
Sugar Beet Pulp as a Biorefinery Substrate for Designing Feed



Abstract
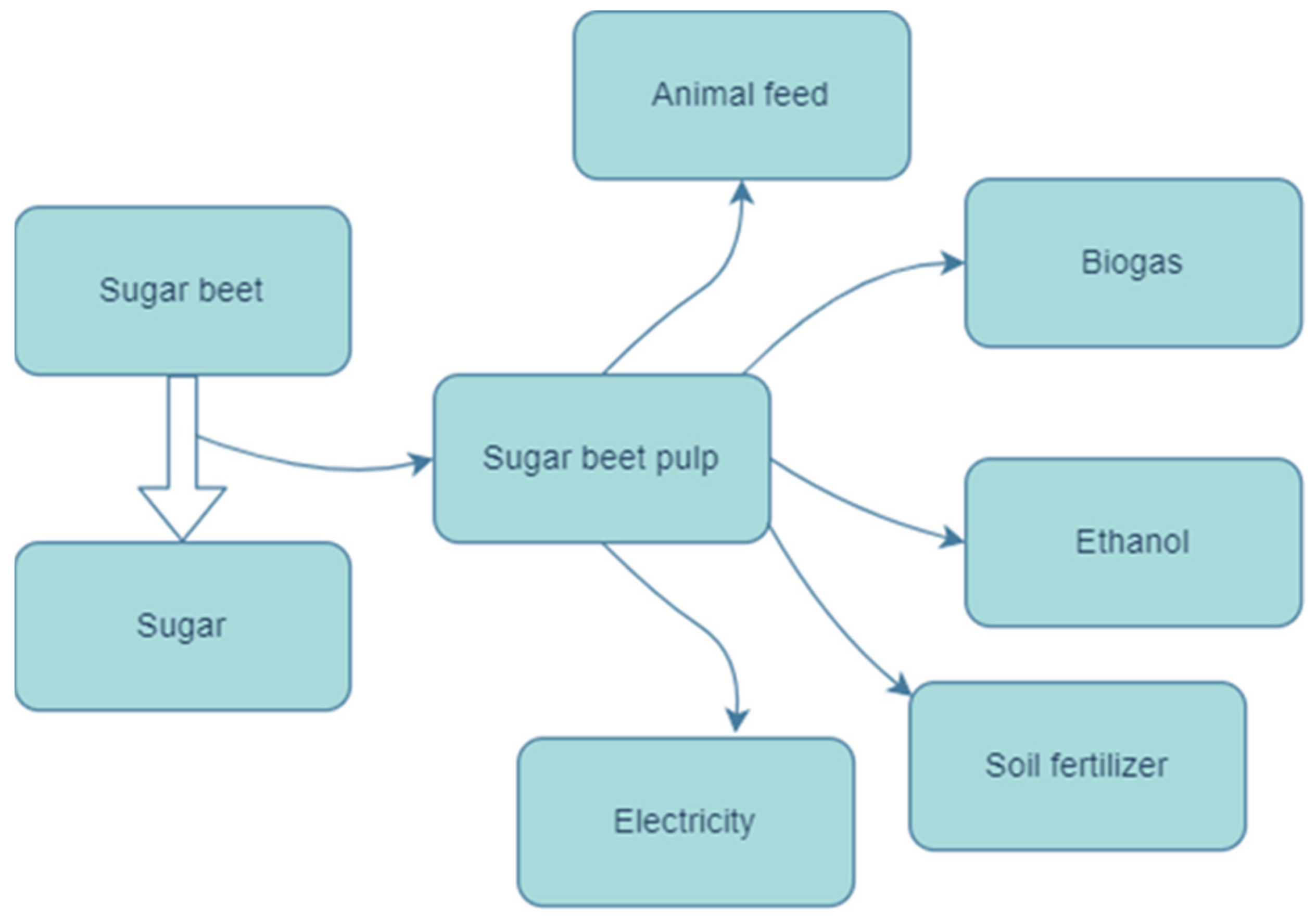
:1. Introduction
2. Results
2.1. Determination of Carbohydrates
2.2. Yeast Growth
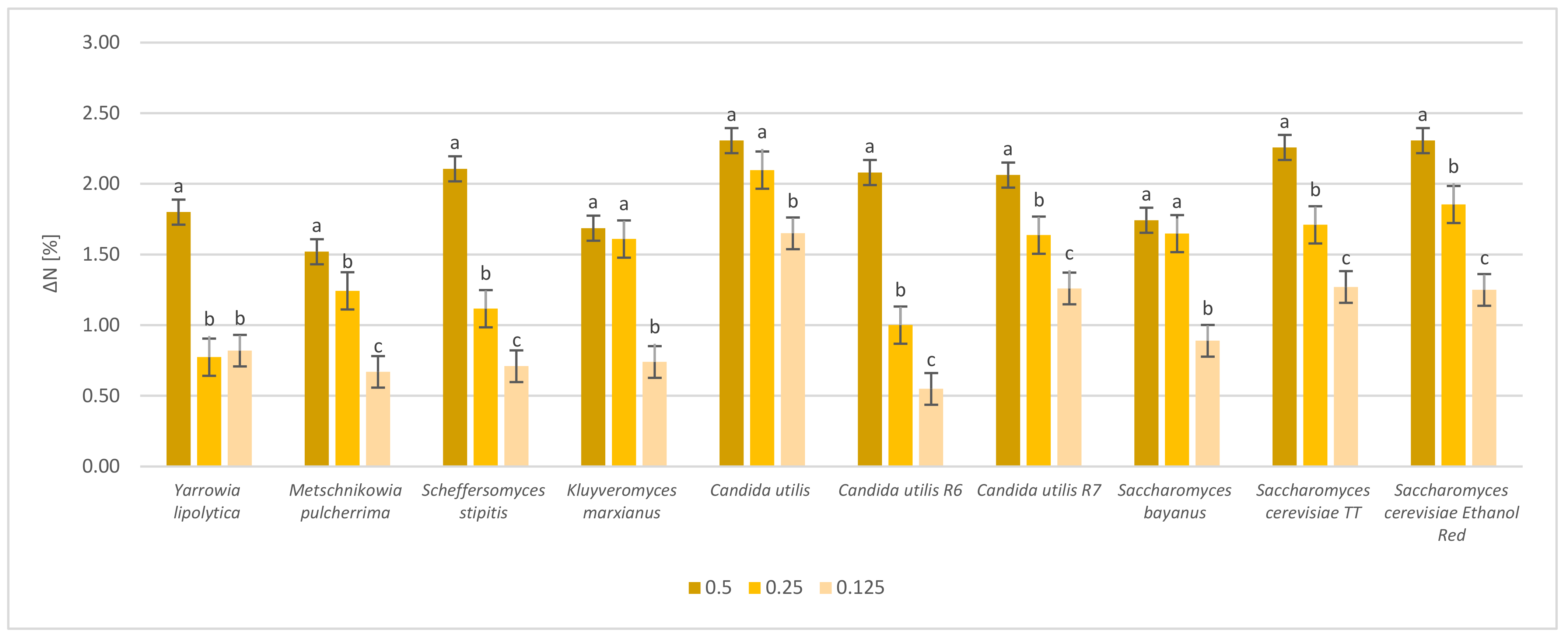
2.3. Protein Content
2.4. Free Amino Nitrogen (FAN) Content in Fresh and Dried Sugar Beet Pulp Samples
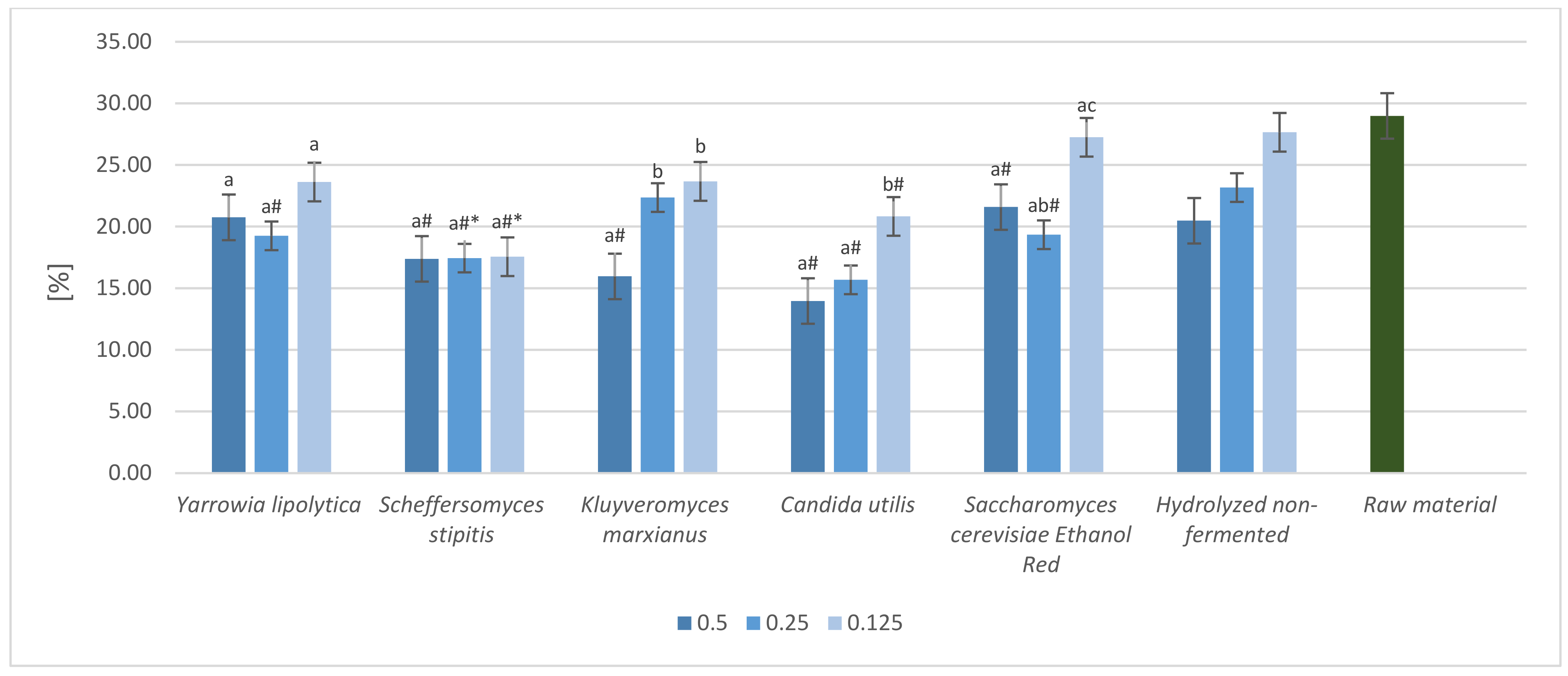
2.5. Crude Fiber Content
3. Discussion
4. Materials and Methods
4.1. Research Material
4.2. Yeasts Strains
4.3. Enzyme Preparations
4.4. Sample Preparation
4.5. Simultaneous Sachcrification and Fermentation (SSF)
4.6. Carbohydrates Determinantion
4.7. Microbial Growth
4.8. Protein Content Determination
4.9. Free Amino Nitrogen (FAN) Content Determination
4.10. Determination of Crude Fiber Content
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- UN General Assembly Transforming Our World: The 2030 Agenda for Sustainable Developmentary 2023. Available online: https://www.refworld.org/docid/57b6e3e44.html (accessed on 5 January 2023).
- Ptak, M.; Skowrońska, A.; Pińkowska, H.; Krzywonos, M. Sugar Beet Pulp in the Context of Developing the Concept of Circular Bioeconomy. Energies 2021, 15, 175. [Google Scholar] [CrossRef]
- Modelska, M.; Binczarski, M.J.; Dziugan, P.; Nowak, S.; Romanowska-Duda, Z.; Sadowski, A.; Witońska, I.A. Potential of Waste Biomass from the Sugar Industry as a Source of Furfural and Its Derivatives for Use as Fuel Additives in Poland. Energies 2020, 13, 6684. [Google Scholar] [CrossRef]
- Som-ard, J.; Atzberger, C.; Izquierdo-Verdiguier, E.; Vuolo, F.; Immitzer, M. Remote Sensing Applications in Sugarcane Cultivation: A Review. Remote Sens. 2021, 13, 4040. [Google Scholar] [CrossRef]
- World Integrated Trade Solutions Beet-Pulp, Bagasse and Other Waste of Sugar Manufacture; Whether or Not in the Form of Pellets Exports by Country in 2021. Available online: https://wits.worldbank.org/trade/comtrade/en/country/ALL/year/2021/tradeflow/Exports/partner/WLD/product/230320 (accessed on 4 September 2022).
- Alves de Oliveira, R.; Schneider, R.; Hoss Lunelli, B.; Vaz Rossell, C.E.; Maciel Filho, R.; Venus, J. A Simple Biorefinery Concept to Produce 2G-Lactic Acid from Sugar Beet Pulp (SBP): A High-Value Target Approach to Valorize a Waste Stream. Molecules 2020, 25, 2113. [Google Scholar] [CrossRef]
- Cárdenas-Fernández, M.; Bawn, M.; Hamley-Bennett, C.; Bharat, P.K.V.; Subrizi, F.; Suhaili, N.; Ward, D.P.; Bourdin, S.; Dalby, P.A.; Hailes, H.C.; et al. An Integrated Biorefinery Concept for Conversion of Sugar Beet Pulp into Value-Added Chemicals and Pharmaceutical Intermediates. Faraday Discuss. 2017, 202, 415–431. [Google Scholar] [CrossRef] [Green Version]
- Kühnel, S.; Schols, H.A.; Gruppen, H. Aiming for the Complete Utilization of Sugar-Beet Pulp: Examination of the Effects of Mild Acid and Hydrothermal Pretreatment Followed by Enzymatic Digestion. Biotechnol. Biofuels 2011, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Berłowska, J.; Pielech-Przybylska, K.; Balcerek, M.; Dziekońska-Kubczak, U.; Patelski, P.; Dziugan, P.; Kręgiel, D. Simultaneous Saccharification and Fermentation of Sugar Beet Pulp for Efficient Bioethanol Production. BioMed Res. Int. 2016, 2016, 3154929. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.; Shahzad, F.; Rahman, H.; Rajab, M. Effects of Sugar Beet Pulp Based Total Mixed Ration on Growth Performance and Blood Profile Status in Male Nili Ravi Buffalo Calves. Turk. J. Vet. Anim. Sci. 2020, 44, 928–933. [Google Scholar] [CrossRef]
- Klein, B.C.; de Mesquita Sampaio, I.L.; Mantelatto, P.E.; Filho, R.M.; Bonomi, A. Beyond Ethanol, Sugar, and Electricity: A Critical Review of Product Diversification in Brazilian Sugarcane Mills. Biofuels Bioprod. Biorefining 2019, 13, 809–821. [Google Scholar] [CrossRef]
- Ubando, A.T.; Felix, C.B.; Chen, W.-H. Biorefineries in Circular Bioeconomy: A Comprehensive Review. Bioresour. Technol. 2020, 299, 122585. [Google Scholar] [CrossRef]
- Usmani, Z.; Sharma, M.; Diwan, D.; Tripathi, M.; Whale, E.; Jayakody, L.N.; Moreau, B.; Thakur, V.K.; Tuohy, M.; Gupta, V.K. Valorization of Sugar Beet Pulp to Value-Added Products: A Review. Bioresour. Technol. 2022, 346, 126580. [Google Scholar] [CrossRef]
- Demirbas, A. Waste Management, Waste Resource Facilities and Waste Conversion Processes. Energy Convers. Manag. 2011, 52, 1280–1287. [Google Scholar] [CrossRef]
- Münnich, M.; Khiaosa-ard, R.; Klevenhusen, F.; Hilpold, A.; Khol-Parisini, A.; Zebeli, Q. A Meta-Analysis of Feeding Sugar Beet Pulp in Dairy Cows: Effects on Feed Intake, Ruminal Fermentation, Performance, and Net Food Production. Anim. Feed Sci. Technol. 2017, 224, 78–89. [Google Scholar] [CrossRef]
- Kelly, P. Sugar Beet Pulp—A Review. Anim. Feed Sci. Technol. 1983, 8, 1–18. [Google Scholar] [CrossRef]
- Berlowska, J.; Cieciura-Wloch, W.; Kalinowska, H.; Kregiel, D.; Borowski, S.; Pawlikowska, E.; Binczarski, M.; Witonska, I. Enzymatic Conversion of Sugar Beet Pulp: A Comparison of Simultaneous Saccharification and Fermentation and Separate Hydrolysis and Fermentation for Lactic Acid Production. Food Technol. Biotechnol. 2018, 56, 188. [Google Scholar] [CrossRef]
- Berlowska, J.; Pielech-Przybylska, K.; Balcerek, M.; Cieciura, W.; Borowski, S.; Kregiel, D. Integrated Bioethanol Fermentation/Anaerobic Digestion for Valorization of Sugar Beet Pulp. Energies 2017, 10, 1255. [Google Scholar] [CrossRef] [Green Version]
- Al-Mhanna, N.; Huebner, H.; Buchholz, R. Analysis of the Sugar Content in Food Products by Using Gas Chromatography Mass Spectrometry and Enzymatic Methods. Foods 2018, 7, 185. [Google Scholar] [CrossRef] [Green Version]
- Tomaszewska, J.; Bieliński, D.; Binczarski, M.; Berlowska, J.; Dziugan, P.; Piotrowski, J.; Stanishevsky, A.; Witońska, I.A. Products of Sugar Beet Processing as Raw Materials for Chemicals and Biodegradable Polymers. RSC Adv. 2018, 8, 3161–3177. [Google Scholar] [CrossRef] [Green Version]
- Dygas, D.; Nowak, S.; Olszewska, J.; Szymańska, M.; Mroczyńska-Florczak, M.; Berłowska, J.; Dziugan, P.; Kręgiel, D. Ability of Yeast Metabolic Activity to Reduce Sugars and Stabilize Betalains in Red Beet Juice. Fermentation 2021, 7, 105. [Google Scholar] [CrossRef]
- Dygas, D.; Janicka, P.; Berłowska, J.; Kręgiel, D. Conventional and Unconventional Yeasts Able to Grow on Rapeseed Meal Hydrolysates. BioResources 2022, 17, 3082–3094. [Google Scholar] [CrossRef]
- Yang, Z.; Jiang, L.; Zhang, M.; Deng, Y.; Suo, W.; Zhang, H.; Wang, C.; Li, H. Bioconversion of Apple Pomace into Microbial Protein Feed Based on Extrusion Pretreatment. Appl. Biochem. Biotechnol. 2022, 194, 1496–1509. [Google Scholar] [CrossRef]
- Zhu, W.; He, Q.; Gao, H.; Nitayavardhana, S.; Khanal, S.K.; Xie, L. Bioconversion of Yellow Wine Wastes into Microbial Protein via Mixed Yeast-Fungus Cultures. Bioresour. Technol. 2020, 299, 122565. [Google Scholar] [CrossRef]
- Mastella, L.; Senatore, V.; Beltrani, T.; Branduardi, P. Scheffersomyces stipitis Ability to Valorize Different Residual Biomasses for Vitamin B 9 Production. Microb. Biotechnol. 2022, 16, 392–403. [Google Scholar] [CrossRef]
- Tian, Y.; Li, J.; Meng, J.; Li, J. High-Yield Production of Single-Cell Protein from Starch Processing Wastewater Using Co-Cultivation of Yeasts. Bioresour. Technol. 2023, 370, 128527. [Google Scholar] [CrossRef]
- Gervasi, T.; Pellizzeri, V.; Calabrese, G.; Di Bella, G.; Cicero, N.; Dugo, G. Production of Single Cell Protein (SCP) from Food and Agricultural Waste by Using Saccharomyces Cerevisiae. Nat. Prod. Res. 2018, 32, 648–653. [Google Scholar] [CrossRef]
- Larroude, M.; Rossignol, T.; Nicaud, J.-M.; Ledesma-Amaro, R. Synthetic Biology Tools for Engineering Yarrowia lipolytica. Biotechnol. Adv. 2018, 36, 2150–2164. [Google Scholar] [CrossRef]
- Wilkowska, A.; Berlowska, J.; Nowak, A.; Motyl, I.; Antczak-Chrobot, A.; Wojtczak, M.; Kunicka-Styczyńska, A.; Binczarski, M.; Dziugan, P. Combined Yeast Cultivation and Pectin Hydrolysis as an Effective Method of Producing Prebiotic Animal Feed from Sugar Beet Pulp. Biomolecules 2020, 10, 724. [Google Scholar] [CrossRef]
- Yang, R.; Chen, Z.; Hu, P.; Zhang, S.; Luo, G. Two-Stage Fermentation Enhanced Single-Cell Protein Production by Yarrowia lipolytica from Food Waste. Bioresour. Technol. 2022, 361, 127677. [Google Scholar] [CrossRef]
- Li, Q.; Wang, D.; Li, A.; Gu, J. Microbial Lipids Production from Wastes by Metschnikowia pulcherrima: A Review. Chin. J. Biotechnol. 2021, 37, 2753–2764. [Google Scholar] [CrossRef]
- Yadav, J.S.S.; Bezawada, J.; Ajila, C.M.; Yan, S.; Tyagi, R.D.; Surampalli, R.Y. Mixed Culture of Kluyveromyces marxianus and Candida krusei for Single-Cell Protein Production and Organic Load Removal from Whey. Bioresour. Technol. 2014, 164, 119–127. [Google Scholar] [CrossRef]
- Ermis, E.; Özkan, M. Sugar Beet Powder Production Using Different Drying Methods, Characterization and Influence on Sensory Quality of Cocoa-Hazelnut Cream. J. Food Sci. Technol. 2021, 58, 2068–2077. [Google Scholar] [CrossRef]
- Yan, J.; Han, B.; Gui, X.; Wang, G.; Xu, L.; Yan, Y.; Madzak, C.; Pan, D.; Wang, Y.; Zha, G.; et al. Engineering Yarrowia lipolytica to Simultaneously Produce Lipase and Single Cell Protein from Agro-Industrial Wastes for Feed. Sci. Rep. 2018, 8, 758. [Google Scholar] [CrossRef] [Green Version]
- Ruchala, J.; Kurylenko, O.O.; Dmytruk, K.V.; Sibirny, A.A. Construction of Advanced Producers of First- and Second-Generation Ethanol in Saccharomyces Cerevisiae and Selected Species of Non-Conventional Yeasts (Scheffersomyces stipitis, Ogataea polymorpha). J. Ind. Microbiol. Biotechnol. 2020, 47, 109–132. [Google Scholar] [CrossRef] [Green Version]
- Santos, S.C.; de Sousa, A.S.; Dionísio, S.R.; Tramontina, R.; Ruller, R.; Squina, F.M.; Vaz Rossell, C.E.; da Costa, A.C.; Ienczak, J.L. Bioethanol Production by Recycled Scheffersomyces stipitis in Sequential Batch Fermentations with High Cell Density Using Xylose and Glucose Mixture. Bioresour. Technol. 2016, 219, 319–329. [Google Scholar] [CrossRef]
- Karim, A.; Gerliani, N.; Aïder, M. Kluyveromyces marxianus: An Emerging Yeast Cell Factory for Applications in Food and Biotechnology. Int. J. Food Microbiol. 2020, 333, 108818. [Google Scholar] [CrossRef]
- Kim, J.K.; Tak, K.-T.; Moon, J.-H. A Continuous Fermentation of Kluyveromyces Fragilis for the Production of a Highly Nutritious Protein Diet. Aquac. Eng. 1998, 18, 41–49. [Google Scholar] [CrossRef]
- Ritala, A.; Häkkinen, S.T.; Toivari, M.; Wiebe, M.G. Single Cell Protein—State-of-the-Art, Industrial Landscape and Patents 2001–2016. Front. Microbiol. 2017, 8, 2009. [Google Scholar] [CrossRef] [Green Version]
- Rajoka, M.I.; Khan, S.H.; Jabbar, M.A.; Awan, M.S.; Hashmi, A.S. Kinetics of Batch Single Cell Protein Production from Rice Polishings with Candida utilis in Continuously Aerated Tank Reactors. Bioresour. Technol. 2006, 97, 1934–1941. [Google Scholar] [CrossRef]
- Razzaq, Z.U.; Khan, M.K.I.; Maan, A.A.; Rahman, S.U. Characterization of Single Cell Protein from Saccharomyces Cerevisiae for Nutritional, Functional and Antioxidant Properties. J. Food Meas. Charact. 2020, 14, 2520–2528. [Google Scholar] [CrossRef]
- Shimizu, H.; Akamatsu, F.; Kamada, A.; Koyama, K.; Iwashita, K.; Goto-Yamamoto, N. Variation in the Mineral Composition of Wine Produced Using Different Winemaking Techniques. J. Biosci. Bioeng. 2020, 130, 166–172. [Google Scholar] [CrossRef]
- Seyis, I.; Aksoz, N. Effect of Carbon and Nitrogen Sources on Xylanase Production by Trichoderma Harzianum 1073 D3. Int. Biodeterior. Biodegrad. 2005, 55, 115–119. [Google Scholar] [CrossRef]
- Prior, K.J.; Bauer, F.F.; Divol, B. The Utilisation of Nitrogenous Compounds by Commercial Non-Saccharomyces Yeasts Associated with Wine. Food Microbiol. 2019, 79, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Boudreau, T.F.; Peck, G.M.; O’Keefe, S.F.; Stewart, A.C. Free Amino Nitrogen Concentration Correlates to Total Yeast Assimilable Nitrogen Concentration in Apple Juice. Food Sci. Nutr. 2018, 6, 119–123. [Google Scholar] [CrossRef]
- Vuong, M.-D.; Thanh, N.-T.; Son, C.-K.; Yves, W. Protein Enrichment of Cassava-Based Dried Distiller’s Grain by Solid State Fermentation Using Trichoderma Harzianum and Yarrowia lipolytica for Feed Ingredients. Waste Biomass Valorization 2021, 12, 3875–3888. [Google Scholar] [CrossRef]
- Wang, W.; Wei, H.; Alahuhta, M.; Chen, X.; Hyman, D.; Johnson, D.K.; Zhang, M.; Himmel, M.E. Heterologous Expression of Xylanase Enzymes in Lipogenic Yeast Yarrowia lipolytica. PLoS ONE 2014, 9, e111443. [Google Scholar] [CrossRef] [Green Version]
- Balagurunathan, B.; Jonnalagadda, S.; Tan, L.; Srinivasan, R. Reconstruction and Analysis of a Genome-Scale Metabolic Model for Scheffersomyces stipitis. Microb. Cell Factories 2012, 11, 27. [Google Scholar] [CrossRef] [Green Version]
- Ma, M.; Wang, X.; Zhang, X.; Zhao, X. Alcohol Dehydrogenases from Scheffersomyces stipitis Involved in the Detoxification of Aldehyde Inhibitors Derived from Lignocellulosic Biomass Conversion. Appl. Microbiol. Biotechnol. 2013, 97, 8411–8425. [Google Scholar] [CrossRef]
- Agbogbo, F.K.; Coward-Kelly, G. Cellulosic Ethanol Production Using the Naturally Occurring Xylose-Fermenting Yeast, Pichia Stipitis. Biotechnol. Lett. 2008, 30, 1515–1524. [Google Scholar] [CrossRef]
- Unrean, P.; Nguyen, N.H.A. Optimized Fed-Batch Fermentation of Scheffersomyces stipitis for Efficient Production of Ethanol from Hexoses and Pentoses. Appl. Biochem. Biotechnol. 2013, 169, 1895–1909. [Google Scholar] [CrossRef]
- Ray, M.K.; Devi, K.U.; Kumar, G.S.; Shivaji, S. Extracellular Protease from the Antarctic Yeast Candida Humicola. Appl. Environ. Microbiol. 1992, 58, 1918–1923. [Google Scholar] [CrossRef] [Green Version]
- Rajoka, M.I.; Ahmed, S.; Hashmi, A.S.; Athar, M. Production of Microbial Biomass Protein from Mixed Substrates by Sequential Culture Fermentation of Candida utilis and Brevibacterium lactofermentum. Ann. Microbiol. 2012, 62, 1173–1179. [Google Scholar] [CrossRef]
- Smith, I.M.; Baker, A.; Arneborg, N.; Jespersen, L. Non-Saccharomyces yeasts protect against epithelial cell barrier disruption induced by Salmonella enterica subsp. enterica serovar Typhimurium. Lett. Appl. Microbiol. 2015, 61, 491–497. [Google Scholar] [CrossRef]
- Foligné, B.; Lafont, F.; Daniel, C. Study of the persistence and dynamics of recombinant mCherry-producing Yarrowia lipolytica strains in the mouse intestine using fluorescence imaging. Microb. Biotechnol. 2022, in press. [Google Scholar] [CrossRef]
- Singh, A.K.; Kim, W.K. Effects of Dietary Fiber on Nutrients Utilization and Gut Health of Poultry: A Review of Challenges and Opportunities. Animals 2021, 11, 181. [Google Scholar] [CrossRef]
- Allen, M.S.; Mertens, D.R. Evaluating Constraints on Fiber Digestion by Rumen Microbes. J. Nutr. 1988, 118, 261–270. [Google Scholar] [CrossRef]
- Wilfart, A.; Montagne, L.; Simmins, P.H.; van Milgen, J.; Noblet, J. Sites of Nutrient Digestion in Growing Pigs: Effect of Dietary Fiber1. J. Anim. Sci. 2007, 85, 976–983. [Google Scholar] [CrossRef]
- Brown, M.S.; Ponce, C.H.; Pulikanti, R. Adaptation of Beef Cattle to High-Concentrate Diets: Performance and Ruminal Metabolism1. J. Anim. Sci. 2006, 84, E25–E33. [Google Scholar] [CrossRef]
- Jeon, S.; Jeong, S.; Lee, M.; Seo, J.; Kam, D.K.; Kim, J.H.; Park, J.; Seo, S. Effects of Reducing Inclusion Rate of Roughages by Changing Roughage Sources and Concentrate Types on Intake, Growth, Rumen Fermentation Characteristics, and Blood Parameters of Hanwoo Growing Cattle (Bos Taurus coreanae). Asian-Australas. J. Anim. Sci. 2019, 32, 1705–1714. [Google Scholar] [CrossRef] [Green Version]
- Shabbir, A.; Sharif, M.; Ashfaq, K.; Aqib, A.; Saeed, M.; Di Cerbo, A.; Alagawany, M. Effect of Yeast-Fermented Citrus Pulp as a Protein Source on Nutrient Intake, Digestibility, Nitrogen Balance and In Situ Digestion Kinetics in Nili Ravi Buffalo Bulls. Animals 2021, 11, 1713. [Google Scholar] [CrossRef]
- Berlowska, J.; Binczarski, M.; Dudkiewicz, M.; Kalinowska, H.; Witonska, I.A.; Stanishevsky, A.V. A Low-Cost Method for Obtaining High-Value Bio-Based Propylene Glycol from Sugar Beet Pulp. RSC Adv. 2015, 5, 2299–2304. [Google Scholar] [CrossRef]
- Geisler, J.; Weiß, N. Free Amino Nitrogen (FAN) Measurement in Beer Using an Eppendorf BioSpectrometer®. Eppendorf 2015, 9, 3. [Google Scholar]
- European Commission. Commission Regulation (EC) No 152/2009 of 27 January 2009 Laying down the Methods of Sampling and Analysis for the Official Control of Feed (Text with EEA Relevance). Off. J. Eur. Union 2009, 54, 2–54. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbohydrate | Fresh Sugar Beet Pulp | Dried Sugar Beet Pulp | ||||||
|---|---|---|---|---|---|---|---|---|
| Enzymatic Treatment [mL/10 g DM] | Enzymatic Treatment [mL/10 g DM] | |||||||
| 0 (Without) | 0.5 | 0.25 | 0.125 | 0 (Without) | 0.5 | 0.25 | 0.125 | |
| D-Xylose | 0.002 ± 0.001 | 1.113 ± 0.032 | 0.856 ± 0.031 | 0.826 ± 0.045 | 0.013 ± 0.003 | 1.046 ± 0.045 | 0.761 ± 0.050 | 0.681 ± 0.009 |
| D-Mannose | 0.011 ± 0.004 | 0.649 ± 0.046 | 0.088 ± 0.012 | 0.013 ± 0.003 | 0.021 ± 0.006 | 0.431 ± 0.020 | 0.261 ± 0.010 | 0.201 ± 0.021 |
| D-Fructose | 0.044 ± 0.010 | 0.655 ± 0.027 | 0.081 ± 0.008 | 0.587 ± 0.065 | 0.051 ± 0.011 | 0.564 ± 0.016 | 0.201 ± 0.031 | 0.164 ± 0.043 |
| D-Glucose | 0.049 ± 0.009 | 16.823 ± 0.097 | 14.277 ± 0.056 | 13.217 ± 0.080 | 0.059 ± 0.008 | 14.374 ± 0.080 | 13.031 ± 0.094 | 11.264 ± 0.032 |
| Yeast Strain | 0.5 | 0.25 | 0.125 | Control |
|---|---|---|---|---|
| Yarrowia lipolytica | AV: 1.54 × 107 a# | AV: 1.32 × 107 a | AV: 1.90 × 107 a# | AV: 6.50 × 106 |
| SD: 5.90 × 106 | SD: 4.60 × 106 | SD: 4.00 × 106 | SD: 1.29 × 106 | |
| Metschnikowia pulcherrima | AV: 2.46 × 107 ab | AV: 2.38 × 107 b | AV: 4.66 × 107 a# | AV: 7.05 × 106 |
| SD: 3.58 × 106 | SD: 4.49 × 106 | SD: 2.34 × 107 | SD: 6.86 × 105 | |
| Scheffersomyces stipitis | AV: 2.82 × 108 a# | AV: 1.13 × 108 ab | AV: 3.32 × 107 b | AV: 3.18 × 107 |
| SD: 1.31 × 108 | SD: 3.22 × 107 | SD: 1.64 × 107 | SD: 8.88 × 106 | |
| Kluyveromyces marxianus | AV: 4.82 × 107 a# | AV: 2.10 × 107 b | AV: 2.52 × 107 b | AV: 1.34 × 107 |
| SD: 8.11 × 106 | SD: 3.67 × 106 | SD: 9.56 × 106 | SD: 7.33 × 106 | |
| Candida utilis | AV: 2.23 × 108 a# | AV: 2.01 × 108 a# | AV: 2.60 × 108 a# | AV: 2.43 × 107 |
| SD: 8.54 × 107 | SD: 4.10 × 107 | SD: 6.39 × 107 | SD: 1.69 × 107 | |
| Candida utilis R6 | AV: 3.74 × 107 ab | AV: 5.78 × 107 a# | AV: 2.26 × 107 b | AV: 2.34 × 107 |
| SD: 6.91 × 106 | SD: 2.29 × 107 | SD: 1.10 × 107 | SD: 5.29 × 106 | |
| Candida utilis R7 | AV: 4.12 × 107 a | AV: 4.78 × 107 a | AV: 2.13 × 108 b# | AV: 5.95 × 107 |
| SD: 7.26 × 106 | SD: 4.79 × 107 | SD: 5.90 × 107 | SD: 2.14 × 107 | |
| Saccharomyces bayanus BC S103 | AV: 1.34 × 108 a# | AV: 1.63 × 108 a# | AV: 1.60 × 108 a# | AV: 3.56 × 107 |
| SD: 2.08 × 107 | SD: 2.52 × 107 | SD: 5.51 × 107 | SD: 1.11 × 107 | |
| Saccharomyces cerevisiae TT | AV: 1.07 × 108 ab | AV: 5.53 × 107 a | AV: 1.75 × 108 b# | AV: 7.23 × 107 |
| SD: 2.69 × 107 | SD: 5.21 × 106 | SD: 8.73 × 107 | SD: 1.74 × 107 | |
| Saccharomyces cerevisiae Ethanol Red | AV: 2.16 × 108 a# | AV: 1.87 × 108 a# | AV: 2.05 × 108 a# | AV: 8.08 × 107 |
| SD: 2.29 × 107 | SD: 1.18 × 107 | SD: 1.64 × 107 | SD: 9.43 × 106 |
| Yeast Strain | 0.5 | 0.25 | 0.125 | Control |
|---|---|---|---|---|
| Yarrowia lipolytica | AV: 1.36 × 108 a# | AV: 1.63 × 108 a# | AV: 1.38 × 108 a# | AV: 2.46 × 107 |
| SD: 2.93 × 107 | SD: 5.94 × 107 | SD: 4.64 × 107 | SD: 3.92 × 106 | |
| Metschnikowia pulcherrima | AV: 5.46 × 107 a | AV: 2.88 × 107 ab | AV: 1.20 × 108 b# | AV: 2.46 × 107 |
| SD: 1.13 × 107 | SD: 5.84 × 106 | SD: 2.40 × 107 | SD: 2.08 × 106 | |
| Scheffersomyces stipitis | AV: 9.34 × 107 a# | AV: 6.68 × 107 ab# | AV: 1.26 × 108 b# | AV: 3.38 × 107 |
| SD: 1.06 × 107 | SD: 9.04 × 106 | SD: 1.28 × 107 | SD: 1.74 × 107 | |
| Kluyveromyces marxianus | AV: 1.20 × 108 a# | AV: 5.36 × 107 b | AV: 5.42 × 107 b | AV: 1.22 × 107 |
| SD: 2.00 × 107 | SD: 3.86 × 107 | SD: 1.22 × 107 | SD: 1.29 × 106 | |
| Candida utilis | AV: 1.10 × 108 a | AV: 1.12 × 108 a | AV: 1.62 × 108 a | AV: 1.37 × 108 |
| SD: 1.60 × 107 | SD: 1.84 × 107 | SD: 5.44 × 107 | SD: 1.74 × 107 | |
| Candida utilis R6 | AV: 2.14 × 107 a | AV: 2.40 × 107 a# | AV: 1.16 × 107 a# | AV: 2.75 × 106 |
| SD: 4.48 × 106 | SD: 9.20 × 106 | SD: 1.52 × 106 | SD: 1.00 × 106 | |
| Candida utilis R7 | AV: 5.67 × 107 a | AV: 4.96 × 107 b | AV: 2.80 × 107 c | AV: 3.80 × 107 |
| SD: 2.89 × 106 | SD: 1.23 × 107 | SD: 9.60 × 106 | SD: 3.89 × 106 | |
| Saccharomyces bayanus BC S103 | AV: 1.23 × 108 a# | AV: 1.36 × 108 b# | AV: 1.26 × 108 b# | AV: 1.27 × 107 |
| SD: 1.34 × 107 | SD: 1.68 × 107 | SD: 1.52 × 107 | SD: 2.26 × 106 | |
| Saccharomyces cerevisiae TT | AV: 4.30 × 107 a | AV: 1.33 × 108 b# | AV: 2.04 × 108 b# | AV: 1.14 × 107 |
| SD: 1.40 × 107 | SD: 2.98 × 107 | SD: 6.48 × 107 | SD: 6.25 × 106 | |
| Saccharomyces cerevisiae Ethanol Red | AV: 1.34 × 108 a# | AV: 1.64 × 108 ab# | AV: 1.92 × 108 b# | AV: 3.54 × 107 |
| SD: 1.52 × 107 | SD: 2.72 × 107 | SD: 3.04 × 107 | SD: 7.68 × 106 |
| Yeast Strain | Strain Code |
|---|---|
| Yarrowia lipolytica | LOCK 0264 |
| Metschnikowia pulcherrima | NCYC 747 |
| Scheffersomyces stipitis | NCYC 1541 |
| Kluyveromyces marxianus | LOCK 0024 |
| Candida utilis | LOCK 0021 |
| Candida utilis | R6 |
| Candida utilis | R7 |
| Saccharomyces bayanus BC S103 | Fermentis Lesaffre for Beverages |
| Saccharomyces cerevisiae TT | LOCK 0105 |
| Saccharomyces cerevisiae Ethanol Red | Leaf/Lesaffre Advanced Fermentation |
| Enzyme Preparation | Cellulase | Invertase | Xylanase | Pectinase |
|---|---|---|---|---|
| Viscozyme | 20.9 | 61.2 | 25.9 | 312.6 |
| Ultraflo Max | 32.7 | 1.8 | 64.7 | 21.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dygas, D.; Kręgiel, D.; Berłowska, J. Sugar Beet Pulp as a Biorefinery Substrate for Designing Feed. Molecules 2023, 28, 2064. https://doi.org/10.3390/molecules28052064
Dygas D, Kręgiel D, Berłowska J. Sugar Beet Pulp as a Biorefinery Substrate for Designing Feed. Molecules. 2023; 28(5):2064. https://doi.org/10.3390/molecules28052064
Chicago/Turabian StyleDygas, Dawid, Dorota Kręgiel, and Joanna Berłowska. 2023. "Sugar Beet Pulp as a Biorefinery Substrate for Designing Feed" Molecules 28, no. 5: 2064. https://doi.org/10.3390/molecules28052064