham, C.J. Adhesion of Rhodococcus bacteria to solid hydrocarbons and enhanced biodegradation of these compounds. Sci. Rep. 2022, 12, 21559. [Google Scholar] [CrossRef]
- Shahabivand, S.; Mortazavi, S.S.; Mahdavinia, G.R.; Darvishi, F. Phenol biodegradation by immobilized Rhodococcus qingshengii isolated from coking effluent on Na-alginate and magnetic chitosan-alginate nanocomposite. J. Environ. Manag. 2022, 307, 114586. [Google Scholar] [CrossRef] [PubMed]
- Ivshina, I.B.; Kuyukina, M.S.; Krivoruchko, A.V. Hydrocarbon-oxidizing bacteria and their potential in eco-biotechnology and bioremediation. In Microbial Resources: From Functional Existence in Nature to Industrial Applications, 1st ed.; Kurtböke, I., Ed.; Academic Press Elsevier: Cambridge, MA, USA, 2017; pp. 121–148. [Google Scholar] [CrossRef]
- Declerck, S.W.A.; van der Heijden, M.G.A.; Varese, G.C.; Turkovskaya, O.V.; Evtushenko, L.I.; Ivshina, I.B.; Desmeth, P. PERN: An EU-Russia initiative for rhizosphere microbial resources. Trends Biotechnol. 2015, 33, 377–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivshina, I.B.; Kuyukina, M.S.; Krivoruchko, A.V.; Barbe, V.; Fischer, C. Draft genome sequence of propane- and butane-oxidizing actinobacterium Rhodococcus ruber IEGM 231. Genome Announc. 2014, 2, e01297-14. [Google Scholar] [CrossRef] [Green Version]
- Othman, H.B.; Pick, F.R.; Hlaili, A.S.; Leboulanger, C. Effects of polycyclic aromatic hydrocarbons on marine and freshwater microalgae—A review. J. Hazard. Mater. 2023, 441, 129869. [Google Scholar] [CrossRef]
- Huang, J.; Ling, J.; Kuang, C.; Chen, J.; Xu, Y.; Li, Y. Microbial biodegradation of aniline at low concentrations by Pigmentiphaga daeguensis isolated from textile dyeing sludge. Int. Biodeterior. Biodegrad. 2018, 129, 117–122. [Google Scholar] [CrossRef]
- Hu, R.; Zhao, H.; Xu, X.; Wang, Z.; Yu, K.; Shu, L.; Yan, Q.; Wu, B.; Mo, C.; He, Z.; et al. Bacteria-driven phthalic acid ester biodegradation: Current status and emerging opportunities. Environ. Int. 2021, 154, 106560. [Google Scholar] [CrossRef]
- Stes, E.; Francis, E.; Pertry, I.; Dolzblasz, A.; Depuydt, S.; Vereecke, D. The leafy gall syndrome induced by Rhodococcus fascians. FEMS Microbiol. Lett. 2013, 342, 187–194. [Google Scholar] [CrossRef] [Green Version]
- Tomás-Gallardo, L.; Gómez-Álvarez, H.; Santero, E.; Floriano, B. Combination of degradation pathways for naphthalene utilization in Rhodococcus sp. strain TFB. Microb. Biotechnol. 2014, 7, 100–113. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Chen, Y.; Zhou, H.; Li, X.; Tan, Z. Adaptation mechanisms of Rhodococcus sp. CNS16 under different temperature gradients: Physiological and transcriptome. Chemosphere 2020, 238, 124571. [Google Scholar] [CrossRef] [PubMed]
- Gakhar, L.; Malik, Z.A.; Allen, C.C.; Lipscomb, D.A.; Larkin, M.J.; Ramaswamy, S. Structure and increased thermostability of Rhodococcus sp. naphthalene 1,2-dioxygenase. J. Bacteriol. 2005, 187, 7222–7231. [Google Scholar] [CrossRef] [Green Version]
- Baratto, M.C.; Lipscomb, D.A.; Larkin, M.J.; Basosi, R.; Allen, C.C.R.; Pogni, R. Spectroscopic characterisation of the naphthalene dioxygenase from Rhodococcus sp. strain NCIMB12038. Int. J. Mol. Sci. 2019, 20, 3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anokhina, T.O.; Esikova, T.Z.; Gafarov, A.B.; Polivtseva, V.N.; Baskunov, B.P.; Solyanikova, I.P. Alternative naphthalene metabolic pathway includes formation of ortho-phthalic acid and cinnamic acid derivatives in the Rhodococcus opacus strain 3D. Biochemistry 2020, 85, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Gibson, D.T.; Parales, R.E. Aromatic hydrocarbon dioxygenases in environmental biotechnology. Curr. Opin. Biotechnol. 2000, 11, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Mate, D.M.; Alcalde, M. Laccase: A multi-purpose biocatalyst at the forefront of biotechnology. Microb. Biotechnol. 2017, 10, 1457–1467. [Google Scholar] [CrossRef] [Green Version]
- Lopes, J.M.; Marques-da-Silva, D.; Videira, P.Q.; Lago, R.L. Comparison of laccases and hemeproteins systems in bioremediation of organic pollutants. Curr. Protein Pept. Sci. 2022, 23, 402–423. [Google Scholar] [CrossRef]
- Ivshina, I.B.; Kuyukina, M.S.; Krivoruchko, A.V.; Plekhov, O.A.; Naimark, O.B.; Podorozhko, E.A.; Lozinsky, V.I. Biosurfactant-enhanced immobilization of hydrocarbon-oxidizing Rhodococcus ruber on sawdust. Appl. Microbiol. Biotechnol. 2013, 97, 5315–5327. [Google Scholar] [CrossRef]
- Ivshina, I.B.; Tyumina, E.A.; Bazhutin, G.A.; Vikhareva, E.V. Response of Rhodococcus cerastii IEGM 1278 to toxic effects of ibuprofen. PLoS ONE 2021, 16, e0260032. [Google Scholar] [CrossRef]
- Ivshina, I.B.; Tyumina, E.A.; Kuzmina, M.V.; Vikhareva, E.V. Features of diclofenac biodegradation by Rhodococcus ruber IEGM 346. Sci. Rep. 2019, 9, 9159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.D.; Kim, I.S. Rhodococcus spelaei sp. nov., isolated from a cave, and proposals that Rhodococcus biphenylivorans is a later synonym of Rhodococcus pyridinivorans, Rhodococcus qingshengii and Rhodococcus baikonurensis are later synonyms of Rhodococcus erythropolis, and Rhodococcus percolatus and Rhodococcus imtechensis are later synonyms of Rhodococcus opacus. Int. J. Syst. Evol. Microbiol. 2021, 71, 4890. [Google Scholar] [CrossRef]
- Wrenn, B.A.; Venosa, A.D. Selective enumeration of aromatic and aliphatic hydrocarbon degrading bacteria by a most-probable-number procedure. Can. J. Microbiol. 1996, 42, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Ivshina, I.B.; Kuyukina, M.S.; Ritchkova, M.I.; Philp, J.C.; Cunningham, C.J.; Christofi, N. Oleophilic biofertilizer based on a Rhodococcus surfactant complex for the bioremediation of crude oil-contaminated soil. In Proceedings of the 1st International Congress on Petroleum Contaminated Soils, Sediments and Water, London, UK, 14–17 August 2001. [Google Scholar]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid Annotations using Subsystems Technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aromatic Compound | MIC, мM | Aromatic Compound | MIC, мM |
|---|---|---|---|
| Benzene | 0.2 | Phenol | 25.0 |
| Toluene | 0.8 | Aniline | 25.0 |
| o-Xylene | 25.0 | Pyridine | 50.0 |
| Naphthalene | 50.0 | 2-Picoline | 50.0 |
| Anthracene | 50.0 | 3-Picoline | 50.0 |
| Phenanthrene | 50.0 | 4-Picoline | 50.0 |
| Benzo[a]anthracene | 50.0 | 2,6-Lutidine | 50.0 |
| Benzo[a]pyrene | 50.0 | 2-Hydroxipyridine | 50.0 |
| o-Phthalic acid | 50.0 | 4-Hydroxipyridine | 50.0 |
| m-Phthalic acid | 0.2 | Quinoline | 6.3 |
| p-Phthalic acid | 0.2 | Coumarin | 50.0 |
| Salicylic acid | 50.0 |
| Species | Number of Strains | ||
|---|---|---|---|
| Total | Growing | Not Growing | |
| Phenol | |||
| R. erythropolis/R. qingshengii | 30 | 4 | 26 |
| R. corynebacterioides | 1 1 | 0 | 1 |
| R. fascians | 4 | 0 | 4 |
| R. jostii | 1 1 | 0 | 1 |
| R. opacus | 1 1 | 0 | 1 |
| R. pyridinivorans | 1 1 | 0 | 1 |
| R. rhodochrous | 2 | 2 | 0 |
| R. ruber | 16 | 3 | 13 |
| R. wratislaviensis | 1 | 0 | 1 |
| Total | 57 | 9 | 1 |
| Pyridine | |||
| R. aetherivorans | 1 | 0 | 1 |
| R. erythropolis/R. qingshengii | 2 | 0 | 2 |
| R. jostii | 1 2 | 0 | 1 |
| R. pyridinivorans | 1 2 | 0 | 1 |
| R. rhodochrous | 3 | 1 | 2 |
| R. ruber | 2 | 1 | 1 |
| Total | 9 | 2 | 7 |
| Strains | R. qingshengii IEGM 267 | R. qingshengii IEGM 746 | R. erythropolis IEGM 1189 | R. qingshengii IEGM 1359 | R. opacus IEGM 249 | R. pyridinivorans IEGM 1137 | R. rhodochrous IEGM 107 | R. rhodochrous IEGM 757 | R. rhodochrous IEGM 1360 | R. ruber IEGM 231 |
| Growth substrates | o-Xylene, PAHs | o-Xylene | Toluene, o-xylene | Benzene, o-xylene | Naphthalene | - | - | Pyridine | Benzene | Benzene, toluene, PAHs, aniline, pyridine, coumarin |
| Number of genes coded for | ||||||||||
| Dioxygenase (total) | 35 | 30 | 28 | 30 | 69 | 23 | 32 | 41 | 32 | 46 |
| Catechol 1,2-dioxygenase | 1 | 1 | 1 | 1 | 4 | 1 | 2 | 3 | 2 | 2 |
| Protocatechuate 3,4-dioxygenase | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 |
| Extradiol dioxygenase | 2 | 2 | 2 | 2 | 2 | 1 | 2 | 1 | 2 | 6 |
| Catechol 2,3-dioxygenase | 0 | 0 | 0 | 0 | 2 | 1 | 0 | 0 | 0 | 1 |
| Gentisate 1,2-dioxygenase | 0 | 0 | 0 | 0 | 3 | 0 | 1 | 2 | 1 | 1 |
| Homogentisate 1,2-dioxygenase | 1 | 2 | 1 | 2 | 1 | 1 | 2 | 2 | 2 | 1 |
| Benzene 1,2-dioxygenase | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Anthranilate 1,2-dioxygenase reductase | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 1 |
| 3-Hydroxy anthranilate 3,4-dioxygenase | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Monooxygenase (total) | 66 | 66 | 66 | 66 | 97 | 45 | 63 | 74 | 63 | 72 |
| Phenol monooxygenase | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 |
| Benzoyl-CoA oxygenase/reductase | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 |
| Phenylacetyl-CoA oxygenase/reductase | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 4-Hydroxybenzoate 3-hydroxylase | 1 | 1 | 1 | 1 | 4 | 2 | 3 | 3 | 3 | 1 |
| 3-Hydroxybenzoate 6-hydroxylase | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Salicylate 1-hydroxylase | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 1 | 0 |
| Multicopper polyphenol oxidase | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 4 |
| Dehydrogenase (total) | 387 | 367 | 350 | 348 | 731 | 256 | 352 | 429 | 348 | 288 |
| Aldehyde dehydrogenase | 13 | 13 | 11 | 12 | 41 | 6 | 15 | 20 | 15 | 5 |
| Succinate-semialdehyde dehydrogenase | 1 | 1 | 1 | 1 | 5 | 1 | 1 | 0 | 1 | 1 |
| Hydrolase | 125 | 116 | 116 | 116 | 229 | 96 | 115 | 133 | 115 | 123 |
| Phenol hydrolase | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 |
| Amidase | 2 | 2 | 2 | 2 | 2 | 0 | 0 | 0 | 0 | 3 |
| Formamidase | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 |
| Fumarylacetoacetate hydrolase family | 1 | 1 | 0 | 1 | 6 | 0 | 2 | 2 | 2 | 0 |
| Hydroxymuconic semialdehyde hydrolase | 3 | 2 | 2 | 2 | 0 | 1 | 2 | 2 | 2 | 1 |
| Vanillate 3-O demethylase | 4 | 2 | 2 | 4 | 20 | 4 | 7 | 8 | 7 | 5 |
| Maleate isomerase | 0 | 0 | 1 | 0 | 2 | 0 | 0 | 0 | 0 | 0 |
| Strain | A Complex Aromatic Compound, Biodegradation Efficiency | Reference |
|---|---|---|
| R. cerastii IEGM 1278 | (RS)-2-(4-(2-Methylpropyl)phenyl)propanoic acid (ibuprofen), 100% degradation of 100 mg/L in the presence of 0.1% (v/v) n-hexadecane within 6 days | [52] |
| R. ruber IEGM 346 | [2-(2,6-Dichloroanilino)phenyl]acetic acid (diclofenac), 100% degradation of 0.05 mg/L in the presence of 0.5% (w/w) D-glucose within 6 days | [53] |
| Group of Aromatic Compounds | Compound | Solvent | Heating at Dissolution |
|---|---|---|---|
| Monocyclic aromatic hydrocarbons | Benzene | DMSO | No |
| Toluene | DMSO | No | |
| o-Xylene | DMSO | No | |
| PAHs | Naphthalene | Acetone | No |
| Anthracene | Acetone | 70 °C | |
| Phenanthrene | Acetone | No | |
| Benzo[a]anthracene | Acetone | 70 °C | |
| Benzo[a]pyrene | Acetone | 70 °C | |
| Aromatic compounds with hydroxy groups | Phenol | Water | No |
| Aromatic amines | Aniline | DMSO | No |
| N-heterocyclic aromatic compounds | Pyridine | Water | No |
| 2-Picoline | Water | No | |
| 3-Picoline | Water | No | |
| 4-Picoline | Water | No | |
| 2,6-Lutidine | Water | No | |
| 2-Hydroxipyridine | Water | No | |
| 4-Hydroxipyridine | Water | No | |
| Quinoline | 70% ethanol | No | |
| Aromatic acids | o-Phthalic acid | Acetone | No |
| m-Phthalic acid | Acetone | No | |
| p-Phthalic acid | Acetone | 70 °C | |
| Salicylic acid | Acetone | 70 °C | |
| Aromatic lactones | Coumarin | Acetone | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krivoruchko, A.; Kuyukina, M.; Peshkur, T.; Cunningham, C.J.; Ivshina, I. Rhodococcus Strains from the Specialized Collection of Alkanotrophs for Biodegradation of Aromatic Compounds. Molecules 2023, 28, 2393. https://doi.org/10.3390/molecules28052393
Krivoruchko A, Kuyukina M, Peshkur T, Cunningham CJ, Ivshina I. Rhodococcus Strains from the Specialized Collection of Alkanotrophs for Biodegradation of Aromatic Compounds. Molecules. 2023; 28(5):2393. https://doi.org/10.3390/molecules28052393
Chicago/Turabian StyleKrivoruchko, Anastasiia, Maria Kuyukina, Tatyana Peshkur, Colin J. Cunningham, and Irina Ivshina. 2023. "Rhodococcus Strains from the Specialized Collection of Alkanotrophs for Biodegradation of Aromatic Compounds" Molecules 28, no. 5: 2393. https://doi.org/10.3390/molecules28052393