
Acenocoumarol Exerts Anti-Inflammatory Activity via the Suppression of NF-κB and MAPK Pathways in RAW 264.7 Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
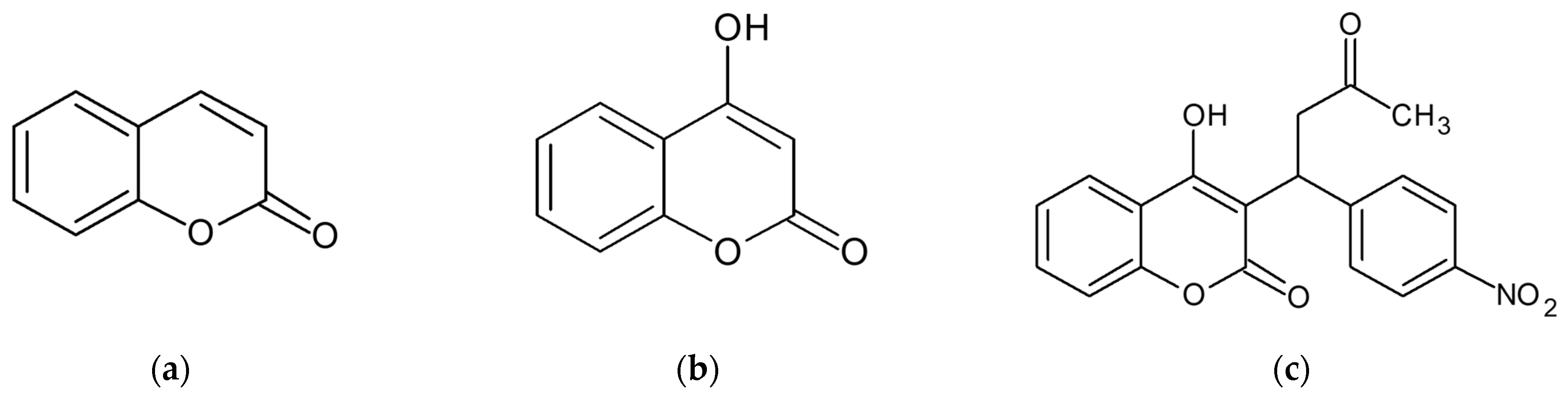
:1. Introduction
2. Results
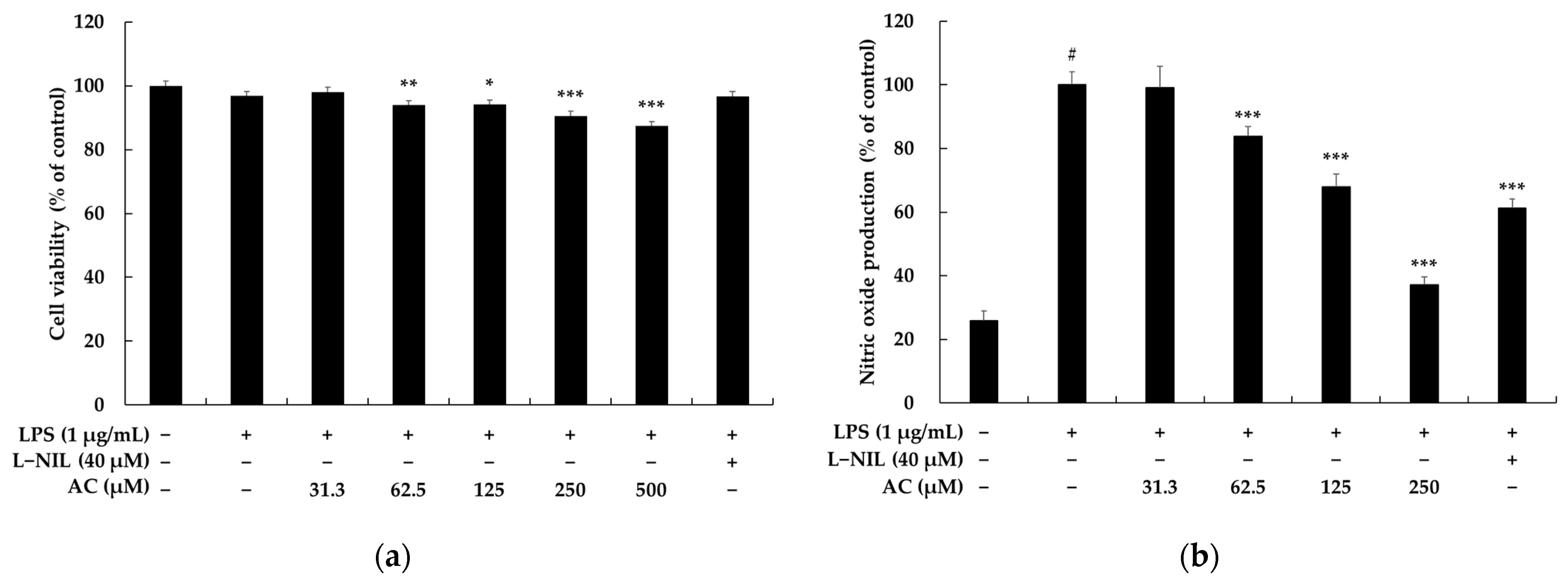
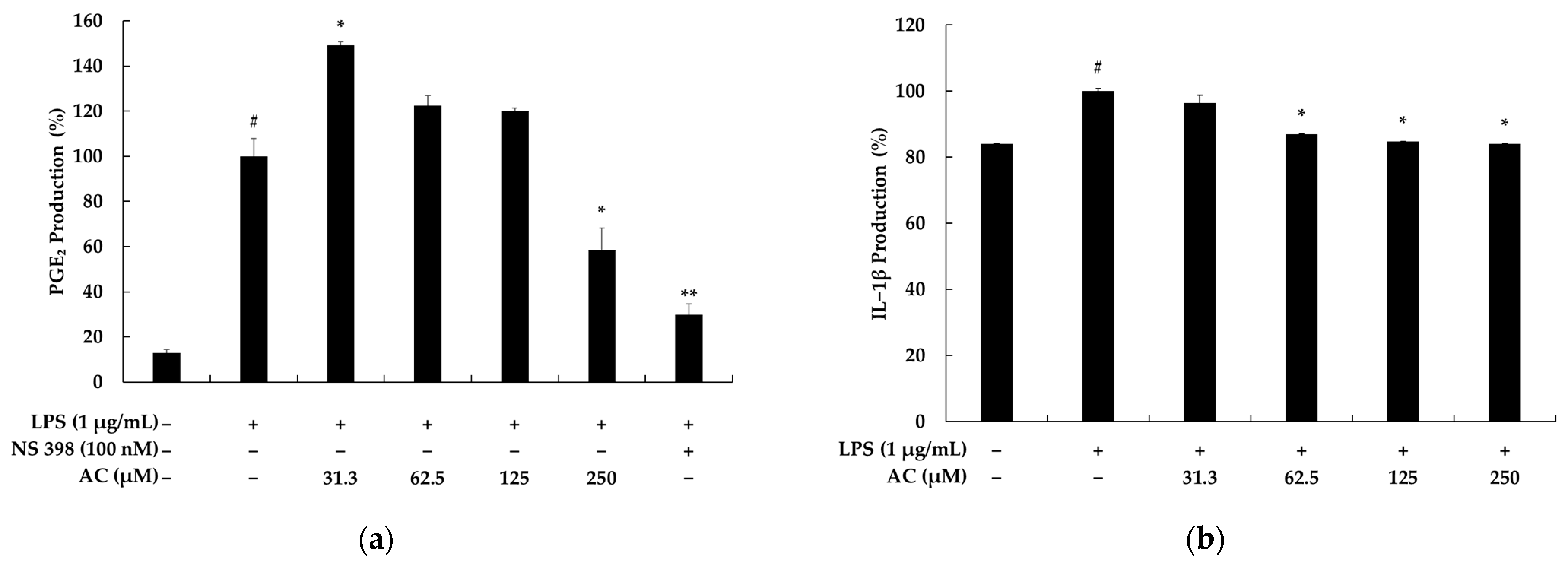
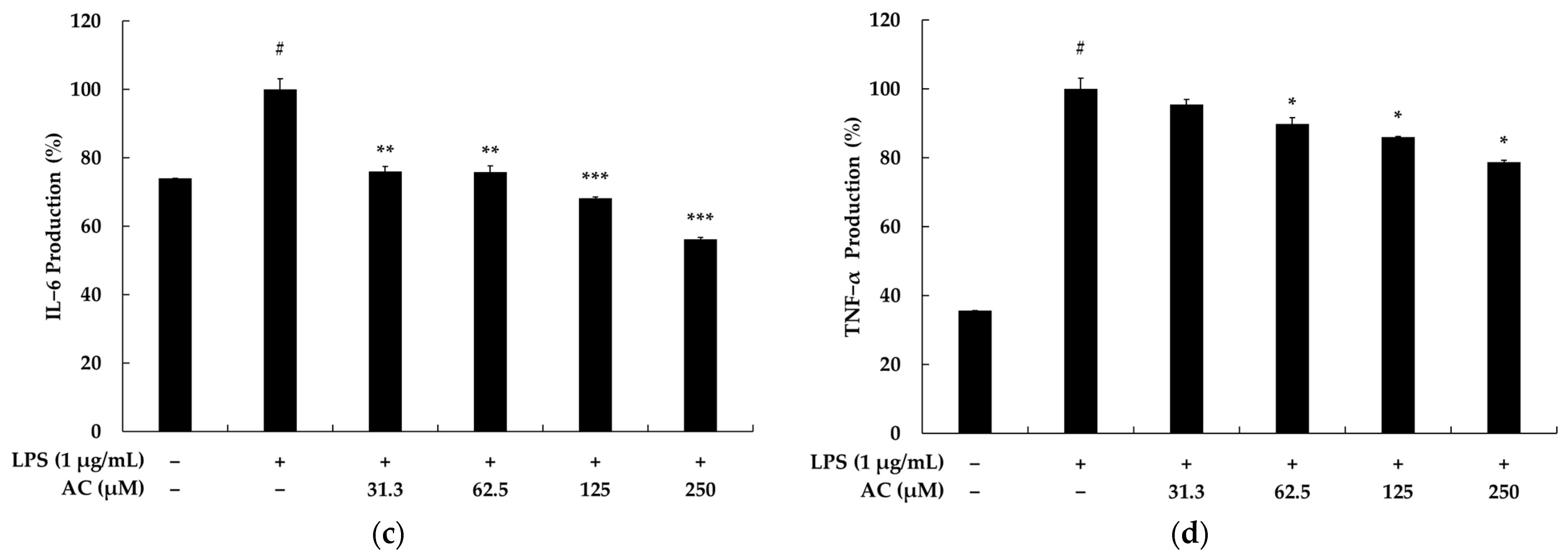
2.1. Effects of Acenocoumarol on Viability, Pro-Inflammatory Mediators, and Cytokines of RAW 264.7 Cells
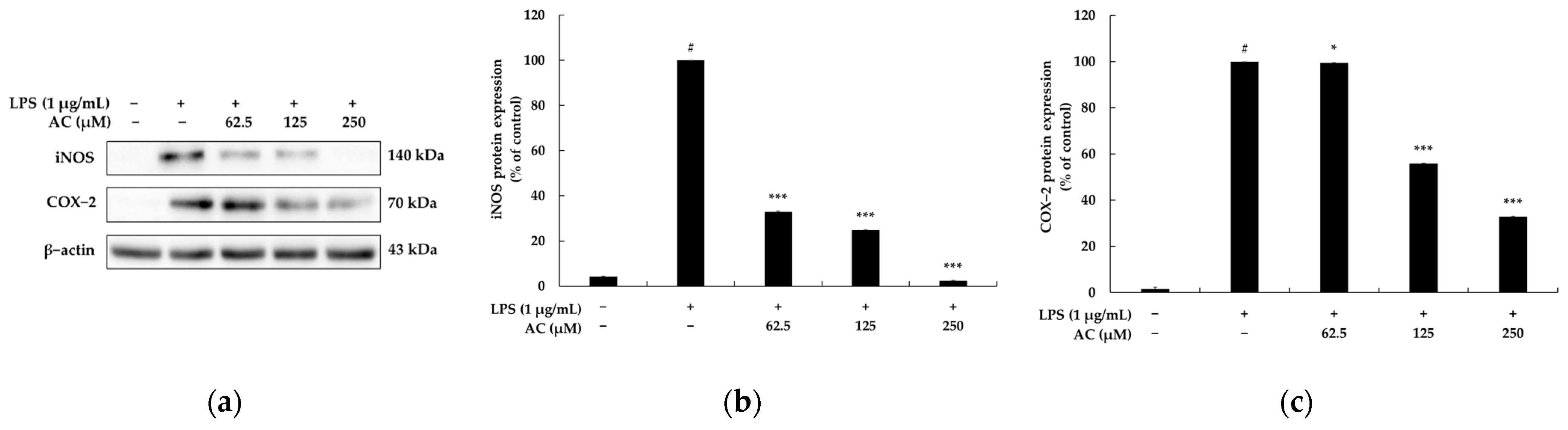
2.2. Effect of Acenocoumarol on NO Synthase (iNOS) and Cyclooxygenase (COX)-2 Production
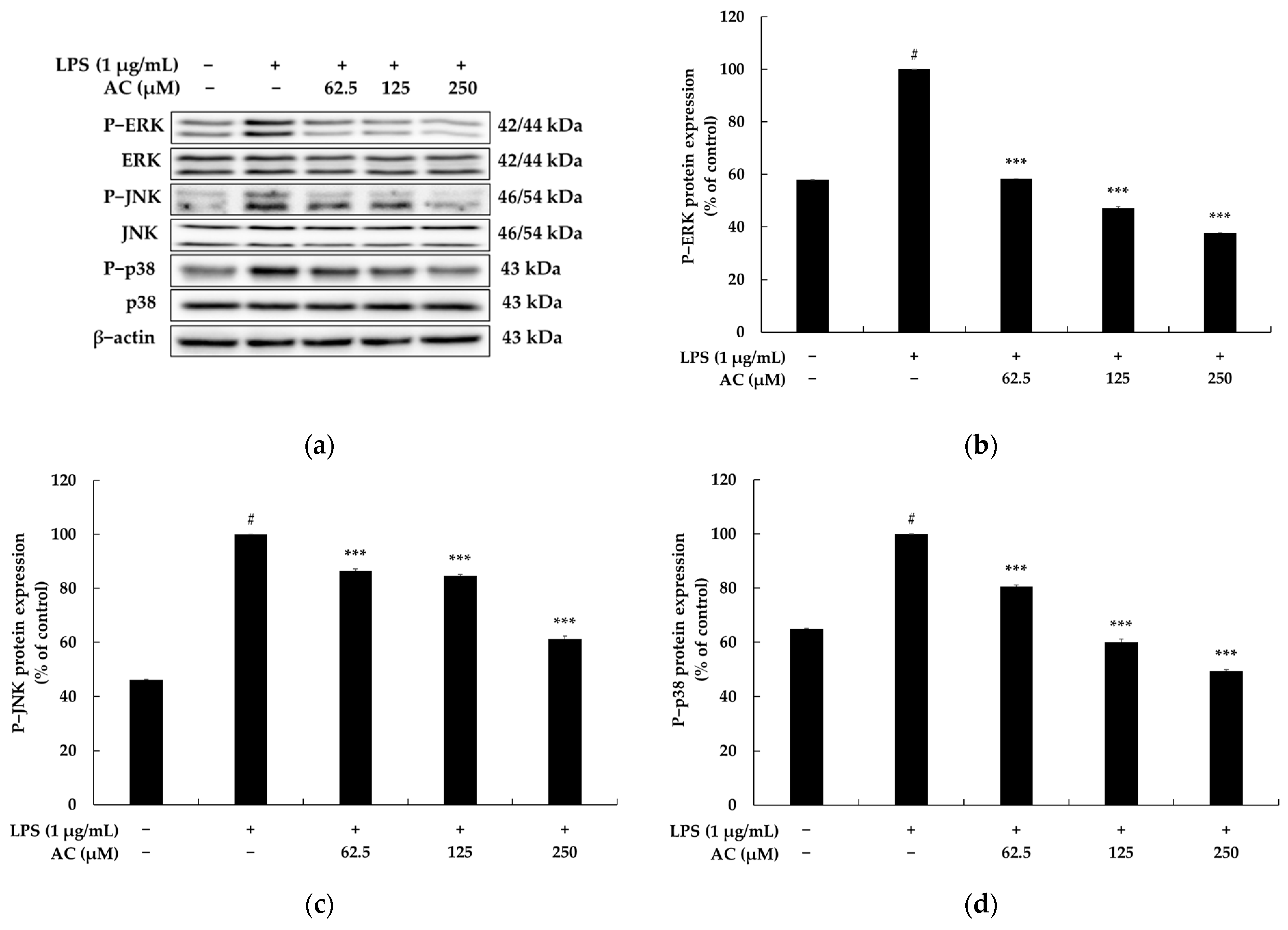
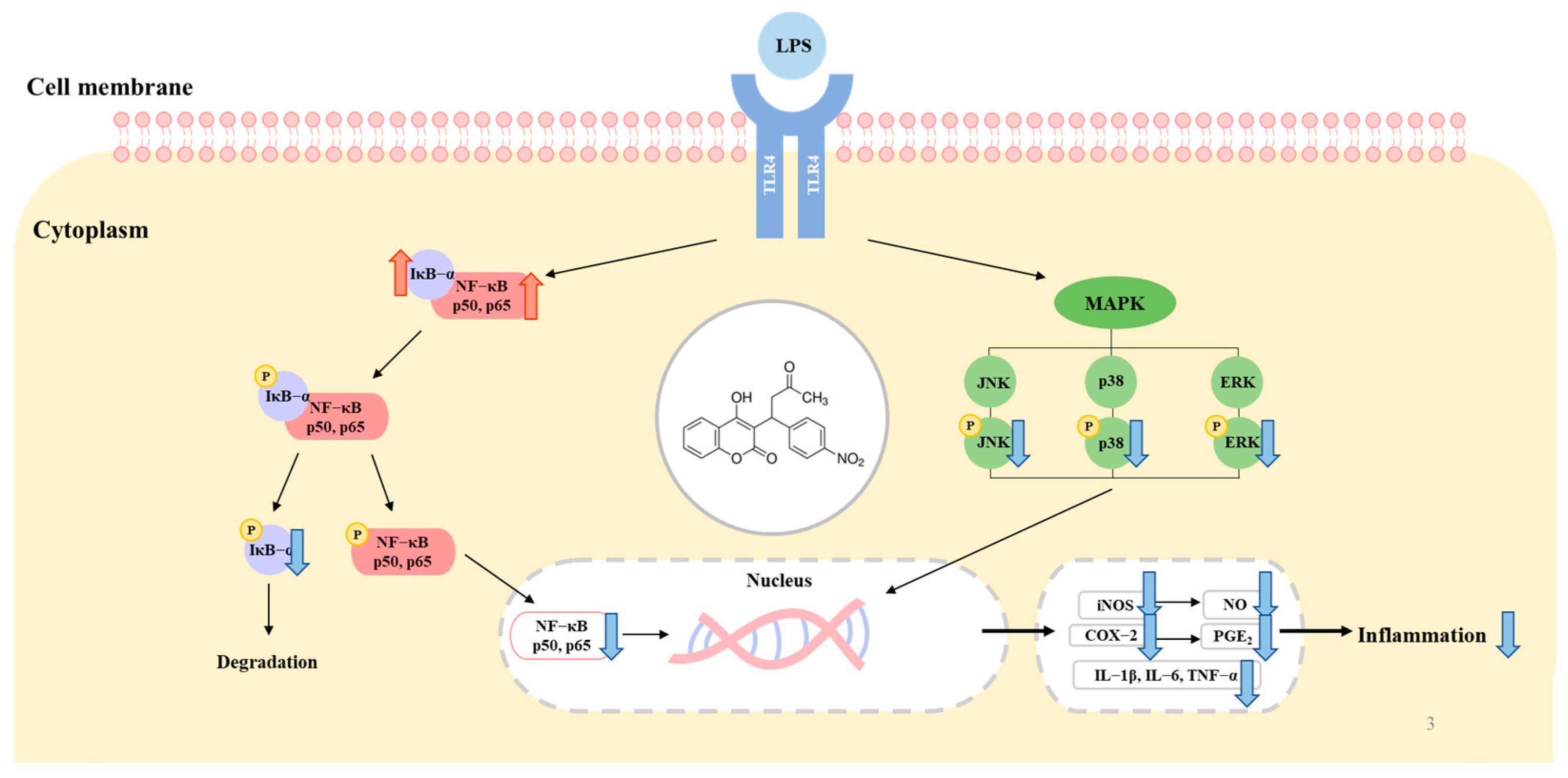
2.3. Effect of Acenocoumarol on the Mitogen-Activated Protein Kinase (MAPK) Signaling Pathway
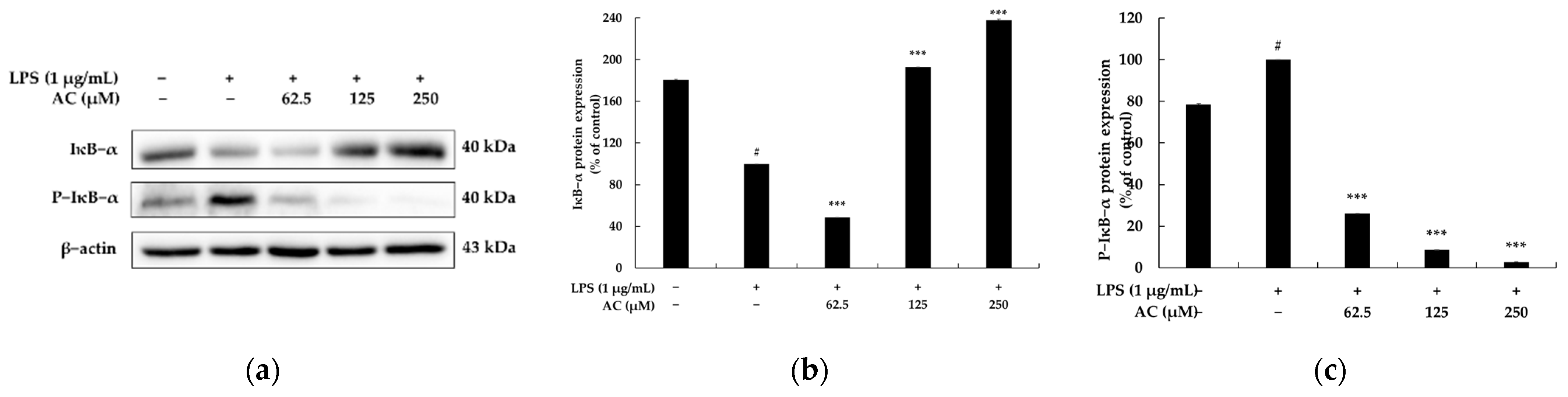
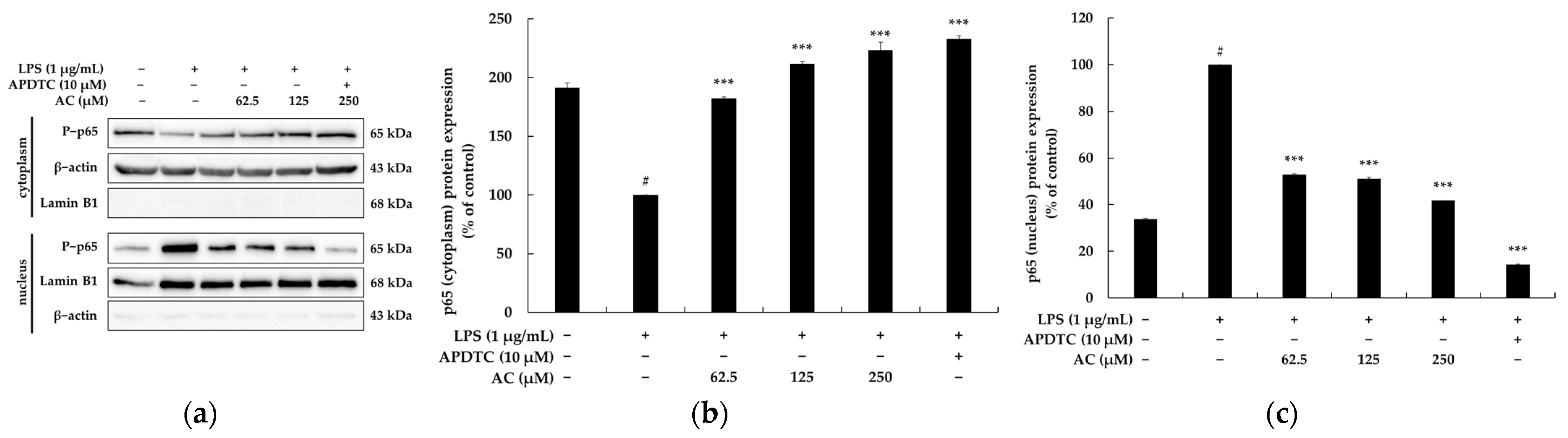
2.4. Effect of Acenocoumarol on the Nuclear Factor κB (NF-κB) Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. MTT Assay
4.4. Measurement of NO Production
4.5. Measurement of PGE2 and Cytokines
4.6. Preparation of Nuclear and Cytoplasmic Extraction
4.7. Western Blotting
4.8. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Takahashi, S.; Yoshida, M.; Watanabe, M.M.; Isoda, H. Anti-Inflammatory Effects of Aurantiochytrium limacinum 4W-1b Ethanol Extract on Murine Macrophage RAW264 Cells. Biomed. Res. Int. 2019, 2019, 3104057. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef] [Green Version]
- Henein, M.Y.; Vancheri, S.; Longo, G.; Vancheri, F. The Role of Inflammation in Cardiovascular Disease. Int. J. Mol. Sci. 2022, 23, 12906. [Google Scholar] [CrossRef]
- Li, H.; Meng, Y.; He, S.; Tan, X.; Zhang, Y.; Zhang, X.; Wang, L.; Zheng, W. Macrophages, Chronic Inflammation, and Insulin Resistance. Cells 2022, 11, 3001. [Google Scholar] [CrossRef]
- Nowowiejska, J.; Baran, A.; Flisiak, I. Psoriasis and neurodegenerative diseases—A review. Front. Mol. Neurosci. 2022, 15, 917751. [Google Scholar] [CrossRef]
- Yang, L.; He, J. Anti-inflammatory effects of flavonoids and phenylethanoid glycosides from Hosta plantaginea flowers in LPS-stimulated RAW 264.7 macrophages through inhibition of the NF-κB signaling pathway. BMC Complement. Med. Ther. 2022, 22, 55. [Google Scholar] [CrossRef]
- Facchin, B.M.; Dos Reis, G.O.; Vieira, G.N.; Mohr, E.T.B.; da Rosa, J.S.; Kretzer, I.F.; Demarchi, I.G.; Dalmarco, E.M. Inflammatory biomarkers on an LPS-induced RAW 264.7 cell model: A systematic review and meta-analysis. Inflamm. Res. 2022, 71, 741–758. [Google Scholar] [CrossRef]
- Qu, L.; Matz, A.J.; Karlinsey, K.; Cao, Z.; Vella, A.T.; Zhou, B. Macrophages at the Crossroad of Meta-Inflammation and Inflammaging. Genes 2022, 13, 2074. [Google Scholar] [CrossRef]
- Hartley, J.W.; Evans, L.H.; Green, K.Y.; Naghashfar, Z.; Macias, A.R.; Zerfas, P.M.; Ward, J.M. Expression of infectious murine leukemia viruses by RAW264.7 cells, a potential complication for studies with a widely used mouse macrophage cell line. Retrovirology 2008, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Ghelani, H.; Khursheed, M.; Adrian, T.E.; Jan, R.K. Anti-Inflammatory Effects of Compounds from Echinoderms. Mar. Drugs 2022, 20, 693. [Google Scholar] [CrossRef]
- Zhao, H.; Wu, L.; Yan, G.; Chen, Y.; Zhou, M.; Wu, Y.; Li, Y. Inflammation and tumor progression: Signaling pathways and targeted intervention. Signal Transduct. Target. Ther. 2021, 6, 263. [Google Scholar] [CrossRef]
- Yeung, Y.T.; Aziz, F.; Guerrero-Castilla, A.; Arguelles, S. Signaling Pathways in Inflammation and Anti-inflammatory Therapies. Curr. Pharm. Des. 2018, 24, 1449–1484. [Google Scholar] [CrossRef]
- Chaffey, L.; Roberti, A.; Greaves, D.R. Drug repurposing in cardiovascular inflammation: Successes, failures, and future opportunities. Front. Pharmacol. 2022, 13, 1046406. [Google Scholar] [CrossRef]
- Paul, A.; Kumar, M.; Das, P.; Guha, N.; Rudrapal, M.; Zaman, M.K. Drug repurposing—A search for novel therapy for the treatment of diabetic neuropathy. Biomed. Pharmacother. 2022, 156, 113846. [Google Scholar] [CrossRef]
- Kuang, Y.; Xia, Y.; Wang, X.; Rao, Q.; Yang, S. Magnetic Surface Molecularly Imprinted Polymer for Selective Adsorption of 4-Hydroxycoumarin. Front. Chem. 2022, 10, 862777. [Google Scholar] [CrossRef]
- Stanchev, S.; Jensen, F.; Hinkov, A.; Atanasov, V.; Genova-Kalou, P.; Argirova, R.; Manolov, I. Synthesis and Inhibiting Activity of Some 4-Hydroxycoumarin Derivatives on HIV-1 Protease. ISRN Pharm. 2011, 2011, 137637. [Google Scholar] [CrossRef]
- Li, M.K.; Li, J.; Liu, B.H.; Zhou, Y.; Li, X.; Xue, X.Y.; Hou, Z.; Luo, X.X. Synthesis, crystal structures, and anti-drug-resistant Staphylococcus aureus activities of novel 4-hydroxycoumarin derivatives. Eur. J. Pharmacol. 2013, 721, 151–157. [Google Scholar] [CrossRef]
- Basak, V.; Bahar, T.E.; Emine, K.; Yelda, K.; Mine, K.; Figen, S.; Rustem, N. Evaluation of Cytotoxicity and Gelatinases Activity in 3T3 Fibroblast Cell by Root Repair Materials. Biotechnol. Biotechnol. Equip. 2016, 30, 984–990. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Hyun, C.G. Anti-Inflammatory Effects of Psoralen Derivatives on RAW264.7 Cells via Regulation of the NF-κB and MAPK Signaling Pathways. Int. J. Mol. Sci. 2022, 23, 5813. [Google Scholar] [CrossRef]
- Kang, J.K.; Chung, Y.C.; Hyun, C.G. Anti-Inflammatory Effects of 6-Methylcoumarin in LPS-Stimulated RAW 264.7 Macrophages via Regulation of MAPK and NF-κB Signaling Pathways. Molecules 2021, 26, 5351. [Google Scholar] [CrossRef]
- Ullah, S.; Chung, Y.C.; Hyun, C.G. Induction of Melanogenesis by Fosfomycin in B16F10 Cells Through the Upregulation of P-JNK and P-p38 Signaling Pathways. Antibiotics 2020, 9, 172. [Google Scholar] [CrossRef]
- Moon, S.H.; Chung, Y.C.; Hyun, C.G. Tobramycin Promotes Melanogenesis by Upregulating p38 MAPK Protein Phosphorylation in B16F10 Melanoma Cells. Antibiotics 2019, 8, 140. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.S.; Chung, Y.C.; Moon, S.H.; Hyun, C.G. Lincomycin induces melanogenesis through the activation of MITF via p38 MAPK, AKT, and PKA signaling pathways. J. Appl. Biol. Chem. 2021, 64, 323–331. [Google Scholar] [CrossRef]
- Kang, J.K.; Kang, H.K.; Hyun, C.G. Anti-Inflammatory Effects of Spiramycin in LPS-Activated RAW 264.7 Macrophages. Molecules 2022, 27, 3202. [Google Scholar] [CrossRef]
- Kang, H.K.; Hyun, C.G. Anti-inflammatory Effect of d-(+)-Cycloserine Through Inhibition of NF-κB and MAPK Signaling Pathways in LPS-Induced RAW 264.7 Macrophages. Nat. Prod. Commun. 2020, 15, 1934578X20920481. [Google Scholar] [CrossRef]
- Schroecksnadel, S.; Gostner, J.; Jenny, M.; Kurz, K.; Schennach, H.; Weiss, G.; Fuchs, D. Immunomodulatory effects in vitro of vitamin K antagonist acenocoumarol. Thromb. Res. 2013, 131, e264–e269. [Google Scholar] [CrossRef]
- Salvador, J.P.; Brettschneider, T.; Dorrer, C.; Marco, M.P. Development of a Fluorescent Microfluidic Device Based on Antibody Microarray Read-Out for Therapeutic Drug Monitoring of Acenocoumarol. Front. Bioeng. Biotechnol. 2022, 10, 848501. [Google Scholar] [CrossRef]
- Warzecha, Z.; Sendur, P.; Ceranowicz, P.; Dembiński, M.; Cieszkowski, J.; Kuśnierz-Cabala, B.; Olszanecki, R.; Tomaszewska, R.; Ambroży, T.; Dembiński, A. Protective Effect of Pretreatment with Acenocoumarol in Cerulein-Induced Acute Pancreatitis. Int. J. Mol. Sci. 2016, 17, 1709. [Google Scholar] [CrossRef] [Green Version]
- Warzecha, Z.; Sendur, P.; Ceranowicz, P.; Cieszkowski, J.; Dembiński, M.; Sendur, R.; Bonior, J.; Jaworek, J.; Ambroży, T.; Olszanecki, R.; et al. Therapeutic Effect of Low Doses of Acenocoumarol in the Course of Ischemia/Reperfusion-Induced Acute Pancreatitis in Rats. Int. J. Mol. Sci. 2017, 18, 882. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Raeth, T.; Beuerle, T.; Beerhues, L. A novel 4-hydroxycoumarin biosynthetic pathway. Plant Mol. Biol. 2010, 72, 17–25. [Google Scholar] [CrossRef]
- Nigg, H.N.; Nordby, H.E.; Beier, R.C.; Dillman, A.; Macias, C.; Hansen, R.C. Phototoxic coumarins in limes. Food Chem. Toxicol. 1993, 31, 331–335. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.H.; Ma, J.N.; Park, J.H.; Jung, H.W.; Park, Y.K. Anti-inflammatory and antioxidant effects of MOK, a polyherbal extract, on lipopolysaccharide-stimulated RAW 264.7 macrophages. Int. J. Mol. Med. 2019, 43, 26–36. [Google Scholar] [CrossRef]
- Zhang, X.; Xiong, H.; Liu, L. Effects of taraxasterol on inflammatory responses in lipopolysaccharide-induced RAW 264.7 macrophages. J. Ethnopharmacol. 2012, 141, 206–211. [Google Scholar] [CrossRef]
- Choi, H.S.; Seo, H.S.; Kim, S.R.; Choi, Y.K.; Shin, Y.C.; Ko, S.G. Anti-inflammatory and anti-proliferative effect of herbal medicines (APR) in RAW264.7 cells. Mol. Med. Rep. 2014, 9, 1569–1574. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Gao, D.; Du, M.; Cheng, X.; Mao, X. Casein glycomacropeptide hydrolysates inhibit PGE2 production and COX2 expression in LPS-stimulated RAW 264.7 macrophage cells via Akt mediated NF-κB and MAPK pathways. Food Funct. 2018, 9, 2524–2532. [Google Scholar] [CrossRef]
- Lenzlinger, P.M.; Morganti-Kossmann, M.C.; Laurer, H.L.; McIntosh, T.K. The duality of the inflammatory response to traumatic brain injury. Mol. Neurobiol. 2001, 24, 169–181. [Google Scholar] [CrossRef]
- Li, Q.; Yang, S.; Yang, S.; Xin, F.; Wang, M. Anti-inflammatory activity of phlomisoside F isolated from Phlomis younghusbandii Mukerjee. Int. Immunopharmacol. 2015, 28, 724–730. [Google Scholar] [CrossRef]
- Peng, F.; Yin, H.; Du, B.; Niu, K.; Yang, Y.; Wang, S. Anti-inflammatory effect of flavonoids from chestnut flowers in lipopolysaccharide-stimulated RAW 264.7 macrophages and acute lung injury in mice. J. Ethnopharmacol. 2022, 290, 115086. [Google Scholar] [CrossRef]
- Duan, X.; Li, J.; Cu, J.; Dong, Y.; Xin, X.; Aisa, H.A. Anti-inflammatory activity of Anchusa italica Retz. in LPS-stimulated RAW264.7 cells mediated by the Nrf2/HO-1, MAPK and NF-κB signaling pathways. J. Ethnopharmacol. 2022, 286, 114899. [Google Scholar] [CrossRef]
- Liu, Q.; Xiao, X.H.; Hu, L.B.; Jie, H.Y.; Wang, Y.; Ye, W.C.; Li, M.M.; Liu, Z. Anhuienoside C Ameliorates Collagen-Induced Arthritis through Inhibition of MAPK and NF-κB Signaling Pathways. Front. Pharmacol. 2017, 8, 299. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Ahn, D.U.; Xia, M.; Zeng, Q.; Li, X.; Cai, Z. Fab Fragment of Immunoglobulin Y Modulates NF-κB and MAPK Signaling through TLR4 and αVβ3 Integrin and Inhibits the Inflammatory Effect on R264.7 Macrophages. J. Agric. Food Chem. 2021, 69, 8747–8757. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, H.-J.; Hyun, C.-G. Acenocoumarol Exerts Anti-Inflammatory Activity via the Suppression of NF-κB and MAPK Pathways in RAW 264.7 Cells. Molecules 2023, 28, 2075. https://doi.org/10.3390/molecules28052075
Han H-J, Hyun C-G. Acenocoumarol Exerts Anti-Inflammatory Activity via the Suppression of NF-κB and MAPK Pathways in RAW 264.7 Cells. Molecules. 2023; 28(5):2075. https://doi.org/10.3390/molecules28052075
Chicago/Turabian StyleHan, Hyun-Ju, and Chang-Gu Hyun. 2023. "Acenocoumarol Exerts Anti-Inflammatory Activity via the Suppression of NF-κB and MAPK Pathways in RAW 264.7 Cells" Molecules 28, no. 5: 2075. https://doi.org/10.3390/molecules28052075