
The (Poly)phenolic Profile of Separate Winery By-Products Reveals Potential Antioxidant Synergies
 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
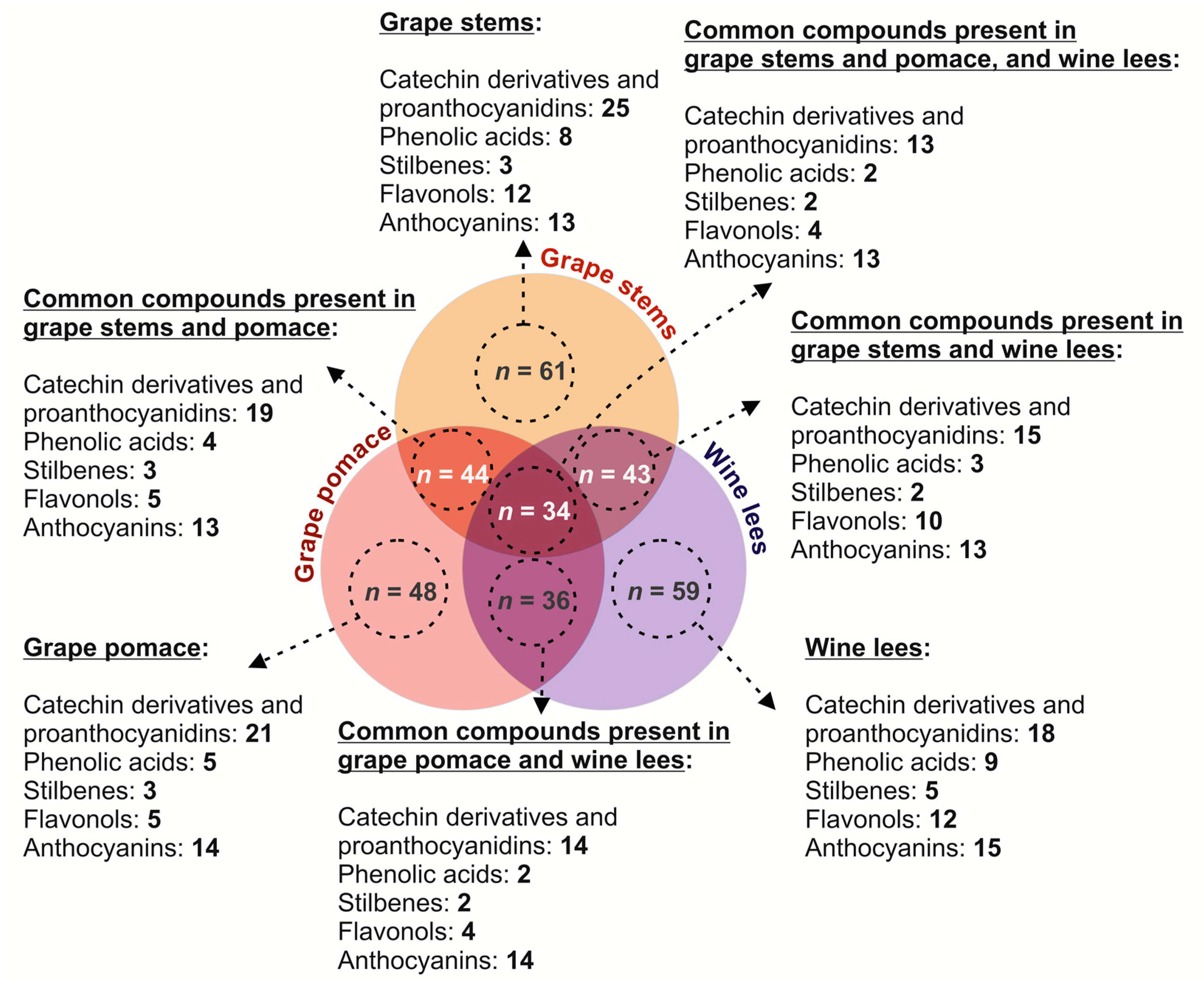
2.1. Comparative Analysis of the (Poly)phenolic Profile of Grape Stems, Grape Pomace, and Wine Lees
2.1.1. Catechin Derivatives and Proanthocyanidins
2.1.2. Phenolic Acids
2.1.3. Stilbenes
2.1.4. Flavonols
2.1.5. Anthocyanins
2.2. Quantitative (Poly)phenolic Content of Grape Stems, Grape Pomace, and Wine Lees
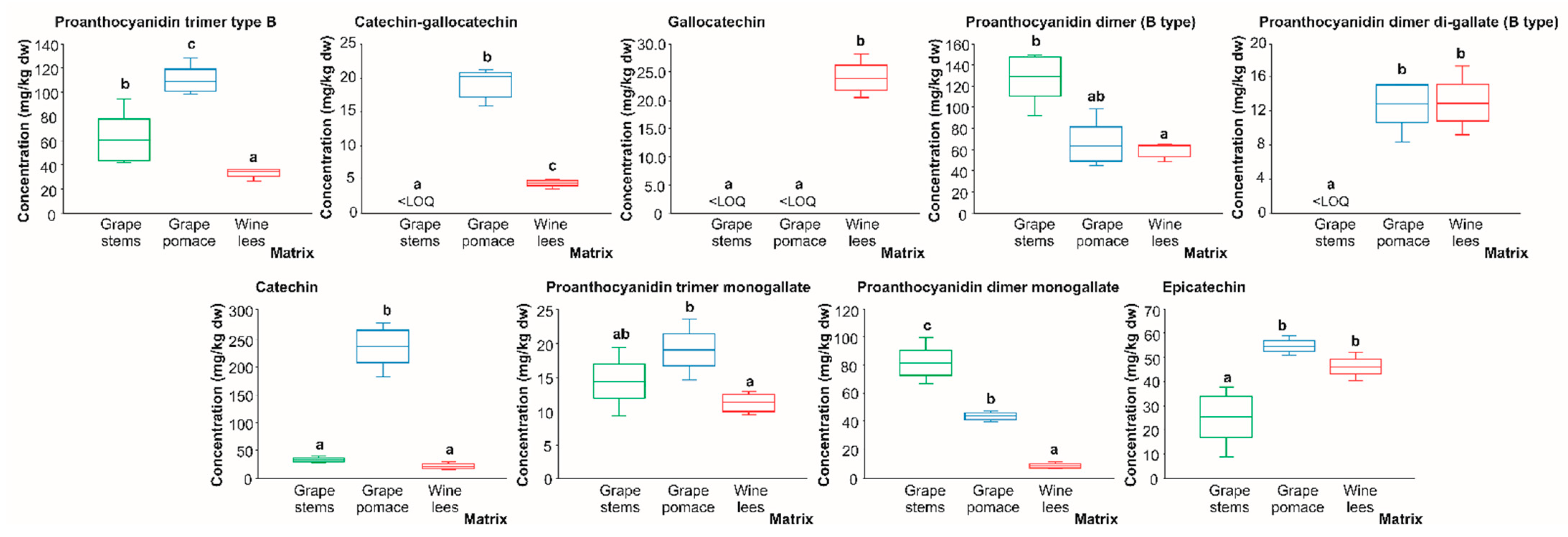
2.2.1. Catechin Derivatives and Proanthocyanidins
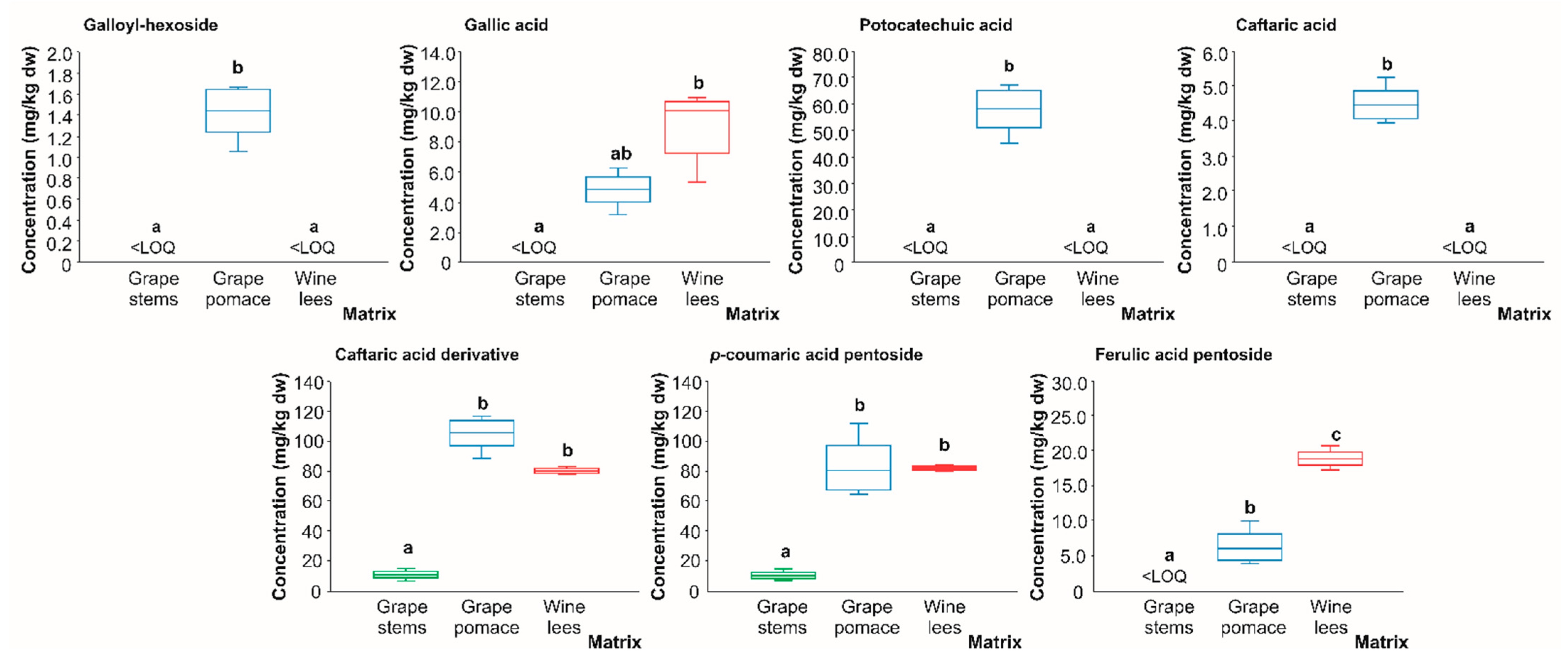
2.2.2. Phenolic Aids
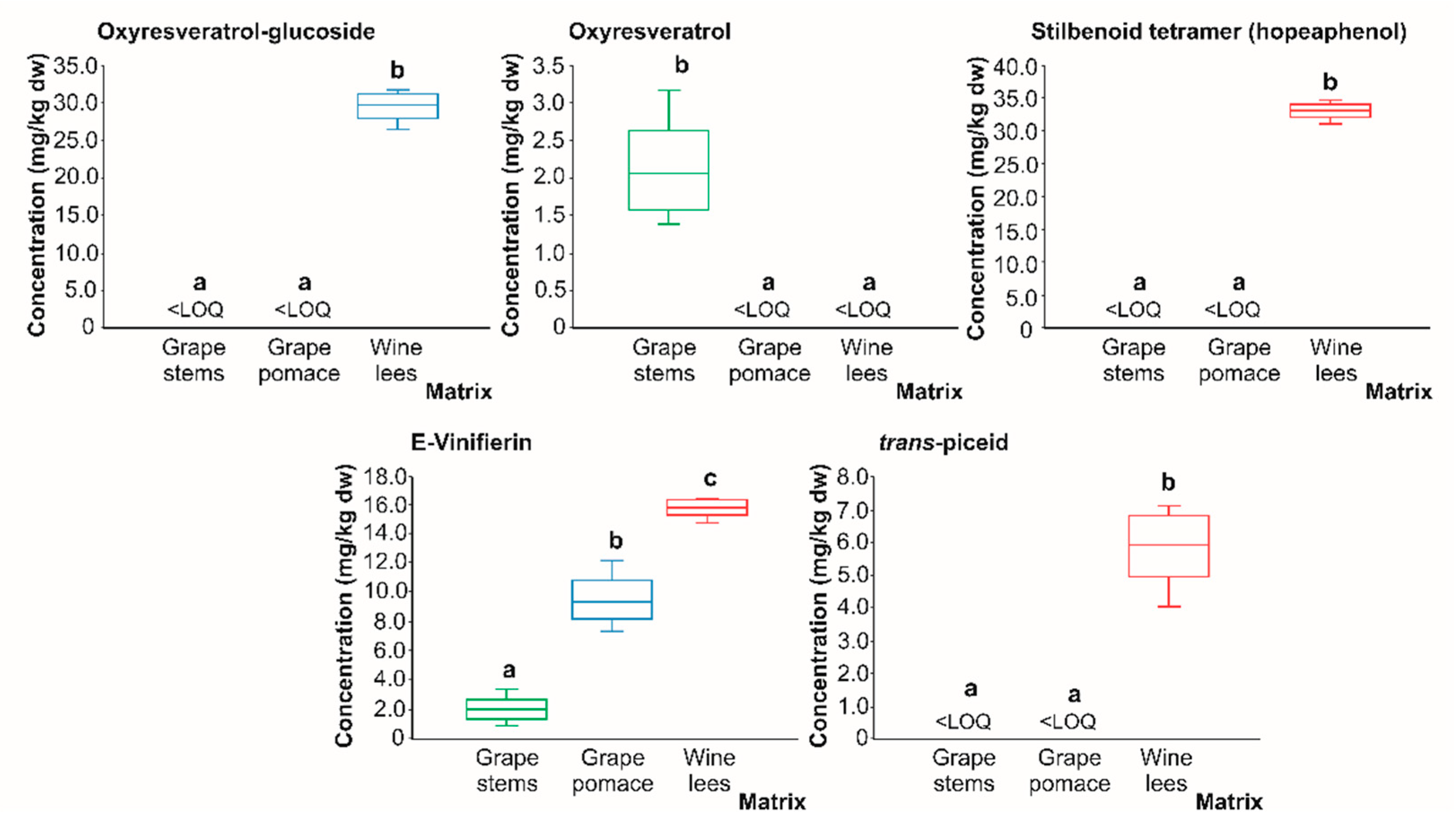
2.2.3. Stilbenes
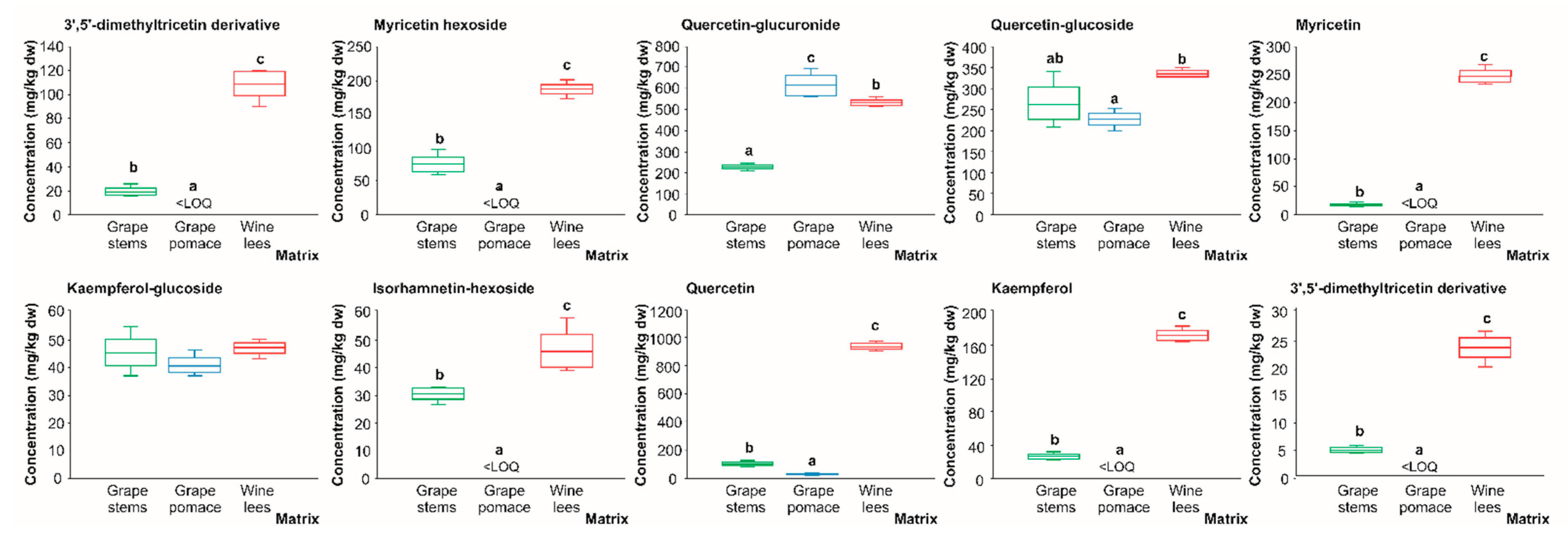
2.2.4. Flavonols
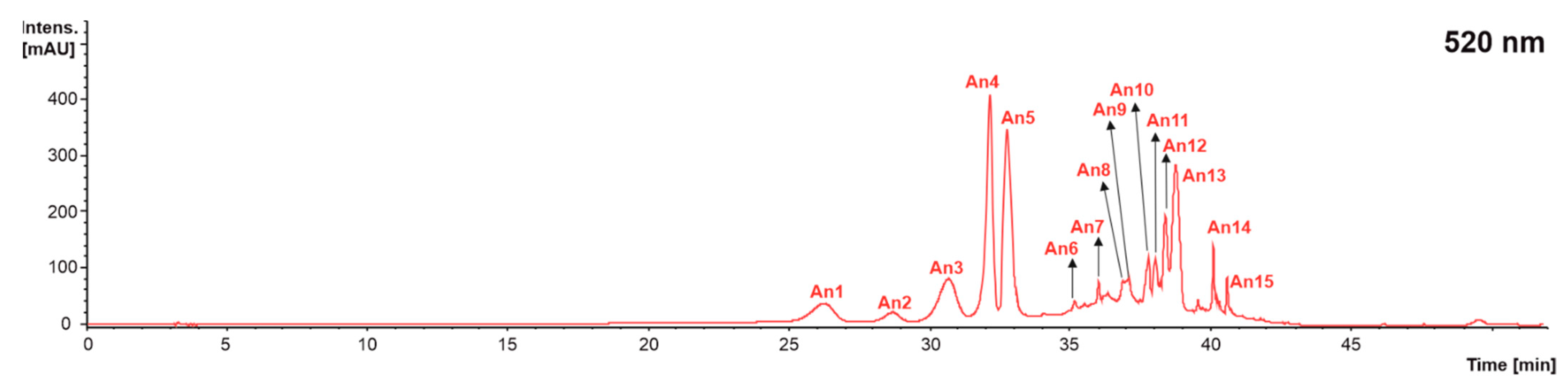
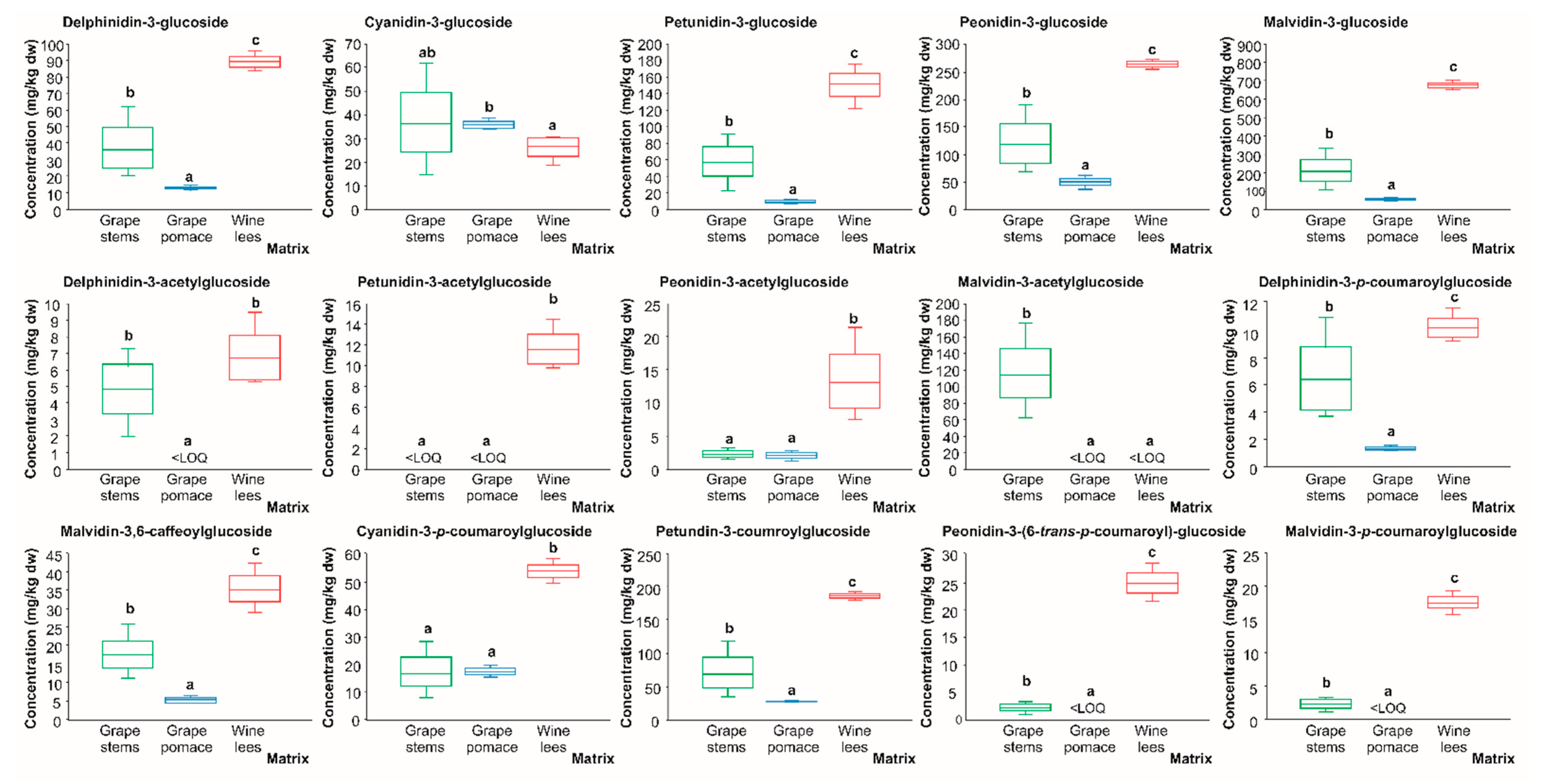
2.2.5. Anthocyanins
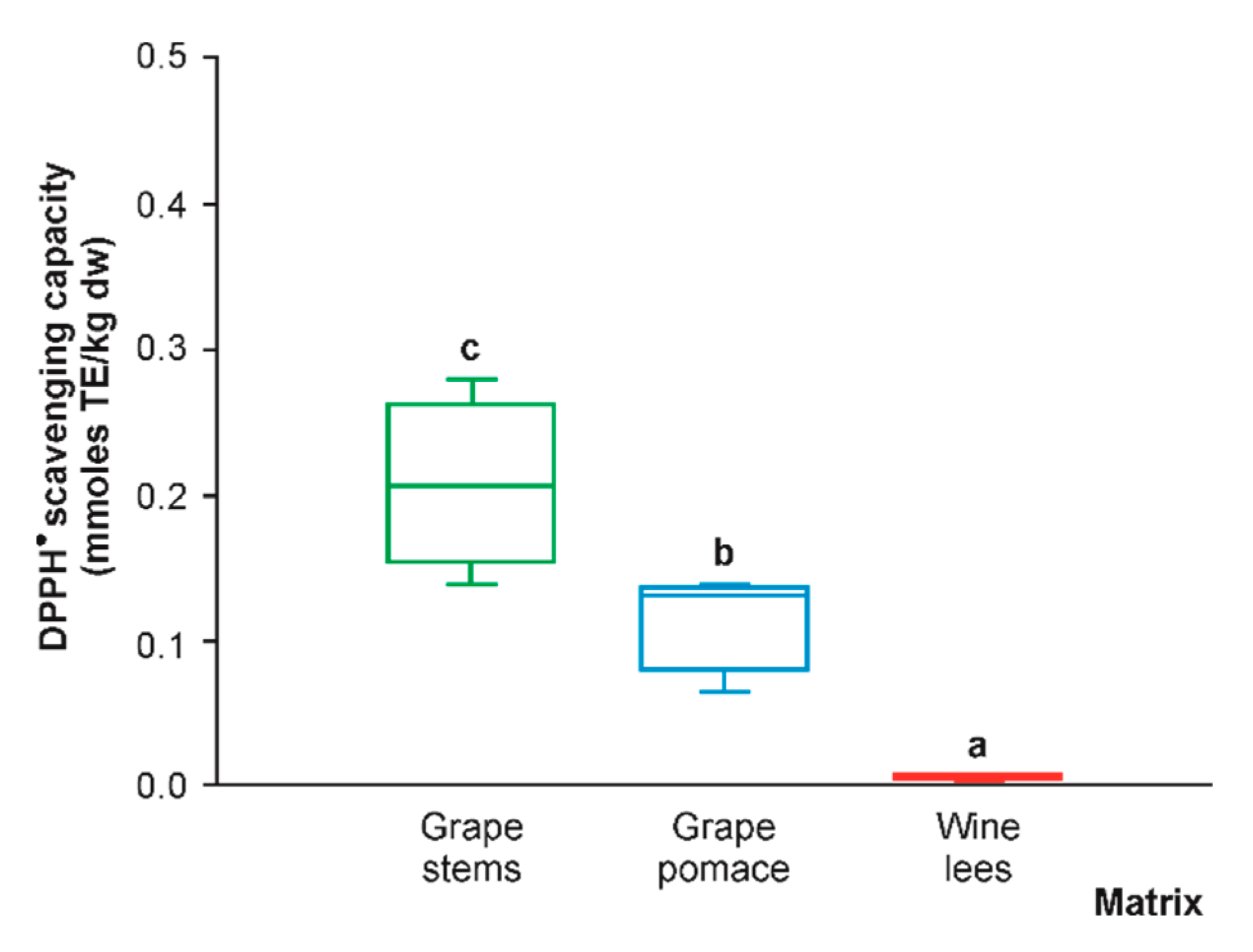
2.3. Differential DPPH● Scavenging Activity and Correlation Analysis
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Plant Material
3.3. Qualitative and Quantitative (Poly)phenolic Profile by HPLC-PDA-ESI-MSn
3.4. Antioxidant Capacity
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Machado, N.F.L.; Domínguez-Perles, R. Addressing Facts and Gaps in the Phenolics Chemistry of Winery By-Products. Molecules 2017, 22, 286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibn Ferjani, A.; Jellali, S.; Akrout, H.; Limousy, L.; Hamdi, H.; Thevenin, N.; Jeguirim, M. Nutrient Retention and Release from Raw Exhausted Grape Marc Biochars and an Amended Agricultural Soil: Static and Dynamic Investigation. Env. Technol. Innov. 2020, 19, 100885. [Google Scholar] [CrossRef]
- Domínguez-Perles, R.; Moreno, D.A.; García-Viguera, C. Waking Up from Four Decades’ Long Dream of Valorizing Agro-Food Byproducts: Toward Practical Applications of the Gained Knowledge. J. Agric. Food Chem. 2018, 66, 3069–3073. [Google Scholar] [CrossRef]
- Beres, C.; Costa, G.N.S.; Cabezudo, I.; da Silva-James, N.K.; Teles, A.S.C.; Cruz, A.P.G.; Mellinger-Silva, C.; Tonon, R.V.; Cabral, L.M.C.; Freitas, S.P. Towards Integral Utilization of Grape Pomace from Winemaking Process: A Review. Waste Manag. 2017, 68, 581–594. [Google Scholar] [CrossRef]
- Silva, V.; Igrejas, G.; Falco, V.; Santos, T.P.; Torres, C.; Oliveira, A.M.P.; Pereira, J.E.; Amaral, J.S.; Poeta, P. Chemical Composition, Antioxidant and Antimicrobial Activity of Phenolic Compounds Extracted from Wine Industry by-Products. Food Control 2018, 92, 516–522. [Google Scholar] [CrossRef] [Green Version]
- Ye, Z.; Qin, Y.; Harrison, R.; Hider, R.; Bekhit, A.E.D.A. Characterization of Bioactive Compounds in Lees from New Zealand Wines with Different Vinification Backgrounds. Antioxidants 2022, 11, 2335. [Google Scholar] [CrossRef] [PubMed]
- Mir-Cerdà, A.; Carretero, I.; Coves, J.R.; Pedrouso, A.; Castro-Barros, C.M.; Alvarino, T.; Cortina, J.L.; Saurina, J.; Granados, M.; Sentellas, S. Recovery of Phenolic Compounds from Wine Lees Using Green Processing: Identifying Target Molecules and Assessing Membrane Ultrafiltration Performance. Sci. Total Env. 2023, 857, 159623. [Google Scholar] [CrossRef] [PubMed]
- Boudet, A.M. Evolution and Current Status of Research in Phenolic Compounds. Phytochemistry 2007, 68, 2722–2735. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Perles, R.; Teixeira, A.I.; Rosa, E.; Barros, A.I. Assessment of (Poly)Phenols in Grape (Vitis Vinifera L.) Stems by Using Food/Pharma Industry Compatible Solvents and Response Surface Methodology. Food Chem. 2014, 164, 339–346. [Google Scholar] [CrossRef]
- Domínguez-Perles, R.; Guedes, A.; Queiroz, M.; Silva, A.M.; Barros, A.I.R.N.A. Oxidative Stress Prevention and Anti-Apoptosis Activity of Grape (Vitis Vinifera L.) Stems in Human Keratinocytes. Food Res. Int. 2016, 87, 92–102. [Google Scholar] [CrossRef]
- Queiroz, M.; Oppolzer, D.; Gouvinhas, I.; Silva, A.M.; Barros, A.I.R.N.A.; Domínguez-Perles, R. New Grape Stems’ Isolated Phenolic Compounds Modulate Reactive Oxygen Species, Glutathione, and Lipid Peroxidation in vitro: Combined Formulations with Vitamins C and E. Fitoterapia 2017, 120, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Dias, C.; Domínguez-Perles, R.; Aires, A.; Teixeira, A.; Rosa, E.; Barros, A.; Saavedra, M.J. Phytochemistry and Activity against Digestive Pathogens of Grape (Vitis Vinifera L.) Stem’s (Poly)Phenolic Extracts. LWT–Food Sci. Technol. 2015, 61, 25–32. [Google Scholar] [CrossRef]
- Gouvinhas, I.; Santos, R.A.; Queiroz, M.; Leal, C.; Saavedra, M.J.; Domínguez-Perles, R.; Rodrigues, M.; Barros, A.I.R.N.A. Monitoring the Antioxidant and Antimicrobial Power of Grape (Vitis Vinifera L.) Stems Phenolics over Long-Term Storage. Int. J. Cardiol. 2018, 126, 83–91. [Google Scholar] [CrossRef]
- Barros, A.; Gironés-Vilaplana, A.; Teixeira, A.; Collado-González, J.; Moreno, D.A.; Gil-Izquierdo, A.; Rosa, E.; Domínguez-Perles, R. Evaluation of Grape (Vitis Vinifera L.) Stems from Portuguese Varieties as a Resource of (Poly)Phenolic Compounds: A Comparative Study. Food Res. Int. 2014, 65, 375–384. [Google Scholar] [CrossRef]
- Remigante, A.; Spinelli, S.; Basile, N.; Caruso, D.; Falliti, G.; Dossena, S.; Marino, A.; Morabito, R. Oxidation Stress as a Mechanism of Aging in Human Erythrocytes: Protective Effect of Quercetin. Int. J. Mol. Sci. 2022, 23, 7781. [Google Scholar] [CrossRef]
- Csekes, E.; Račková, L. Skin Aging, Cellular Senescence and Natural Polyphenols. Int. J. Mol. Sci. 2021, 22, 12641. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.B.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxidative Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [Green Version]
- Barcia, M.T.; Pertuzatti, P.B.; Rodrigues, D.; Gómez-Alonso, S.; Hermosín-Gutiérrez, I.; Godoy, H.T. Occurrence of Low Molecular Weight Phenolics in Vitis Vinifera Red Grape Cultivars and Their Winemaking By-Products from São Paulo (Brazil). Food Res. Int. 2014, 62, 500–513. [Google Scholar] [CrossRef]
- Barcia, M.T.; Pertuzatti, P.B.; Gómez-Alonso, S.; Godoy, H.T.; Hermosín-Gutiérrez, I. Phenolic Composition of Grape and Winemaking By-Products of Brazilian Hybrid Cultivars BRS Violeta and BRS Lorena. Food Chem. 2014, 159, 95–105. [Google Scholar] [CrossRef]
- Romero-Díez, R.; Rodríguez-Rojo, S.; Cocero, M.J.; Duarte, C.M.M.; Matias, A.A.; Bronze, M.R. Phenolic Characterization of Aging Wine Lees: Correlation with Antioxidant Activities. Food Chem. 2018, 259, 188–195. [Google Scholar] [CrossRef] [Green Version]
- Dujmić, F.; Kovačević, K.; Ganićgani, K.; Curić, D.C.; Karlović, S.K.; Bosiljkov, T.; Ježek, D.; Vidrih, R.; Hribar, J.; Zlatić, E.Z.; et al. Non-Thermal Ultrasonic Extraction of Polyphenolic Compounds from Red Wine Lees. Foods 2020, 9, 472. [Google Scholar] [CrossRef] [Green Version]
- López-Fernández-Sobrino, R.; Margalef, M.; Torres-Fuentes, C.; Ávila-Román, J.; Aragonès, G.; Muguerza, B.; Bravo, F.I. Enzyme-Assisted Extraction to Obtain Phenolic-Enriched Wine Lees with Enhanced Bioactivity in Hypertensive Rats. Antioxidants 2021, 10, 517. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, N.; Azevedo, J.; Mateus, N.; de Freitas, V. Proanthocyanidin Screening by LC-ESI-MS of Portuguese Red Wines Made with Teinturier Grapes. Food Chem. 2016, 190, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Kumar, S.; Kumar, B. LC-MS Identification of Proanthocyanidins in Bark and Fruit of Six Terminalia Species. Nat. Prod. Commun. 2018, 13, 555–560. [Google Scholar] [CrossRef] [Green Version]
- Delcambre, A.; André, Y.; Saucier, C. Sequencing of Red Wine Proanthocyanidins by UHPLC-ESI-Q-ToF. J. Appl. Bioanal. 2015, 1, 46–54. [Google Scholar] [CrossRef]
- Rockenbach, I.I.; Jungfer, E.; Ritter, C.; Santiago-Schübel, B.; Thiele, B.; Fett, R.; Galensa, R. Characterization of Flavan-3-ols in Seeds of Grape Pomace by CE, HPLC-DAD-MSn and LC-ESI-FTICR-MS. Food Res. Int. 2012, 48, 848–855. [Google Scholar] [CrossRef] [Green Version]
- Aguilar-Zárate, P.; Wong-Paz, J.E.; Michel, M.; Buenrostro-Figueroa, J.; Díaz, H.R.; Ascacio, J.A.; Contreras-Esquivel, J.C.; Gutiérrez-Sánchez, G.; Aguilar, C.N. Characterisation of Pomegranate-Husk Polyphenols and Semi-Preparative Fractionation of Punicalagin. Phytochem. Anal. 2017, 28, 433–438. [Google Scholar] [CrossRef]
- Ali, A.; Bashmil, Y.M.; Cottrell, J.J.; Suleria, H.A.R.; Dunshea, F.R. LC-MS/MS-Qtof Screening and Identification of Phenolic Compounds from Australian Grown Herbs and Their Antioxidant Potential. Antioxidants 2021, 10, 1770. [Google Scholar] [CrossRef]
- De Iseppi, A.; Lomolino, G.; Marangon, M.; Curioni, A. Current and Future Strategies for Wine Yeast Lees Valorization. Food Res. Int. 2020, 137, 109352. [Google Scholar] [CrossRef]
- Quifer-Rada, P.; Vallverdú-Queralt, A.; Martínez-Huélamo, M.; Chiva-Blanch, G.; Jáuregui, O.; Estruch, R.; Lamuela-Raventós, R. A Comprehensive Characterisation of Beer Polyphenols by High Resolution Mass Spectrometry (LC–ESI-LTQ-Orbitrap-MS). Food Chem. 2015, 169, 336–343. [Google Scholar] [CrossRef]
- Milinčić, D.D.; Stanisavljević, N.S.; Kostić, A.; Soković Bajić, S.; Kojić, M.O.; Gašić, U.M.; Barać, M.B.; Stanojević, S.P.; Tešić, Ž.L.; Pešić, M.B. Phenolic Compounds and Biopotential of Grape Pomace Extracts from Prokupac Red Grape Variety. Food Sci. Technol. 2021, 138, 110739. [Google Scholar] [CrossRef]
- Souza da Costa, B.; Soldevilla Muro, G.; Oliván García, M.; Motilva, M.J. Winemaking By-Products as a Source of Phenolic Compounds: Comparative Study of Dehydration Processes. Food Sci. Technol. 2022, 165, 113774. [Google Scholar] [CrossRef]
- Vallverdú-Queralt, A.; Boix, N.; Piqué, E.; Gómez-Catalan, J.; Medina-Remon, A.; Sasot, G.; Mercader-Martí, M.; Llobet, J.M.; Lamuela-Raventos, R.M. Identification of Phenolic Compounds in Red Wine Extract Samples and Zebrafish Embryos by HPLC-ESI-LTQ-Orbitrap-MS. Food Chem. 2015, 181, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Monagas, M.; Bartolomé, B.; Gómez-Cordovés, C. Evolution of Polyphenols in Red Wines from Vitis Vinifera L. during Aging in the Bottle: IIII. Non-Anthocyanin Phenolic Compounds. Eur. Food Res. Technol. 2005, 220, 331–340. [Google Scholar] [CrossRef]
- Chen, W.; Yeo, S.C.M.; Elhennawy, M.G.A.A.; Lin, H.S. Oxyresveratrol: A Bioavailable Dietary Polyphenol. J. Funct. Foods 2016, 22, 122–131. [Google Scholar] [CrossRef]
- Chiva-Blanch, G.; Urpi-Sarda, M.; Rotchés-Ribalta, M.; Zamora-Ros, R.; Llorach, R.; Lamuela-Raventós, R.M.; Estruch, R.; Andrés-Lacueva, C. Determination of Resveratrol and Piceid in Beer Matrices by Solid-Phase Extraction and Liquid Chromatography-Tandem Mass Spectrometry. J. Chromatogr. A 2011, 1218, 698–705. [Google Scholar] [CrossRef]
- Moss, R.; Mao, Q.; Taylor, D.; Saucier, C. Investigation of Monomeric and Oligomeric Wine Stilbenoids in Red Wines by Ultra-High-Performance Liquid Chromatography/Electrospray Ionization Quadrupole Time-of-Flight Mass Spectrometry. Rapid Commun. Mass Spectrom. 2013, 27, 1815–1827. [Google Scholar] [CrossRef]
- Fuchs, C.; Bakuradze, T.; Steinke, R.; Grewal, R.; Eckert, G.P.; Richling, E. Polyphenolic Composition of Extracts from Winery By-Products and Effects on Cellular Cytotoxicity and Mitochondrial Functions in HepG2 Cells. J. Funct. Foods 2020, 70, 103988. [Google Scholar] [CrossRef]
- Guebailia, H.A.; Chira, K.; Richard, T.; Mabrouk, T.; Furiga, A.; Vitrac, X.; Monti, J.P.; Delaunay, J.C.; Mérillon, J.M. Hopeaphenol: The First Resveratrol Tetramer in Wines from North Africa. J. Agricult. Food Chem. 2006, 54, 9559–9564. [Google Scholar] [CrossRef]
- Gouveia, S.; Castilho, P.C. Characterisation of Phenolic Acid Derivatives and Flavonoids from Different Morphological Parts of Helichrysum Obconicum by a RP-HPLC-DAD-(-)-ESI-MSn Method. Food Chem. 2011, 129, 333–344. [Google Scholar] [CrossRef]
- Jaiswal, R.; Jayasinghe, L.; Kuhnert, N. Identification and Characterization of Proanthocyanidins of 16 Members of the Rhododendron Genus (Ericaceae) by Tandem LC-MS. J. Mass. Spectrom. 2012, 47, 502–515. [Google Scholar] [CrossRef] [PubMed]
- Simirgiotis, M.J. Antioxidant Capacity and HPLC-DAD-MS Profiling of Chilean Peumo (Cryptocarya Alba) Fruits and Comparison with German Peumo (Crataegus Monogyna) from Southern Chile. Molecules 2013, 18, 2061–2080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panchal, H.; Shah, M.B. Development and Validation of a Rapid LC-MS/MS Method for Simultaneous Determination of Kaempferol and Quercetin in Thespesia Populnea Extract. J. AOAC Int. 2017, 100, 971–975. [Google Scholar] [CrossRef] [PubMed]
- Schieber, A.; Keller, P.; Streker, P.; Klaiber, I.; Carle, R. Detection of Isorhamnetin Glycosides in Extracts of Apples (Malus Domestica Cv. “Brettacher”) by HPLC-PDA and HPLC-APCI-MS/MS. Phytochem. Anal. 2002, 13, 87–94. [Google Scholar] [CrossRef]
- Trikas, E.D.; Papi, R.M.; Kyriakidis, D.A.; Zachariadis, G.A. A Sensitive LC-MS Method for Anthocyanins and Comparison of Byproducts and Equivalent Wine Content. Separations 2016, 3, 18. [Google Scholar] [CrossRef]
- Ou, K.; Sarnoski, P.; Schneider, K.R.; Song, K.; Khoo, C.; Gu, L. Microbial Catabolism of Procyanidins by Human Gut Microbiota. Mol. Nutr. Food Res. 2014, 58, 2196–2205. [Google Scholar] [CrossRef]
- Díaz, N.; Aqueveque, P.M.; Vallejos-Almirall, A.; Radrigán, R.; Zúñiga-López, M.C.; Folch-Cano, C. Antioxidant Compound Adsorption in Polyvinylpolypyrrolidone from Chilean Carménère, Cabernet Sauvignon, and Merlot Grape Pomaces as Potential By-Products. Antioxidants 2022, 11, 2017. [Google Scholar] [CrossRef]
- Romano, P.; Capece, A. Chapter 13: Wine Microbiology in Starter Cultures in Food Prodiction; Speranza, B., Bevilacqua, A., Corbo, M.R., Sinigaglia, M., Eds.; Wiley: Hoboken, NJ, USA, 2017; pp. 255–282. [Google Scholar] [CrossRef]
- Stafford, H.A. Proanthocyanidins and the Lignin Connection. Phytochemistry 1988, 27, 1–6. [Google Scholar] [CrossRef]
- De Freitas, V.A.; Glories, Y. Concentration and Compositional Changes of Procyanidins in Grape Seeds and Skin of White Vitis Vinõ Âfera Varieties. J. Sci. Food Agric. 1999, 79, 1601–1606. [Google Scholar] [CrossRef]
- Gonzaíez-Centeno, R.M.; Jourdes, M.; Femenia, A.; Simal, S.; Rosselló, C.R.; Teissedre, P.-L. Characterization of Polyphenols and Antioxidant Potential of White Grape Pomace Byproducts (Vitis Vinifera L.). J. Agric. Food Chem. 2013, 61, 11579–11587. [Google Scholar] [CrossRef]
- Gombau, J.; Pons-Mercadé, P.; Conde, M.; Asbiro, L.; Pascual, O.; Gómez-Alonso, S.; García-Romero, E.; Miquel Canals, J.; Hermosín-Gutiérrez, I.; Zamora, F. Influence of Grape Seeds on Wine Composition and Astringency of Tempranillo, Garnacha, Merlot and Cabernet Sauvignon Wines. Food Sci. Nutr. 2020, 8, 3442–3455. [Google Scholar] [CrossRef]
- Jiménez, N.; Esteban-Torres, M.; Mancheño, J.M.; de las Rivas, B.; Muñoza, R. Tannin Degradation by a Novel Tannase Enzyme Present in Some Lactobacillus Plantarum Strains. Appl. Environ. Microbiol. 2014, 80, 2991–2997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tork, S.; Qari, H.; Zainal, H.; Aly, M. View of Biodegradation of Tannins from Polluted Sources Using Natural Enzymes. Kuwait J. Sci. 2020, 47, 82–91. Available online: https://journalskuwait.org/kjs/index.php/KJS/article/view/8777/395 (accessed on 11 January 2023).
- Chávez-González, M.; Rodríguez-Durán, L.V.; Balagurusamy, N.; Prado-Barragán, A.; Rodríguez, R.; Contreras, J.C.; Aguilar, C.N. Biotechnological Advances and Challenges of Tannase: An Overview. Food Bioproc. Tech. 2012, 5, 445–459. [Google Scholar] [CrossRef]
- Cappello, M.S.; Zapparoli, G.; Logrieco, A.; Bartowsky, E.J. Linking Wine Lactic Acid Bacteria Diversity with Wine Aroma and Flavour. Int. J. Food Microbiol. 2017, 243, 16–27. [Google Scholar] [CrossRef]
- Teixeira, A.; Baenas, N.; Dominguez-Perles, R.; Barros, A.; Rosa, E.; Moreno, D.A.; Garcia-Viguera, C. Natural Bioactive Compounds from Winery By-Products as Health Promoters: A Review. Int. J. Mol. Sci. 2014, 15, 15638–15678. [Google Scholar] [CrossRef] [Green Version]
- Leonard, W.; Zhang, P.; Ying, D.; Adhikari, B.; Fang, Z. Fermentation Transforms the Phenolic Profiles and Bioactivities of Plant-Based Foods. Biotechnol. Adv. 2021, 49, 107763. [Google Scholar] [CrossRef]
- Rodríguez, H.; Landete, J.M.; de las Rivas, B.; Muñoz, R. Metabolism of Food Phenolic Acids by Lactobacillus Plantarum CECT 748T. Food Chem. 2008, 107, 1393–1398. [Google Scholar] [CrossRef] [Green Version]
- Kantayos, V.; Kim, J.S.; Baek, S.H. Enhanced Anti-Skin-Aging Activity of Yeast Extract-Treated Resveratrol Rice DJ526. Molecules 2022, 27, 1951. [Google Scholar] [CrossRef]
- Pezet, R.; Perret, C.; Jean-Denis, J.B.; Tabacchi, R.; Gindro, K.; Viret, O. δ-Viniferin, a Resveratrol Dehydrodimer: One of the Major Stilbenes Synthesized by Stressed Grapevine Leaves. J. Agric. Food Chem. 2003, 51, 5488–5492. [Google Scholar] [CrossRef]
- Khamespanah, F.; Patel, N.M.; Forney, A.K.; Heitger, D.R.; Amarasekarage, C.M.; Springer, L.E.; Belecki, K.; Lucas, H.R. Flavonol Dioxygenase Chemistry Mediated by a Synthetic Nickel Superoxide. J. Inorg. Biochem. 2023, 238, 112021. [Google Scholar] [CrossRef] [PubMed]
- López-Fernández-Sobrino, R.; Torres-Fuentes, C.; Bravo, F.I.; Muguerza, B. Winery By-Products as a Valuable Source for Natural Antihypertensive Agents. Crit. Rev. Food Sci. Nutr. 2022, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.J.; Kim, S.R. Beneficial Effects of Flavonoids Against Parkinson’s Disease. J. Med. Food 2018, 21, 421–432. [Google Scholar] [CrossRef]
- Mendonça, C.R.; Manhães-de-Castro, R.; de Santana, B.J.R.C.; Olegário da Silva, L.; Toscano, A.E.; Guzmán-Quevedo, O.; Galindo, L.C.M. Effects of Flavonols on Emotional Behavior and Compounds of the Serotonergic System: A Preclinical Systematic Review. Eur. J. Pharmacol. 2022, 916, 174697. [Google Scholar] [CrossRef] [PubMed]
- Barreca, D.; Trombetta, D.; Smeriglio, A.; Mandalari, G.; Romeo, O.; Felice, M.R.; Gattuso, G.; Nabavi, S.M. Food Flavonols: Nutraceuticals with Complex Health Benefits and Functionalities. Trends Food Sci. Technol. 2021, 117, 194–204. [Google Scholar] [CrossRef]
- Fornairon-Bonnefond, C.; Camarasa, C.; Moutounet, M.; Salmon, J.M. New Trends on Yeast Autolysis and Wine Ageing on Lees: A Bibliographic Review. OENO One 2001, 35, 57–78. [Google Scholar] [CrossRef]
- Ciani, M.; Comitini, F. Yeast Interactions in Multi-Starter Wine Fermentation. Curr. Opin. Food Sci. 2015, 1, 1–6. [Google Scholar] [CrossRef]
- Virdis, C.; Sumby, K.; Bartowsky, E.; Jiranek, V. Lactic Acid Bacteria in Wine: Technological Advances and Evaluation of Their Functional Role. Front. Microbiol. 2021, 11, 3192. [Google Scholar] [CrossRef]
- Devi, A.; Konerira Aiyappaa, A.A.; Waterhouse, A.L. Adsorption and Biotransformation of Anthocyanin Glucosides and Quercetin Glycosides by Oenococcus Oeni and Lactobacillus Plantarum in Model Wine Solution. J. Sci. Food Agricult. 2020, 100, 2110–2120. [Google Scholar] [CrossRef]
- Jara-Palacios, M.J. Wine Lees as a Source of Antioxidant Compounds. Antioxidants 2019, 8, 45. [Google Scholar] [CrossRef] [Green Version]
- Baliyan, S.; Mukherjee, R.; Priyadarshini, A.; Vibhuti, A.; Gupta, A.; Pandey, R.P.; Chang, C.M. Determination of Antioxidants by DPPH Radical Scavenging Activity and Quantitative Phytochemical Analysis of Ficus Religiosa. Molecules 2022, 27, 1326. [Google Scholar] [CrossRef] [PubMed]
- Esparza, I.; Moler, J.A.; Arteta, M.; Jiménez-moreno, N.; Ancín-azpilicueta, C. Phenolic Composition of Grape Stems from Different Spanish Varieties and Vintages. Biomolecules 2021, 11, 1221. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Moreno, N.; Volpe, F.; Moler, J.A.; Esparza, I.; Ancín-Azpilicueta, C. Impact of Extraction Conditions on the Phenolic Composition and Antioxidant Capacity of Grape Stem Extracts. Antioxidants 2019, 8, 597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, N.; Mateus, N.; de Freitas, V.; Oliveira, J. Wine Industry By-Product: Full Polyphenolic Characterization of Grape Stalks. Food Chem. 2018, 268, 110–117. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Compound | Rt * (min) | m/z [M − H] | m/z MS2 [M − H]− | m/z MS3 [M − H]− | Ref. | Grape Stem | Grape Pomace | Wine Lees |
|---|---|---|---|---|---|---|---|---|---|
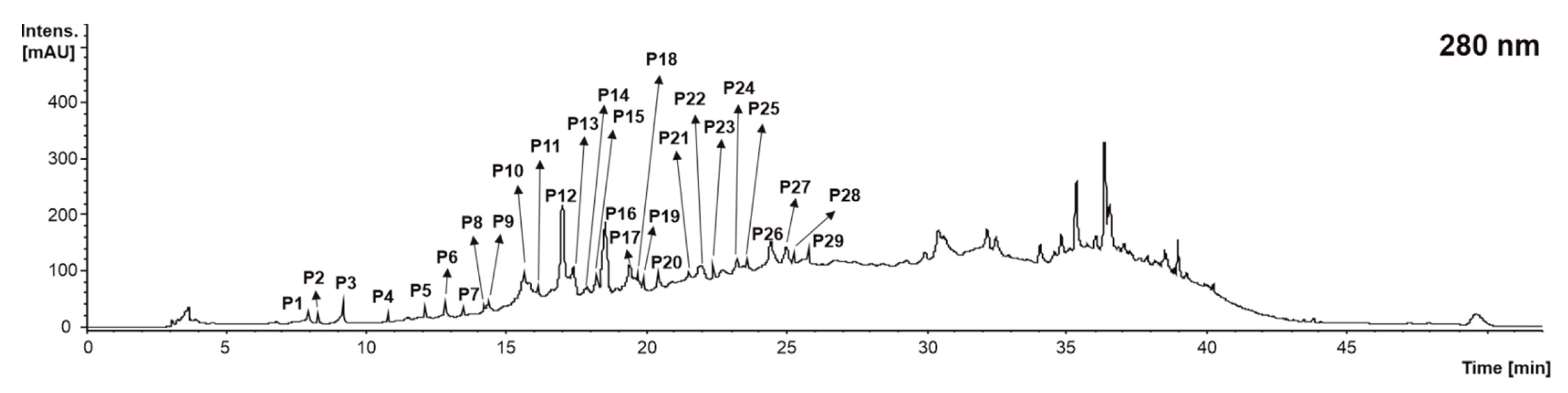
| P1 | Proanthocyanidin trimer (B type) isomer 1 | 7.9 | 865 | 695(100), 405(36), 287(29), 577(27) | 695: 543(100), 391(46), 256(40) | [23,24] | X | X | <LOD |
| P2 | Proanthocyanidin trimer (B type) isomer 2 | 8.4 | 865 | 695(100), 405(36), 287(29), 577(27) | 695: 543(100), 391(46), 256(40) | [23,24] | X | X | X |
| P3 | Catechin-Gallocatechin isomer 1 | 8.8 | 593 | 441(100), 423(15), 407(12) | 441: 315(100), 289(52), 153(27), 191(25) | [23,25] | X | X | X |
| P4 | Catechin-Gallocatechin isomer 2 | 10.7 | 593 | 441(100), 423(15), 407(12) | 441: 315(100), 289(52), 153(27), 191(25) | [23,25] | X | X | X |
| P5 | Gallocatechin | 12.0 | 305 | 179(100), 219(75), 260(30), | 179: 167(100), 151(46), 137(11) | [25] | <LOD | <LOD | X |
| P6 | Catechin-Gallocatechin isomer 3 | 12.5 | 593 | 423(100), 305(43), 441(23) | 423: 283(100), 297(83), 255(25) | [23,25] | <LOD | X | <LOD |
| P7 | Catechin-Gallocatechin isomer 4 | 13.3 | 593 | 423(100), 305(38), 441(25) | 423: 283(100), 297(60), 255(22) | [23,25] | <LOD | <LOD | X |
| P8 | Procyanthocyanidin dimer (B type) isomer 1 | 14.3 | 577 | 425(100), 407(97), 289(29), 451(16) | 425: 407(100), 272(10) | [23,24] | X | X | <LOD |
| P9 | Procyanthocyanidin dimer (B type) isomer 2 | 14.9 | 577 | 425(100), 407(90), 289(33), 451(23), 559(17) | 425: 407(100), 272(10) | [23,24] | X | X | <LOD |
| P10 | Procyanthocyanidin dimer (B type) isomer 3 | 15.5 | 577 | 425(100), 407(97), 289(29), 451(16) | 425: 407(100), 272(10) | [23,24] | X | X | <LOD |
| P11 | Procyanthocyanidin dimer (B type) isomer 4 | 16.1 | 577 | 407(100), 287(62), 425(54) | 425: 285(100), 281(59) | [23,24] | X | X | X |
| P12 | Procyanthocyanidin dimer (B type) isomer 5 | 17.3 | 577 | 425(100), 407(71), 289(45), 451(26) | 425: 407(100), 272(6) | [23,24] | X | X | X |
| P13 | Proanthocyanidin trimer (B type) isomer 3 | 17.5 | 865 | 695(100), 407(43), 287(22), 577(53) | 695: 543(100), 242(80), 525(66), 405(57) | [23,24] | X | X | X |
| P14 | Proanthocyanidin dimer di-gallate (B type) | 17.7 | 881 | 695(100), 289(16), 443(16) | 695: 543(100), 289(32), 242(24) | [23,25] | <LOD | X | X |
| P15 | Proanthocyanidin trimer (B type) isomer 4 | 17.9 | 865 | 695(100), 577(75), 407(63), 303(70), 287(56) | 695: 405(100), 242(99), 543(87), 677(70), 525(75) | [23,24] | X | <LOD | <LOD |
| P16 | Catechin | 18.5 | 289 | 245(100), 205(31) | 245: 202(100), 226(27), 187(22), 161(15) | [24] | X | X | X |
| P17 | Proanthocyanidin trimer (B type) isomer 5 | 19.2 | 865 | 407(100), 577(75), 407(63), 287(56) | 695: 405(100), 695(83), 577(62), 287(53) | [23,24] | X | X | <LOD |
| P18 | Proanthocyanidin trimer (B type) isomer 6 | 19.6 | 865 | 407(100), 577(75), 407(63), 287(56) | 695: 405(100), 695(83), 577(62), 287(53) | [23,24] | X | X | X |
| P19 | Proanthocyanidin trimer monogallate isomer 1 | 20.0 | 1017 | 729(100), 677(36), 577(34), 407(19) | 729: 577(100), 407(56), 451(29), 425(27) | [26,27] | X | <LOD | <LOD |
| P20 | Proanthocyanidin trimer monogallate isomer 2 | 20.6 | 1017 | 729(100), 677(36), 577(34), 407(19) | 729: 577(100), 407(56), 451(29), 425(27) | [26,27] | X | X | X |
| P21 | Procyanthocyanidin dimer (B type) isomer 6 | 21.2 | 577 | 425(100), 407(83), 289(37), 451(23) | 425: 407(100), 272(7) | [23,24] | X | <LOD | X |
| P22 | Procyanthocyanidin dimer (B type) isomer 7 | 21.6 | 577 | 425(100), 407(83), 289(37), 451(23) | 425: 407(100), 272(7) | [23,24] | X | X | X |
| P23 | Proanthocyanidin dimer monogallate isomer 1 | 22.3 | 729 | 577(100), 407(83), 559(62), 425(54), 451(21) | 577: 407(100), 451(70), 289(29), 425(35) | [27,28] | X | <LOD | <LOD |
| P24 | Proanthocyanidin dimer monogallate isomer 2 | 23.1 | 729 | 407(100), 577(36), 559(31), 451(7) | 407: 285(100), 257(38), 297(33), 243 (28) | [27,28] | X | X | <LOD |
| P25 | Proanthocyanidin trimer (B type) isomer 7 | 23.5 | 865 | 577(100), 407(80), 695(74), 451(42), 287(42) | 695: 242(100), 543(90), 407(623), 451(62), 525(57), 289(28) | [23,24] | X | X | X |
| P26 | Epicatechin-glucoside | 24.2 | 449 | 287(100), 269(60) | 287: 259(100), 242(10) | [25] | X | <LOD | <LOD |
| P27 | Epicatechin | 24.8 | 289 | 245(100), 205(29) | 245: 202(100), 226(29), 187(23), 161(15) | [24] | X | X | X |
| P28 | Proanthocyanidin dimer monogallate isomer 3 | 25.2 | 729 | 407(100), 559(53), 441(39), 577(37), 451(29), 289(29) | 577: 407(100), 451(70), 289(29), 425(35) | [27,28] | X | X | X |
| P29 | Procyanthocyanidin dimer (B type) isomer 8 | 25.9 | 577 | 425(100), 407(76), 289(32), 451(32) | 425: 407(100), 272(8) | [23,24] | X | <LOD | X |
| Peak | Compound | Rt * (min) | m/z [M − H] | m/z MS2 [M − H] | m/z MS3 [M − H] | Ref. | Grape Stem | Grape Pomace | Wine Lees |
|---|---|---|---|---|---|---|---|---|---|
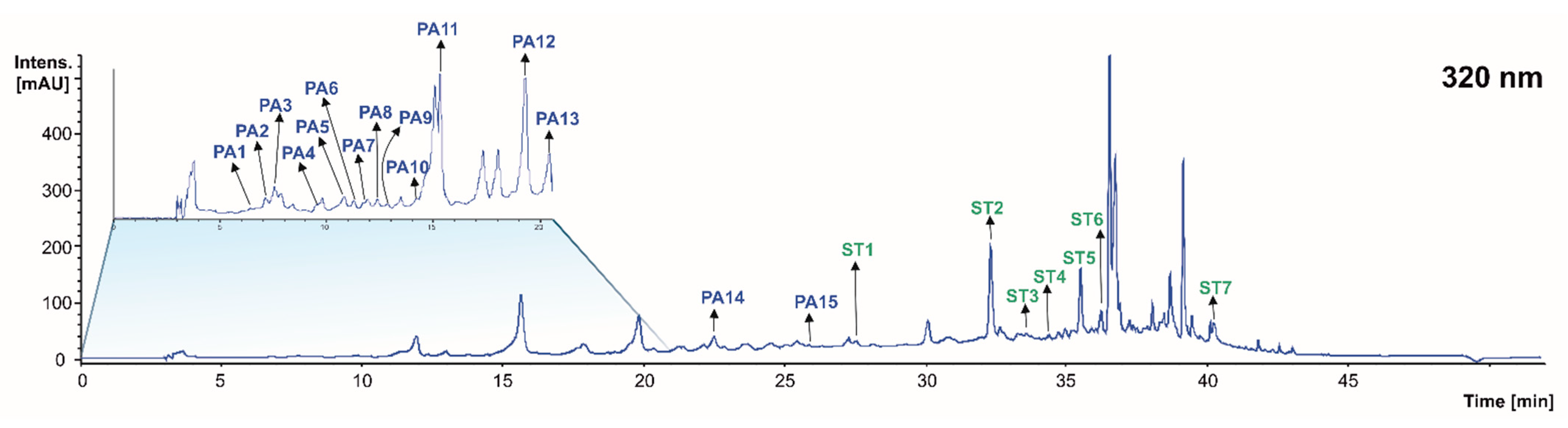
| PA1 | Galloyl-hexoside isomer 1 | 6.2 | 331 | 169(100), 271(40), 193(39), 151(269 | 169: 125(100), 151(20) | [24,27] | X | X | <LOD |
| PA2 | Gallic acid isomer 1 | 7.0 | 169 | 125(100) | N.d. | [28,30] | X | X | X |
| PA3 | Gallic acid isomer 2 | 7.5 | 169 | 125(100) | N.d. | [28,30] | X | X | X |
| PA4 | Gentisic acid | 9.7 | 153 | 153(100) | 153: 123(100), 109(34) | [30] | X | <LOD | <LOD |
| PA5 | Protocatecuic acid-O-hexoside isomer 1 | 10.4 | 315 | 153(100), 165(37), 108(19) | 153: 108(100), 123(35) | [30] | <LOD | X | <LOD |
| PA6 | Protocatecuic acid-O-hexoside isomer 2 | 11.2 | 315 | 153(100), 108(33), 165(15) | 153: 108(100), 123(35) | [30] | X | X | <LOD |
| PA7 | Galloyl-hexoside isomer 2 | 11.7 | 331 | 169(100), 125(7) | 169: 125(100) | [24,27] | X | <LOD | <LOD |
| PA8 | Protocatecuic acid-O-hexoside isomer 3 | 12.3 | 315 | 153(100), 165(24), 108(10) | 153: 123(100) | [30] | X | <LOD | <LOD |
| PA9 | Caftaric acid | 12.8 | 311 | 149(100), 179(38), 135(8) | 149: 103(100), 131(82) | [30] | <LOD | <LOD | X |
| PA10 | Caftaric acid-glucuronide | 14.1 | 487 | 355(100), 311(65), 167(42), 211(13) | 355: 167(100), 311(79), 211(26) | [30] | <LOD | <LOD | X |
| PA11 | Caftaric acid derivative | 15.4 | 623 | 311(100), 179(7) | 311: 149(100), 179(51) | [30] | <LOD | <LOD | X |
| PA12 | p-coumaric acid pentoside | 19.1 | 295 | 163(100), 119(8) | 163: 119(100) | [30] | <LOD | <LOD | X |
| PA13 | p-coumaric acid | 20.4 | 162 | 119(100) | N.d. | [30] | <LOD | <LOD | X |
| PA14 | Ferulic acid pentoside | 22.1 | 325 | 193(100), 148(4) | 193: 178(100), 134(84), 148(75) | [30] | <LOD | <LOD | X |
| PA15 | Ethyl gallate | 26.1 | 197 | 169(100), 125(30) | 169: 125(100) | [31,32] | X | <LOD | X |
| Peak | Compound | Rt * (min) | m/z [M − H] | m/z MS2 [M − H] | m/z MS3 [M − H] | Ref. | Grape Stem | Grape Pomace | Wine Lees |
|---|---|---|---|---|---|---|---|---|---|
| St1 | Oxyresveratrol-glucoside | 27.4 | 405 | 243(100) | 243: 224(100), 198(396), 174(15) | [35] | <LOD | <LOD | X |
| St2 | Trans-Piceid isomer 1 | 32.6 | 389 | 227 (100) | 227: 184(100), 156(37) | [33,36] | X | <LOD | <LOD |
| St3 | Oxyresveratrol | 33.5 | 243 | 224(100), 198(80), 154(69) | 224: 180(100), 137(39), 163(14) | [35] | <LOD | <LOD | X |
| St4 | Stilbenoid tetramer (Hopeaphenol) | 34.4 | 905 | 717(100), 811(94), 359(20), 451(18) | 717: 675(100), 611(66), 357(53) | [37] | X | X | <LOD |
| St5 | Σ-viniferin isomer 1 | 35.1 | 453 | 359(100), 227(45), 265(23) | 359: 265(100) | [38] | X | X | X |
| St6 | Trans-piceid isomer 2 | 36.2 | 389 | 227(100) | 227: 184(100), 156(37) | [33,36] | <LOD | <LOD | X |
| St7 | E-viniferin isomer 2 | 40.2 | 453 | 359(100), 227(45), 265(26) | 359: 265(100) | [38] | X | X | X |
| Peak | Compound | Rt * (min) | m/z [M − H] | m/z MS2 [M − H] | m/z MS3 [M − H] | Ref. | Grape Stem | Grape Pomace | Wine Lees |
|---|---|---|---|---|---|---|---|---|---|
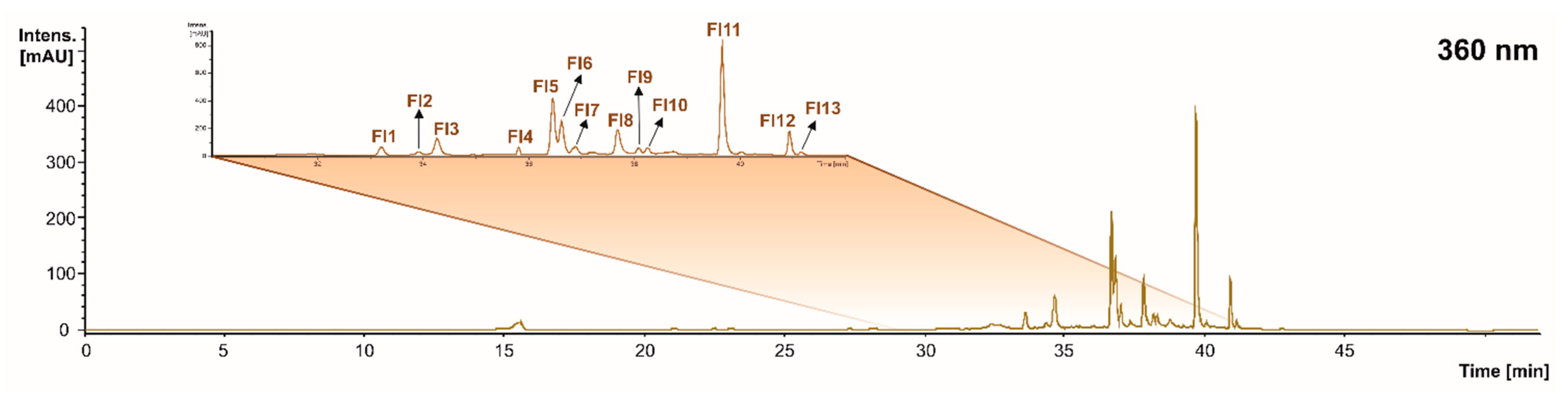
| Fl1 | 3’,5’-di-methyltricetin derivative isomer 1 | 33.2 | 509 | 329(100), 347(61), 355(32), 193(12) | 329: 313(100), 148(4) | [40] | X | <LOD | X |
| Fl2 | 3’,5’-di-methyltricetin derivative isomer 2 | 33.9 | 509 | 329(100), 347(61), 355(32), 193(12) | 329: 313(100), 148(4) | [40] | <LOD | <LOD | X |
| Fl3 | Myricetin hexoside | 34.4 | 479 | 317(100), 271(9), 179(4) | 317: 271(100), 287(36), 179(22), 151(16) | [41] | X | X | X |
| Fl4 | Kaempferol glucoside isomer 1 | 35.6 | 447 | 285(100), 303(53), 151(15), 179(6) | 285: 241(100), 175(47) | [38] | X | X | <LOD |
| Fl5 | Quercetin 3-glucuronide | 36.8 | 477 | 301(100), 151(2) | 301: 151(100), 179(93), 257(34), 272(18) | [38] | X | X | X |
| Fl6 | Quercetin 3-glucoside isomer 1 | 36.6 | 463 | 301(100), 271(7), 343(4), 151(4) | 301: 179(100), 151(93), 271(38), 255(34) | [33] | X | X | X |
| Fl7 | Quercetin 3-glucoside isomer 2 | 36.9 | 463 | 301(100), 271(7), 343(4), 151(4) | 301: 179(100), 151(93), 271(38), 255(34) | [33] | X | <LOD | X |
| Fl8 | Myricetin | 37.6 | 317 | 179(100), 151(60), 193(21) | 179: 150(100), 169(16) | [42] | X | <LOD | X |
| Fl9 | Kaempferol glucoside isomer 2 | 38.2 | 447 | 285(100), 255(53), 227(12), 169(4) | 284: 255(100), 227(16), 163(3) | [33] | X | X | X |
| Fl10 | Isorhamnetin hexoside | 38.4 | 477 | 315(100), 28520), 270(17), 357(15) | 315: 285(100), 270(71), 299(53), 242(23) | [41] | X | <LOD | <LOD |
| Fl11 | Quercetin | 39.7 | 301 | 179(100), 151(84), 272(25) | 179: 151(100), 169(4), 107(4) | [33] | X | <LOD | X |
| Fl12 | Kaempferol | 40.9 | 285 | 214(100), 153(67), 185(67), 165(52), 257(46) | N.d. | [43] | X | <LOD | X |
| Fl13 | Isorhamnetin | 41.2 | 315 | 301(100) | 301: 151(100), 271(70), 227(38), 192(26), 282(27), 164(12) | [44] | <LOD | <LOD | X |
| Peak | Compound | Rt* (min) | m/z [M + H] | m/z MS2 [M + H] | m/z MS3 [M + H] | Ref. | Grape Stem | Grape Pomace | Wine Lees |
|---|---|---|---|---|---|---|---|---|---|
| An1 | Delphinidin 3-glucoside | 26.3 | 465 | 303(100) | 303: 257(100), 229(37), 247(114), 179(13), 275(12) | [45] | X | X | X |
| An2 | Cyanidin 3-glucoside | 28.6 | 449 | 287(100) | 287: 193(100), 231(34), 270(30), 109(27), 137(22), 203(16) | [45] | X | X | X |
| An3 | Petunidin 3-glucoside | 30.6 | 479 | 317(100), 302(1) | 317: 302(100), 274(39) | [45] | X | X | X |
| An4 | Peonidin 3-glucoside | 32.9 | 463 | 301(100) | 301: 286(100), 257(4), 241(2), 230(2) | [45] | X | X | X |
| An5 | Malvidin 3-glucoside | 33.1 | 493 | 331(100), 316(1) | 331: 299(100), 315(52), 270(21), 242(17), 179(12) | [45] | X | X | X |
| An6 | Delphinidin 3-acetylglucoside | 35.4 | 507 | 303(100) | 303: 257(100), 246(7) | [45] | <LOD | <LOD | X |
| An7 | Petunidin 3-acetylglucoside | 36.1 | 521 | 317(100) | 317: 302(100), 274(12) | [45] | <LOD | X | X |
| An8 | Peonidin 3-acetylglucoside | 37.0 | 505 | 301(100), 286(8) | 301: 286(100), 211(5) | [45] | X | X | X |
| An9 | Malvidin 3-acetylglucoside | 37.2 | 535 | 331(100), 343(6) | 331: 315(100), 299(99), 242(49), 270(38), 179(25), 139(9) | [45] | X | X | X |
| An10 | Delphinidin 3-p-coumaroylglucoside | 37.8 | 611 | 303(100) | 303: 257(100), 275(5) | [45] | X | X | X |
| An11 | Malvidin 3-6-caffeoyl-glucoside | 37.9 | 655 | 331(100), 315(3), 242(3) | 331: 315(100), 299(99), 270(75), 287(66), 179(38), 242(30) | [45] | X | X | X |
| An12 | Cyanidin 3-p-coumaroylglucoside | 38.4 | 595 | 287(100), 173(3), 185(3) | 287: 213(100), 157(51), 231(51), 269(32), 185(20), 137(12) | [45] | X | X | X |
| An13 | Petunidin 3- coumaroylglucoside | 38.5 | 625 | 317(100), 302(4), 274(2) | 317: 302(100), 274(18), 228(6) | [45] | X | X | X |
| An14 | Peonidin 3-(6-trans-p-coumaroyl)-glucoside | 40.2 | 609 | 301(100), 286(16), 201(3) | 301: 286(100) | [45] | X | X | X |
| An15 | Malvidin 3-p-coumaroylglucoside | 40.7 | 639 | 331(100), 269(5), 315(4), 241(1) | 331: 299(100), 315(98), 270(77), 242(37), 179(18), 253(6), 150(5) | [45] | X | X | X |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa-Pérez, A.; Medina, S.; Sánchez-Bravo, P.; Domínguez-Perles, R.; García-Viguera, C. The (Poly)phenolic Profile of Separate Winery By-Products Reveals Potential Antioxidant Synergies. Molecules 2023, 28, 2081. https://doi.org/10.3390/molecules28052081
Costa-Pérez A, Medina S, Sánchez-Bravo P, Domínguez-Perles R, García-Viguera C. The (Poly)phenolic Profile of Separate Winery By-Products Reveals Potential Antioxidant Synergies. Molecules. 2023; 28(5):2081. https://doi.org/10.3390/molecules28052081
Chicago/Turabian StyleCosta-Pérez, Antonio, Sonia Medina, Paola Sánchez-Bravo, Raúl Domínguez-Perles, and Cristina García-Viguera. 2023. "The (Poly)phenolic Profile of Separate Winery By-Products Reveals Potential Antioxidant Synergies" Molecules 28, no. 5: 2081. https://doi.org/10.3390/molecules28052081