
Yohimbine Alleviates Oxidative Stress and Suppresses Aerobic Cysteine Metabolism Elevated in the Rat Liver of High-Fat Diet-Fed Rats

 , , , , and
, , , , and
Abstract
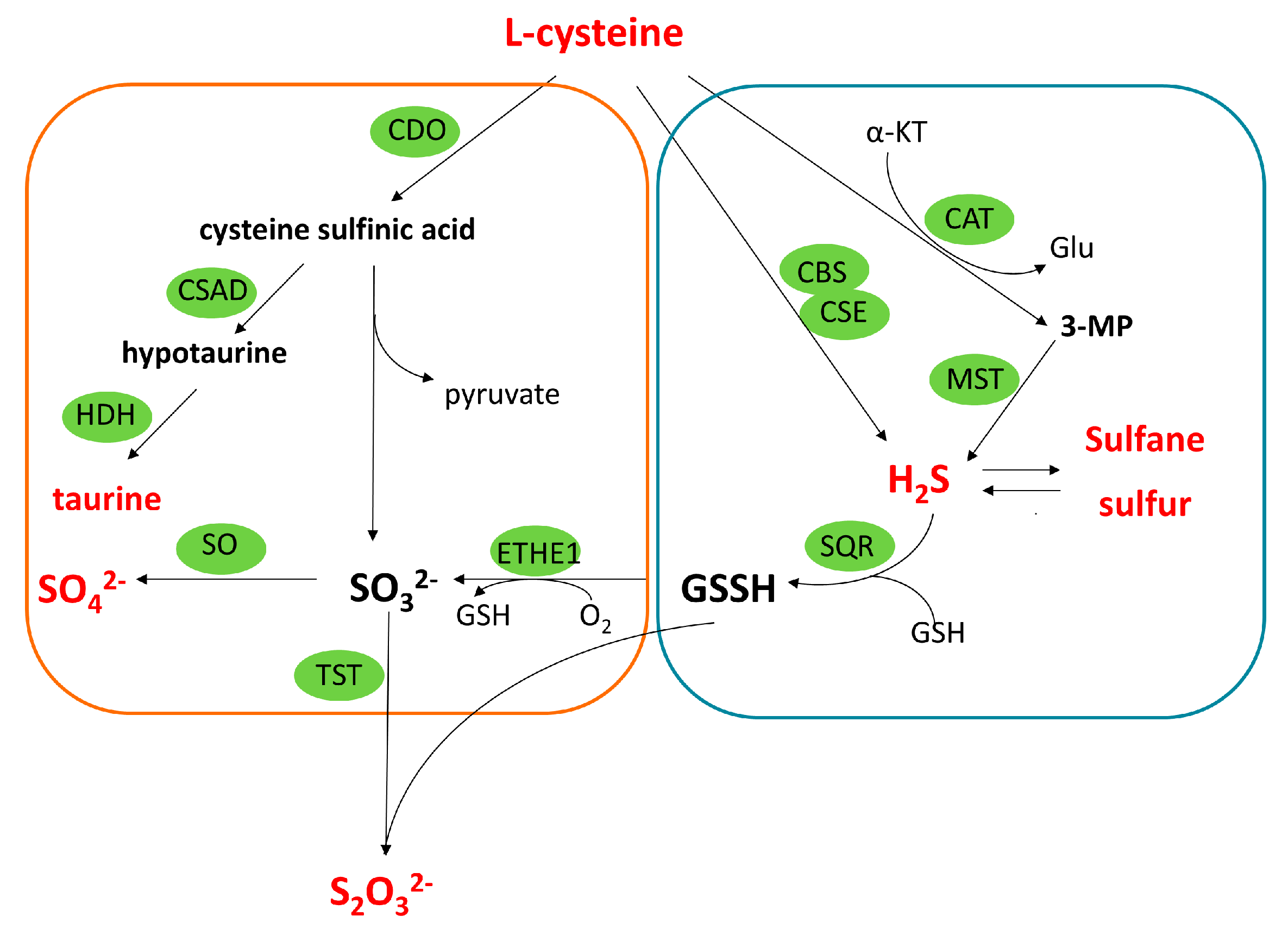
:1. Introduction
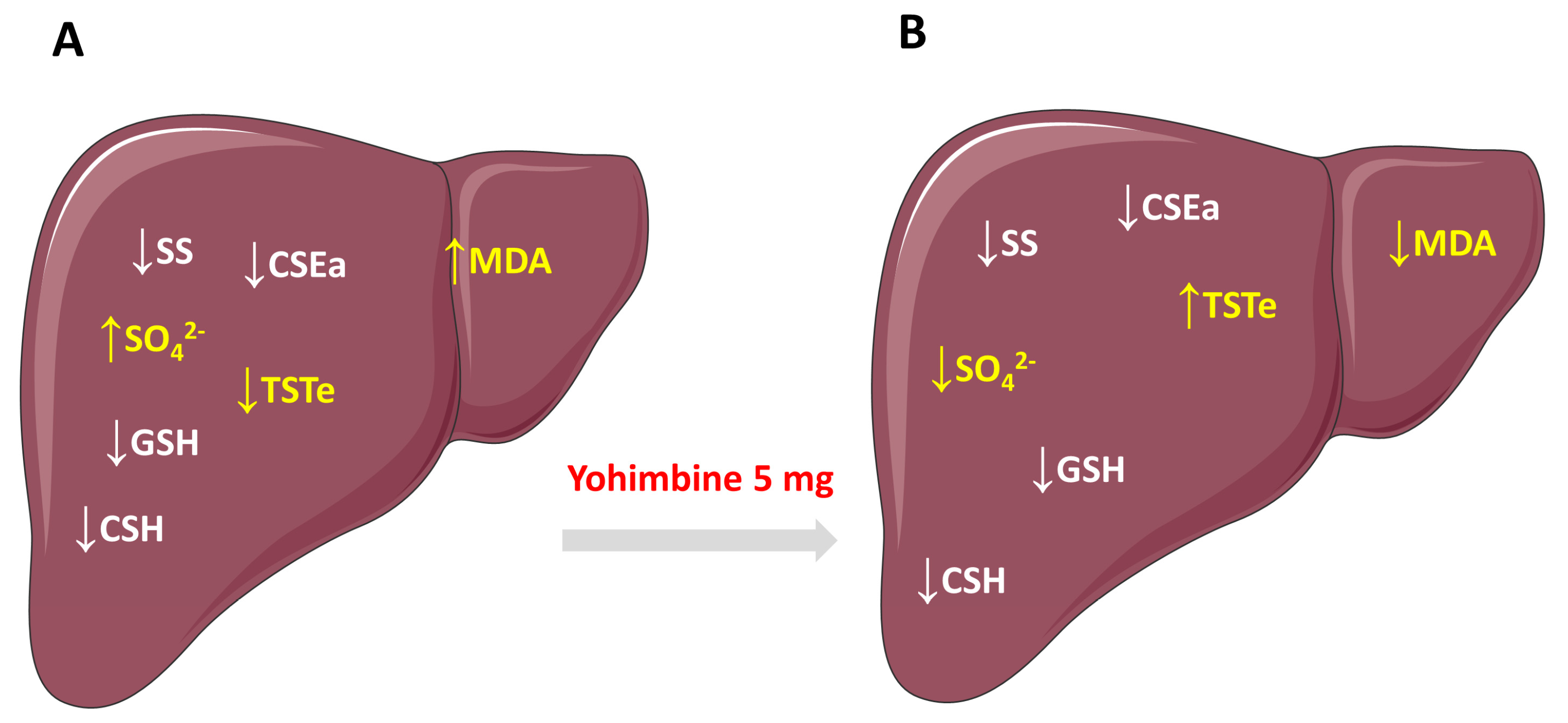
2. Results
2.1. The Effect of HFD and Yohimbine Treatment on the Changes in Body Weight
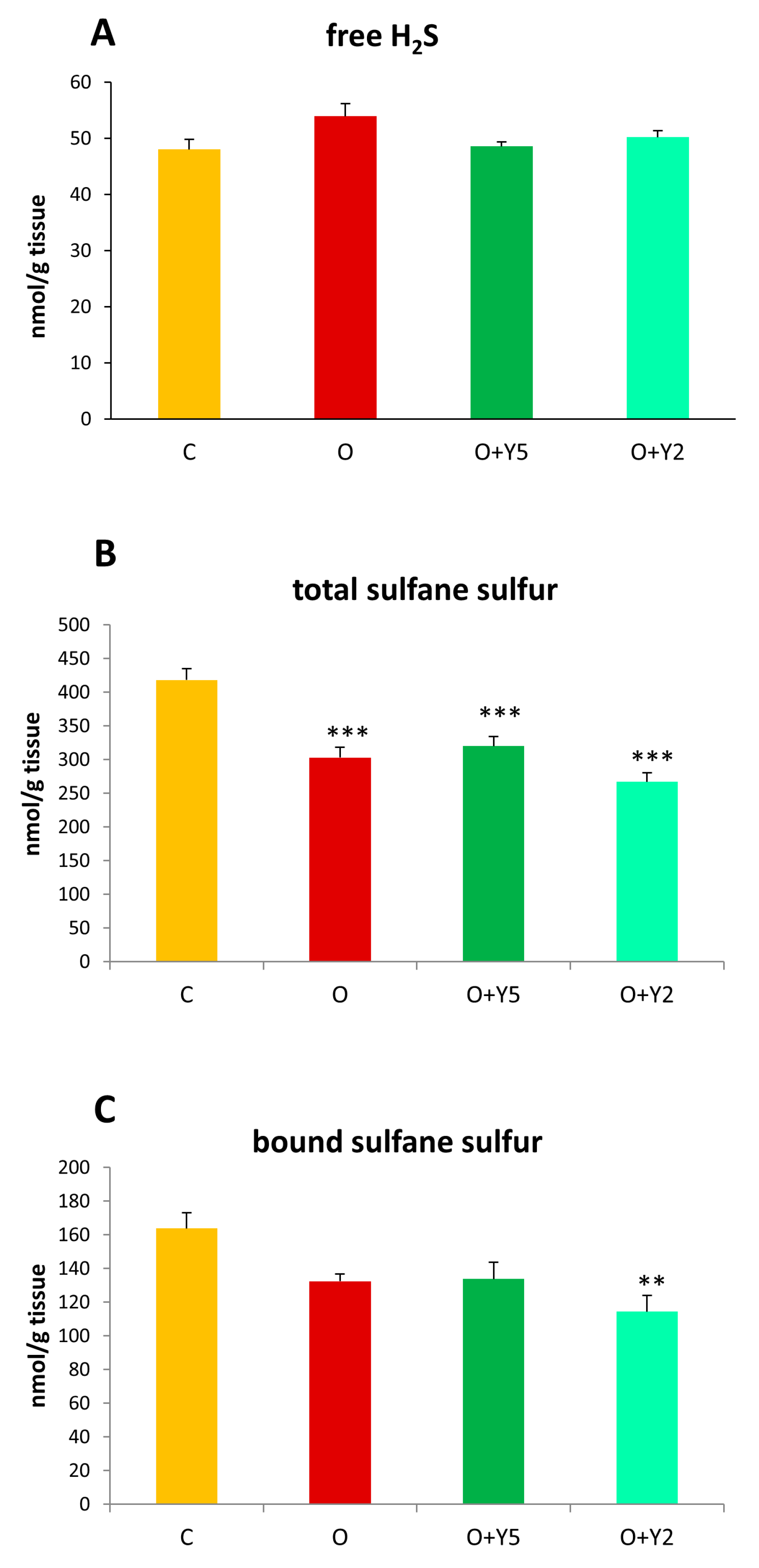
2.2. The Effect of HFD and Yohimbine Treatment on the Level of Free Sulfide, Sulfane Sulfur, and Bound Sulfane Sulfur in the Rat Liver
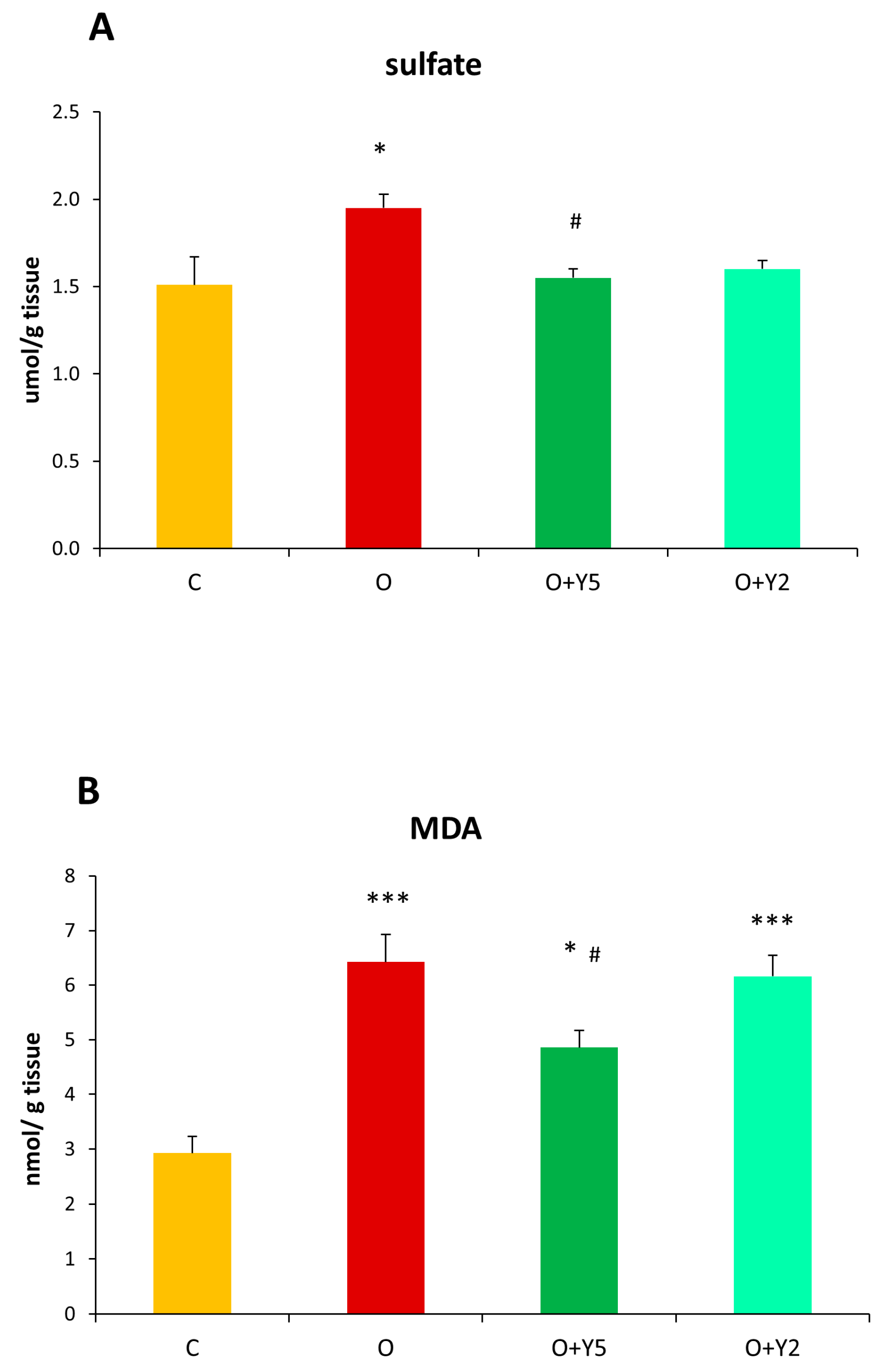
2.3. The Effect of HFD and Yohimbine Treatment on the Sulfate Concentration in the Rat Liver
2.4. The Effect of HFD and Yohimbine Treatment on the Level Malonyl Dialdehyde (MDA) as a Marker of Oxidative Stress
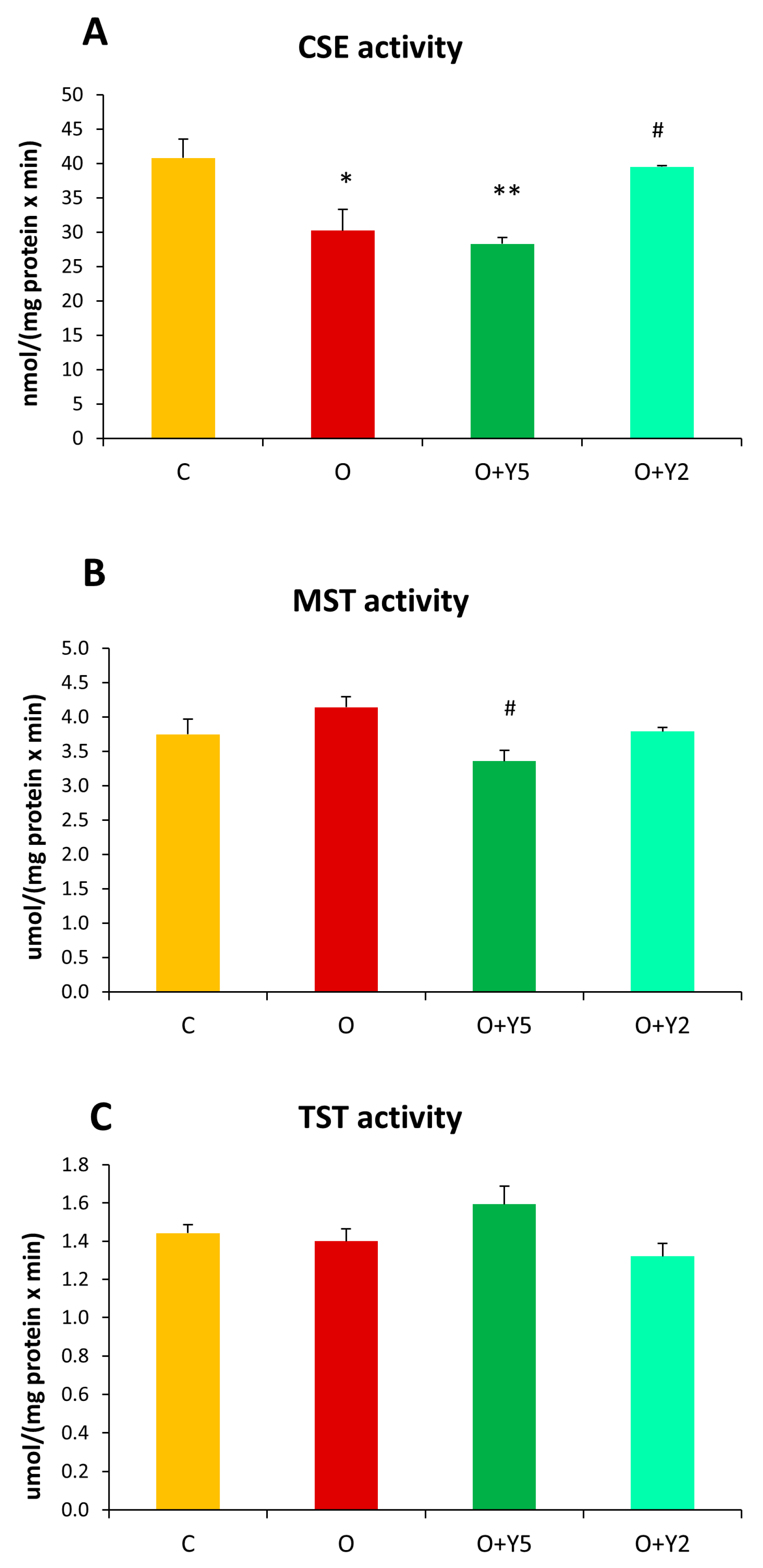
2.5. The Effect of HFD and Yohimbine Treatment on the Activity of Enzymes Involved in Reactive Sulfur Species Formation and Transport (CSE, MST, and TST) in the Rat Liver
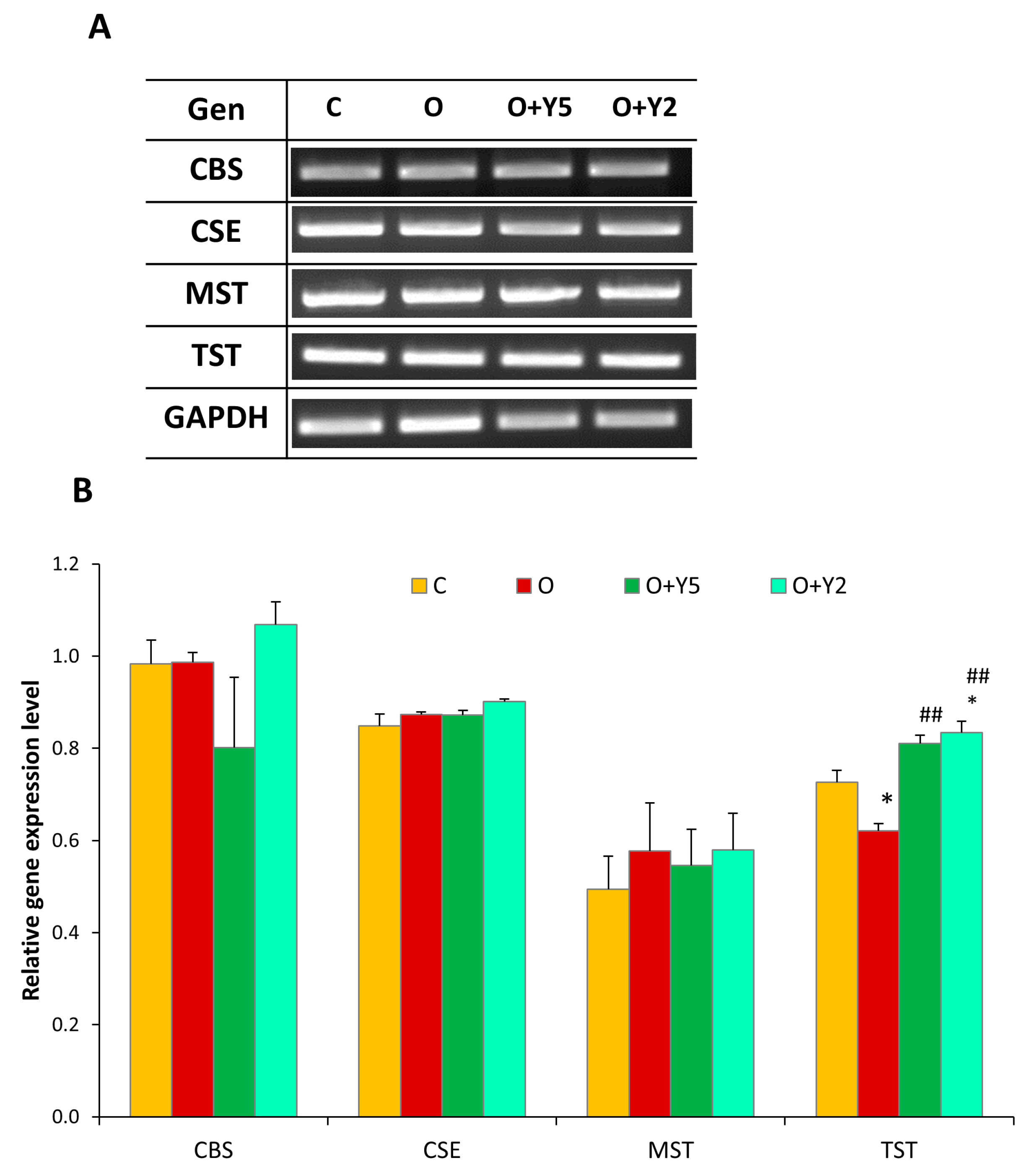
2.6. The Effect of HFD and Yohimbine Treatment on the Expression of Enzymes Involved in the Formation of Reactive Sulfur Species (CSE, MST, CBS, and TST) in the Rat Liver
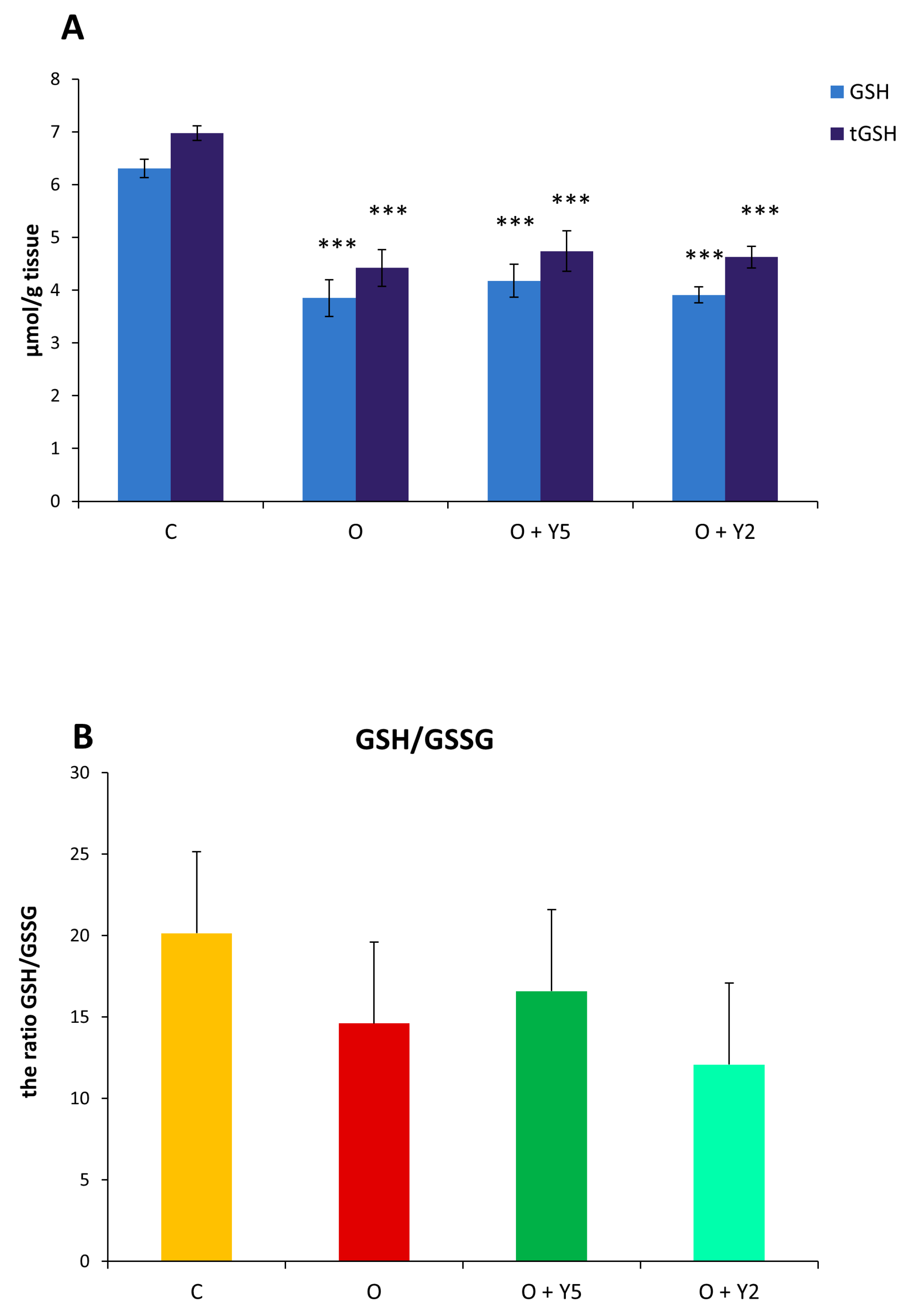
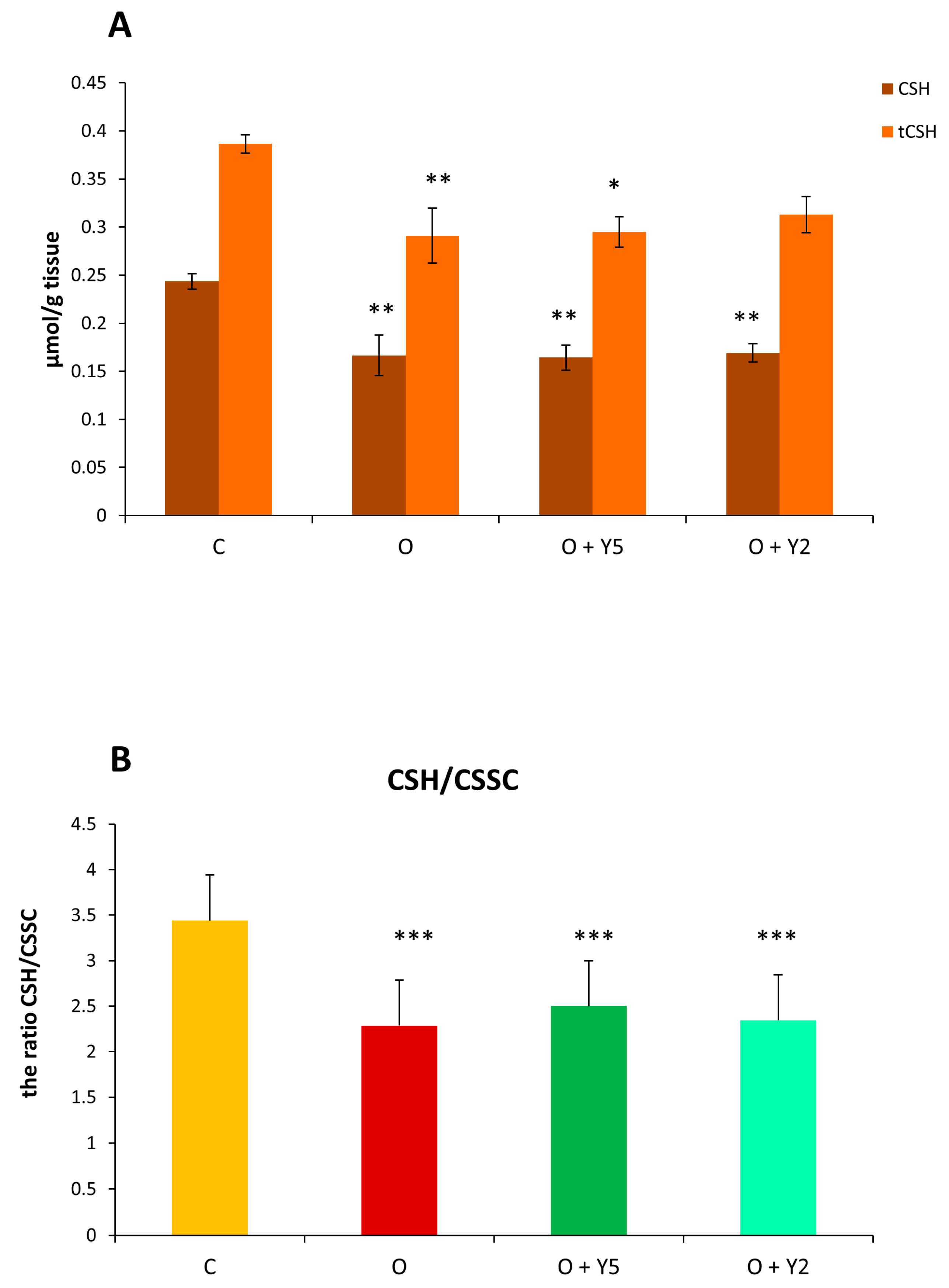
2.7. The Effect of HFD and Yohimbine Treatment on the Level of the Main Low Molecular Weight Thiols (GSH, CSH)
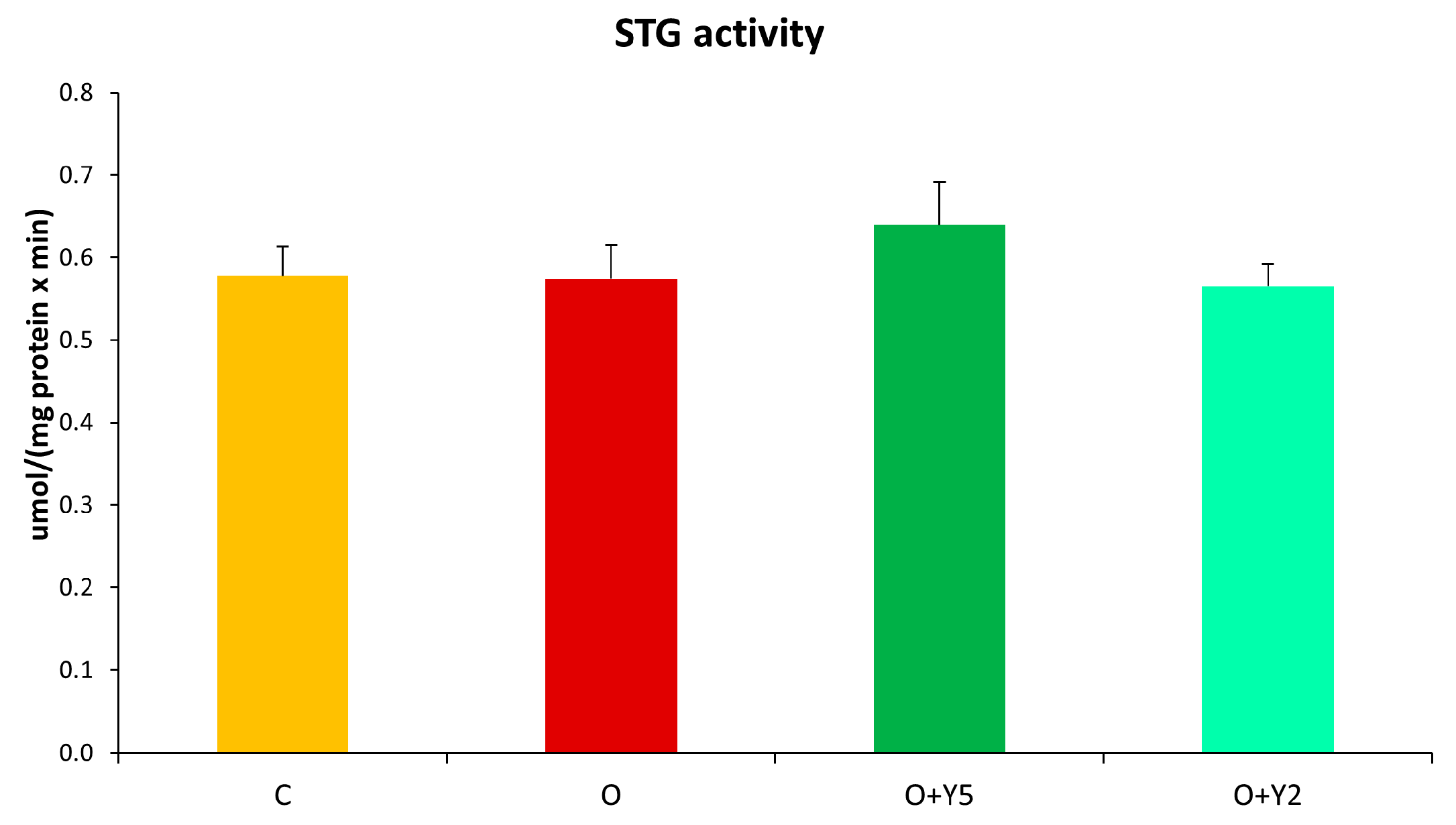
2.8. The Effect of HFD and Yohimbine Treatment on the Activity of Glutathione S-Transferase
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Obesity Induction
4.3. Chemicals
4.4. Preparation of Tissue Homogenates
4.5. Biochemical Assays
4.6. High-Performance Liquid Chromatography (HPLC) Analysis
4.6.1. Reduced Thiols
4.6.2. Total Thiols
4.6.3. Oxidized Thiols
4.7. Enzyme Expression
4.7.1. Isolation of Total RNA
4.7.2. Reverse Transcription of RNA
4.7.3. Polymerase Chain Reaction (PCR)
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Tam, S.W.; Worcel, M.; Wyllie, M. Yohimbine: A clinical review. Pharm. Ther. 2001, 91, 215–243. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, H.Y.; Xu, H.Z.; Liu, J.J.; Meng, X.G.; Zhou, M.; Ruan, H.L. Alkaloids with Immunosuppressive Activity from the Bark of Pausinystalia yohimbe. J. Nat. Prod. 2018, 81, 1841–1849. [Google Scholar] [CrossRef] [PubMed]
- Morales, A. Yohimbine in erectile dysfunction: The facts. Int. J. Impot Res. 2000, 12 (Suppl. 1), S70–S74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arteaga, M.; Motte-Lara, J.; Velázquez-Moctezuma, J. Effects of yohimbine and apomorphine on the male sexual behaviour pattern of the golden hamster (Mesocricetus auratus). Eur. Neuropsychopharmacol. 2002, 12, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Tasleem, M.; Alrehaily, A.; Almeleebia, T.M.; Alshahrani, M.Y.; Ahmad, I.; Asiri, M.; Alabdallah, N.M.; Saeed, M. Investigation of Antidepressant Properties of Yohimbine by Employing Structure-Based Computational Assessments. Curr. Issues Mol. Biol. 2021, 43, 1805–1827. [Google Scholar] [CrossRef]
- Ostojic, S.M. Yohimbine: The effects on body composition and exercise performance in soccer players. Res. Sport. Med. 2006, 14, 289–299. [Google Scholar] [CrossRef]
- McCarty, M.F. Pre-exercise administration of yohimbine may enhance the efficacy of exercise training as a fat loss strategy by boosting lipolysis. Med. Hypotheses 2002, 58, 491–495. [Google Scholar] [CrossRef]
- Loiselle, J.J.; Yang, G.; Wu, L. Hydrogen sulfide and hepatic lipid metabolism—A critical pairing for liver health. Br. J. Pharm. 2020, 177, 757–768. [Google Scholar] [CrossRef] [Green Version]
- Kabil, O.; Vitvitsky, V.; Xie, P.; Banerjee, R. The quantitative significance of the transsulfuration enzymes for H2S production in murine tissues. Antioxid. Redox Signal. 2011, 15, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Rose, P.; Moore, P.K.; Zhu, Y.Z. H2S biosynthesis and catabolism: New insights from molecular studies. Cell Mol. Life Sci. 2017, 74, 1391–1412. [Google Scholar] [CrossRef] [Green Version]
- Iciek, M.; Włodek, L. Biosynthesis and biological properties of compounds containing highly reactive, reduced sulfane sulfur. Pol. J. Pharm. 2001, 53, 215–225. [Google Scholar]
- Iciek, M.; Bilska-Wilkosz, A.; Górny, M. Sulfane sulfur—New findings on an old topic. Acta Biochim. Pol. 2019, 66, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Szabo, C.; Ransy, C.; Módis, K.; Andriamihaja, M.; Murghes, B.; Coletta, C.; Olah, G.; Yanagi, K.; Bouillaud, F. Regulation of mitochondrial bioenergetic function by hydrogen sulfide. Part I. Biochemical and physiological mechanisms. Br. J. Pharm. 2014, 171, 2099–2122. [Google Scholar] [CrossRef] [Green Version]
- Tan, G.; Pan, S.; Li, J.; Dong, X.; Kang, K.; Zhao, M.; Jiang, X.; Kanwar, J.R.; Qiao, H.; Jiang, H.; et al. Hydrogen sulfide attenuates carbon tetrachloride-induced hepatotoxicity, liver cirrhosis and portal hypertension in rats. PLoS ONE 2011, 6, e25943. [Google Scholar] [CrossRef]
- Comas, F.; Moreno-Navarrete, J.M. The Impact of H2S on Obesity-Associated Metabolic Disturbances. Antioxidants 2021, 10, 633. [Google Scholar] [CrossRef]
- Yu, Y.; Ye, S.M.; Liu, D.Y.; Yang, L.Q. AP39 ameliorates high fat diet-induced liver injury in young rats via alleviation of oxidative stress and mitochondrial impairment. Exp. Anim. 2021, 70, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Ju, Y.; Fu, M.; Zhang, Y.; Pei, Y.; Racine, M.; Baath, S.; Merritt, T.J.S.; Wang, R.; Wu, L. Cystathionine gamma-lyase/hydrogen sulfide system is essential for adipogenesis and fat mass accumulation in mice. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 165–176. [Google Scholar] [CrossRef]
- Comas, F.; Latorre, J.; Ortega, F.; Arnoriaga Rodríguez, M.; Lluch, A.; Sabater, M.; Rius, F.; Ribas, X.; Costas, M.; Ricart, W.; et al. Morbidly obese subjects show increased serum sulfide in proportion to fat mass. Int. J. Obes. (Lond.) 2021, 45, 415–426. [Google Scholar] [CrossRef]
- Dudek, M.; Knutelska, J.; Bednarski, M.; Nowiński, L.; Zygmunt, M.; Mordyl, B.; Głuch-Lutwin, M.; Kazek, G.; Sapa, J.; Pytka, K. A Comparison of the Anorectic Effect and Safety of the Alpha2-Adrenoceptor Ligands Guanfacine and Yohimbine in Rats with Diet-Induced Obesity. PLoS ONE 2015, 10, e0141327. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Deng, M.; Su, J.; Lin, Y.; Jia, Z.; Peng, K.; Wang, F.; Yang, T. Specific downregulation of cystathionine β-synthase expression in the kidney during obesity. Physiol. Rep. 2018, 6, e13630. [Google Scholar] [CrossRef] [Green Version]
- Peh, M.T.; Anwar, A.B.; Ng, D.S.; Atan, M.S.; Kumar, S.D.; Moore, P.K. Effect of feeding a high fat diet on hydrogen sulfide (H2S) metabolism in the mouse. Nitric Oxide 2014, 41, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.Y.; Sarna, L.K.; Siow, Y.L.; Karmin, O. High-fat diet stimulates hepatic cystathionine β-synthase and cystathionine γ-lyase expression. Can. J. Physiol. Pharm. 2013, 91, 913–919. [Google Scholar] [CrossRef]
- Bravo, E.; Palleschi, S.; Aspichueta, P.; Buqué, X.; Rossi, B.; Cano, A.; Napolitano, M.; Ochoa, B.; Botham, K.M. High fat diet-induced non alcoholic fatty liver disease in rats is associated with hyperhomocysteinemia caused by down regulation of the transsulphuration pathway. Lipids Health Dis. 2011, 10, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.; Gao, Q.; Li, Y.; Bai, Y.; Zhang, J.; Xu, W.; Xu, J. Flammulina velutipes Mycorrhizae Attenuate High Fat Diet-Induced Lipid Disorder, Oxidative Stress and Inflammation in the Liver and Perirenal Adipose Tissue of Mice. Nutrients 2022, 14, 3830. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, L.; Xu, D.; Deng, W.; Yang, W.; Tang, F.; Da, M. Knockout of calpain-1 protects against high-fat diet-induced liver dysfunction in mouse through inhibiting oxidative stress and inflammation. Food Sci. Nutr. 2021, 9, 367–374. [Google Scholar] [CrossRef]
- Moreno-Fernández, S.; Garcés-Rimón, M.; Vera, G.; Astier, J.; Landrier, J.F.; Miguel, M. High Fat/High Glucose Diet Induces Metabolic Syndrome in an Experimental Rat Model. Nutrients 2018, 10, 1502. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.; Jian, T.; Li, J.; Lv, H.; Tong, B.; Meng, X.; Ren, B.; Chen, J. Chicoric Acid Ameliorates Nonalcoholic Fatty Liver Disease via the AMPK/Nrf2/NFκB Signaling Pathway and Restores Gut Microbiota in High-Fat-Diet-Fed Mice. Oxid. Med. Cell. Longev. 2020, 2020, 9734560. [Google Scholar] [CrossRef] [PubMed]
- Abbas, A.M.; Sakr, H.F. Simvastatin and vitamin E effects on cardiac and hepatic oxidative stress in rats fed on high fat diet. J. Physiol. Biochem. 2013, 69, 737–750. [Google Scholar] [CrossRef]
- Yun, K.U.; Ryu, C.S.; Oh, J.M.; Kim, C.H.; Lee, K.S.; Lee, C.H.; Lee, H.S.; Kim, B.H.; Kim, S.K. Plasma homocysteine level and hepatic sulfur amino acid metabolism in mice fed a high-fat diet. Eur. J. Nutr. 2013, 52, 127–134. [Google Scholar] [CrossRef]
- Chen, J.H.; Yu, G.F.; Jin, S.Y.; Zhang, W.H.; Lei, D.X.; Zhou, S.L.; Song, X.R. Activation of α2 adrenoceptor attenuates lipopolysaccharide-induced hepatic injury. Int. J. Clin. Exp. Pathol. 2015, 8, 10752–10759. [Google Scholar]
- Santativongchai, P.; Srisuksai, K.; Parunyakul, K.; Thiendedsakul, P.; Lertwatcharasarakul, P.; Fungfuang, W.; Tulayakul, P. Effects of Crocodile Oil (Crocodylus siamensis) on Liver Enzymes: Cytochrome P450 and Glutathione S-Transferase Activities in High-fat DietFed Rats. Vet. Med. Int. 2022, 2022, 9990231. [Google Scholar] [CrossRef]
- Akbay, E.; Ulusu, N.N.; Töröner, F.; Ayvaz, G.; Taneri, F.; Aktürk, M.; Arslan, M.; Karasu, C. Effects of rosiglitazone treatment on the pentose phosphate pathway and glutathione-dependent enzymes in liver and kidney of rats fed a high-fat diet. Curr. Res. Clin. Exp. 2004, 65, 79–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Tao, Q.; Wang, X.; Zhang, X. Impact of high-fat diet on liver genes expression profiles in mice model of nonalcoholic fatty liver disease. Environ. Toxicol. Pharm. 2016, 45, 52–62. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Zhou, Z.C.; Song, C.Y.; Chen, X. The Protective Effect and Mechanism of Dexmedetomidine on Diabetic Peripheral Neuropathy in Rats. Front. Pharm. 2020, 11, 1139. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, M.; Cui, E.; Zhang, H.; Zhu, X.; Zhou, J.; Yan, M.; Sun, J. Dexmedetomidine attenuates sevoflurane-induced neurocognitive impairment through α2-adrenoceptors. Mol. Med. Rep. 2021, 23, 38. [Google Scholar] [CrossRef] [PubMed]
- Shen, K.P.; Lo, Y.C.; Yang, R.C.; Liu, H.W.; Chen, I.J.; Wu, B.N. Antioxidant eugenosedin-A protects against lipopolysaccharide-induced hypotension, hyperglycaemia and cytokine immunoreactivity in rats and mice. J. Pharm. Pharm. 2005, 57, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Brannan, R.G. Reactive sulfur species act as prooxidants in liposomal and skeletal muscle model systems. J. Agric. Food Chem. 2010, 58, 3767–3771. [Google Scholar] [CrossRef]
- Nandi, D.L.; Horowitz, P.M.; Westley, J. Rhodanese as a thioredoxin oxidase. Int. J. Biochem. Cell Biol. 2000, 32, 465–473. [Google Scholar] [CrossRef]
- Buonvino, S.; Arciero, I.; Melino, S. Thiosulfate-Cyanide Sulfurtransferase a Mitochondrial Essential Enzyme: From Cell Metabolism to the Biotechnological Applications. Int. J. Mol. Sci. 2022, 23, 8452. [Google Scholar] [CrossRef]
- Bald, E.; Glowacki, R.; Drzewoski, J. Determination by liquid chromatography of free and total cysteine in human urine in the form of its S-quinolinium derivative. J. Chromatogr. A 2001, 913, 319–329. [Google Scholar] [CrossRef]
- Wood, J.L. Sulfane sulfur. Methods Enzym. 1987, 143, 25–29. [Google Scholar] [CrossRef]
- Shen, X.; Pattillo, C.B.; Pardue, S.; Bir, S.C.; Wang, R.; Kevil, C.G. Measurement of plasma hydrogen sulfide in vivo and in vitro. Free Radic Biol. Med. 2011, 50, 1021–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogasawara, Y.; Ishii, K.; Togawa, T.; Tanabe, S. Determination of bound sulfur in serum by gas dialysis/high-performance liquid chromatography. Anal. Biochem. 1993, 215, 73–81. [Google Scholar] [CrossRef]
- Matsuo, Y.; Greenberg, D.M. A crystalline enzyme that cleaves homoserine and cystathionine. IV. Mechanism of action, reversibility, and substrate specificity. J. Biol. Chem. 1959, 234, 516–519. [Google Scholar] [CrossRef] [PubMed]
- Soda, K. A spectrophotometric microdetermination of keto acids with 3-methyl-2-benzothiazolone hydrazone. Agric. Biol. Chem. 1967, 31, 1054–1060. [Google Scholar] [CrossRef]
- Valentine, W.N.; Frankenfeld, J.K. 3-Mercaptopyruvate sulfurtransferase (EC 2.8.1.2): A simple assay adapted to human blood cells. Clin. Chim. Acta 1974, 51, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Sörbo, B. Rohodanese. Methods Enzymol. 1955, 2, 334–337. [Google Scholar]
- Rice-Evans, C.A.; Diplock, A.T.; Symons, M.C.R. Techniques in Free Radical Research, 1st ed.; Elsevier: Amsterdam, The Netherlands, 1991. [Google Scholar]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Bald, E.; Chwatko, G.; Głowacki, R.; Kuśmierek, K. Analysis of plasma thiols by high-performance liquid chromatography with ultraviolet detection. J. Chromatogr. A 2004, 1032, 109–115. [Google Scholar] [CrossRef]
- Kaczor-Kamińska, M.; Kaminski, K.; Wróbel, M. The Expression and Activity of Rhodanese, 3-Mercaptopyruvate Sulfurtransferase, Cystathionine γ-Lyase in the Most Frequently Chosen Cellular Research Models. Biomolecules 2021, 11, 1859. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Changes in Body Weight During 10 Weeks of Obesity Induction (g) | Changes in Body Weight During 30 Days of Treatment (g) | ||||
|---|---|---|---|---|---|
| C | HFD (O) | C | O | O + Y5 | O + Y2 |
| 206.7 ± 3.5 | 256.8 *** ± 4.8 | 38.7 ± 1.6 | 57.6 * ± 4.3 | 19.4 ### ± 6.1 | 37.0 # ± 8.3 |
| n = 6 | n = 18 | n = 6 | n = 6 | n = 6 | n = 6 |
| Target Gene | No. in NCBI Database | Primers (5′→3′) | Location in Gene | Product Size [bp] | Ref. |
|---|---|---|---|---|---|
| CSE | NM_017074.1 | F: CCGACGAGGAATTGCTTGGA R: ACATCACTGTGGCCGTTCAT | 203–666 | 464 | [52] |
| MST | NM_138843.1 | F: GTATCTGCTCAGTGGGTGGC R: CAGGGATGTGTCCAGGTTCG | 240–828 | 589 | |
| CBS | NM_012522.2 | F: ATGGATGCTGCAGAAAGGCT R: AGGTGGATCGGCTTGAACTG | 1302–1607 | 306 | |
| TST | NM_012808.1 | F: ACATCCGTGGCTCTGTCAAC R: TCGTAAACAGCCACATCGGG | 642–830 | 208 | [52] |
| GAPDH | NM_017008.4 | F: AGTGCCAGCCTCGTCTCATA R: GATGGTGATGGGTTTCCCGT | 50–297 | 248 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iciek, M.; Górny, M.; Kotańska, M.; Bilska-Wilkosz, A.; Kaczor-Kamińska, M.; Zagajewski, J. Yohimbine Alleviates Oxidative Stress and Suppresses Aerobic Cysteine Metabolism Elevated in the Rat Liver of High-Fat Diet-Fed Rats. Molecules 2023, 28, 2025. https://doi.org/10.3390/molecules28052025
Iciek M, Górny M, Kotańska M, Bilska-Wilkosz A, Kaczor-Kamińska M, Zagajewski J. Yohimbine Alleviates Oxidative Stress and Suppresses Aerobic Cysteine Metabolism Elevated in the Rat Liver of High-Fat Diet-Fed Rats. Molecules. 2023; 28(5):2025. https://doi.org/10.3390/molecules28052025
Chicago/Turabian StyleIciek, Małgorzata, Magdalena Górny, Magdalena Kotańska, Anna Bilska-Wilkosz, Marta Kaczor-Kamińska, and Jacek Zagajewski. 2023. "Yohimbine Alleviates Oxidative Stress and Suppresses Aerobic Cysteine Metabolism Elevated in the Rat Liver of High-Fat Diet-Fed Rats" Molecules 28, no. 5: 2025. https://doi.org/10.3390/molecules28052025