
Chlamydomonas reinhardtii: A Factory of Nutraceutical and Food Supplements for Human Health

 , , ,
, , ,
Abstract
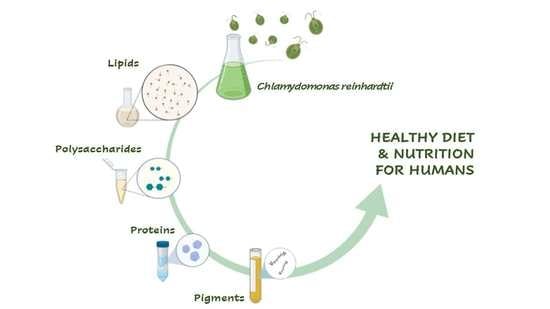
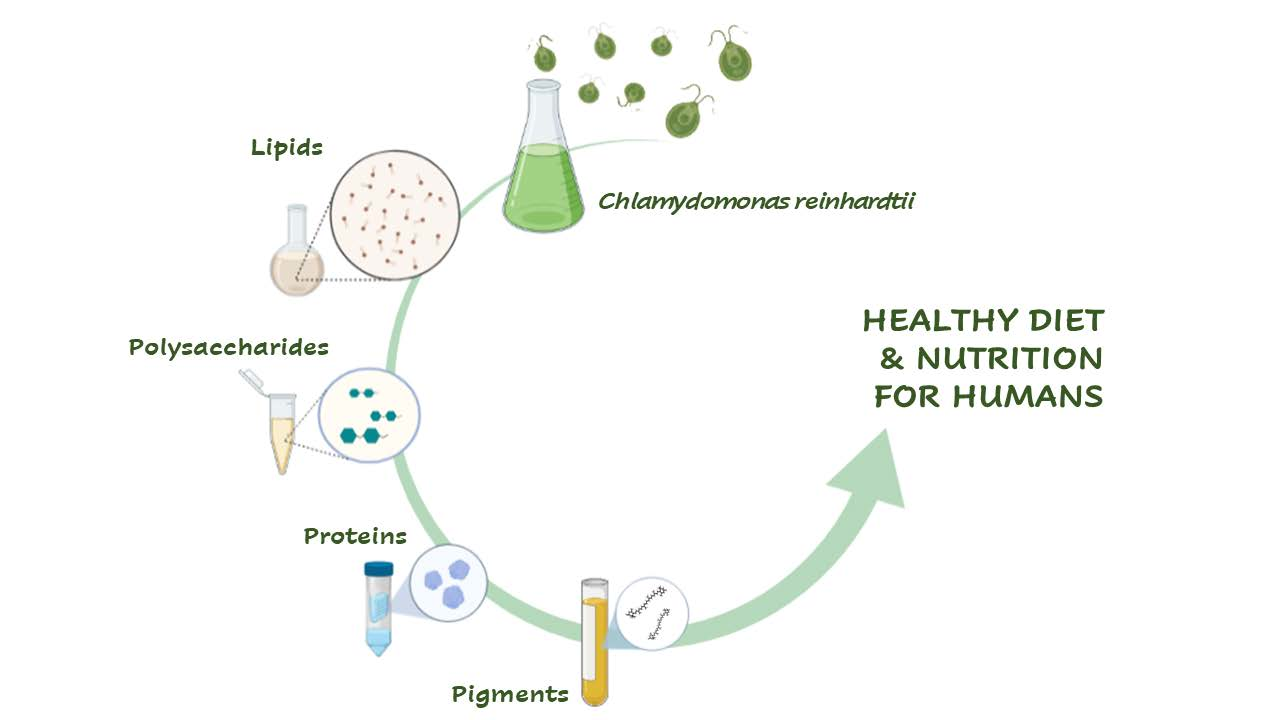
:
1. Introduction
2. Bio-production
2.1. Polysaccharides
2.2. Lipids
2.3. Recombinant Proteins for Therapeutics
2.4. Pigments
3. Nutritional Aspects: Linking with Human Diet and Related Food Products
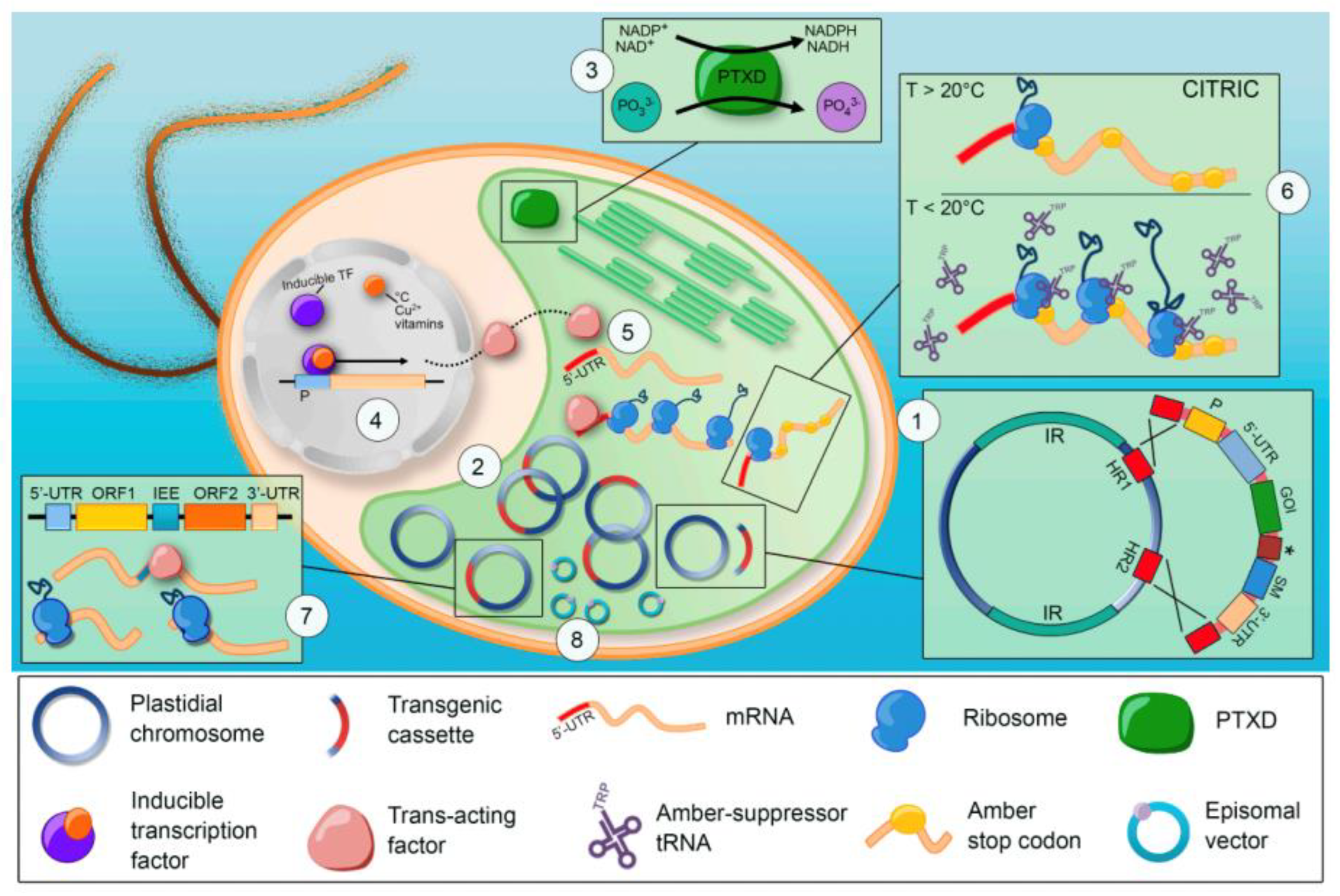
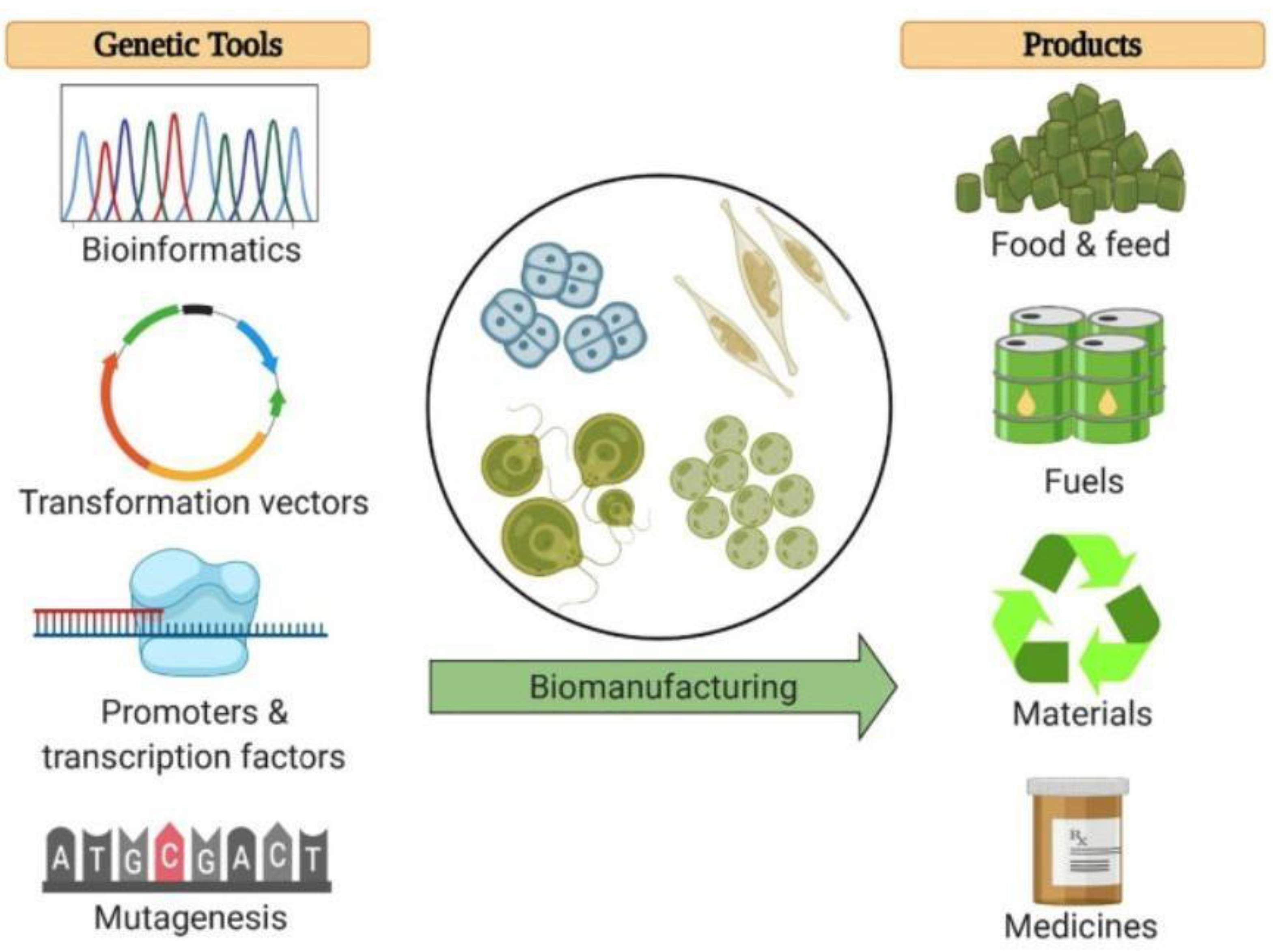
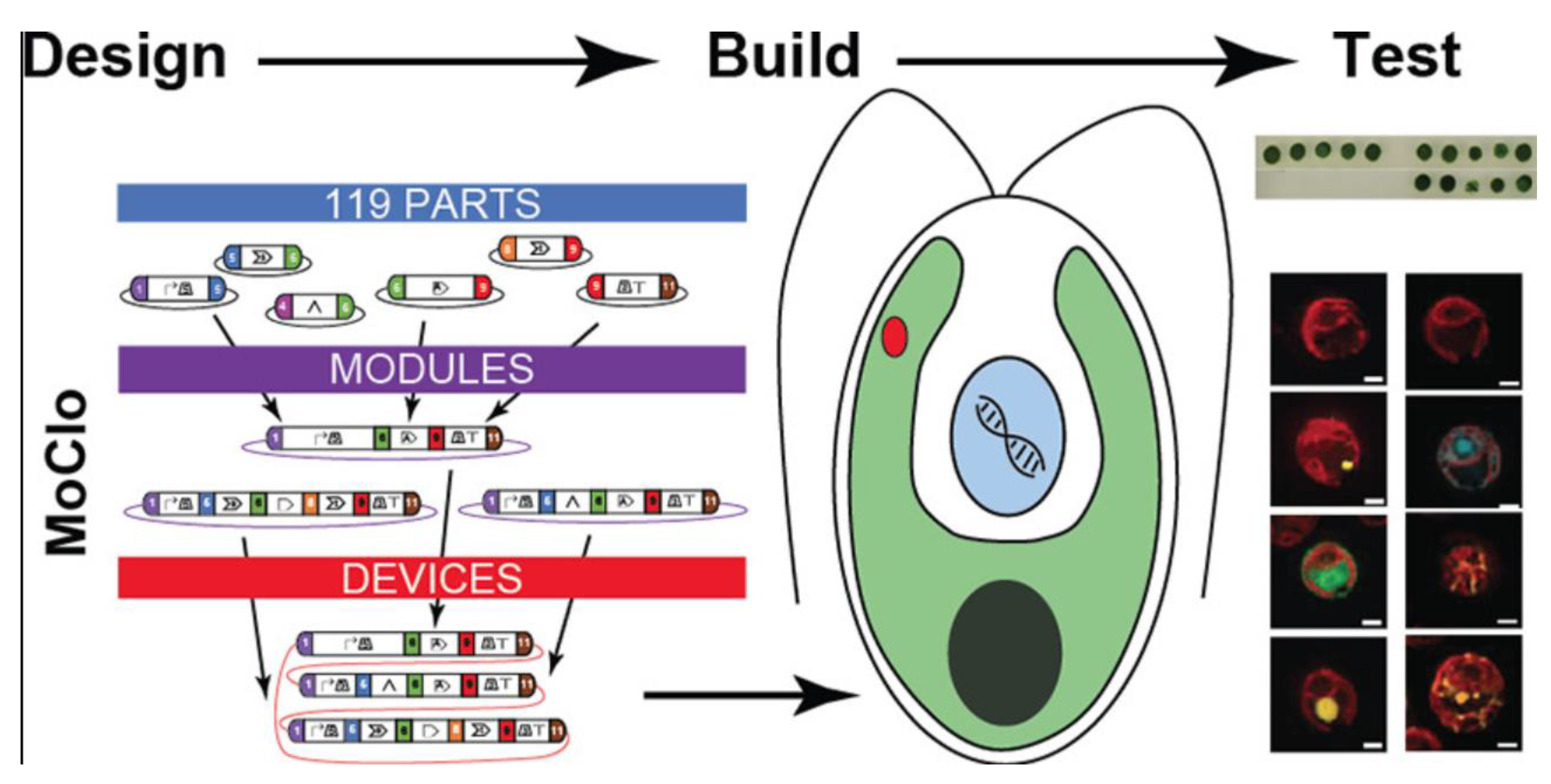
4. C. reinhardtii Molecular Genetic Toolkit: Is an Effective Road for the Market?
5. Conclusions and Future Perspectives
6. Methodology
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dyo, Y.M.; Purton, S. The Algal Chloroplast as a Synthetic Biology Platform for Production of Therapeutic Proteins. Microbiology 2018, 164, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Blanken, W.; Cuaresma, M.; Wijffels, R.H.; Janssen, M. Cultivation of Microalgae on Artificial Light Comes at a Cost. Algal Res. 2013, 2, 333–340. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from Microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef]
- Kay, R.A.; Barton, L.L. Microalgae as Food and Supplement. Crit. Rev. Food Sci. Nutr. 1991, 30, 555–573. [Google Scholar] [CrossRef] [PubMed]
- Potijun, S.; Yaisamlee, C.; Sirikhachornkit, A. Pigment Production under Cold Stress in the Green Microalga Chlamydomonas reinhardtii. Agriculture 2021, 11, 564. [Google Scholar] [CrossRef]
- Kiran, B.R.; Venkata Mohan, S. Microalgal Cell Biofactory—Therapeutic, Nutraceutical and Functional Food Applications. Plants 2021, 10, 836. [Google Scholar] [CrossRef]
- Gifuni, I.; Pollio, A.; Safi, C.; Marzocchella, A.; Olivieri, G. Current Bottlenecks and Challenges of the Microalgal Biorefinery. Trends Biotechnol. 2019, 37, 242–252. [Google Scholar] [CrossRef]
- Harris, E.H. Chlamydomonas as a model organism. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 363–406. [Google Scholar] [CrossRef] [Green Version]
- Harris, E.H. Organellar and Metabolic Processes. The Chlamydomonas Sourcebook; Stern, D.B., Witman, G.B., Eds.; Academic Press: San Diego, CA, USA, 1989; Volume 2. [Google Scholar]
- Sasso, S.; Stibor, H.; Mittag, M.; Grossman, A.R. The natural history of model organisms: From molecular manipulation of domesticated Chlamydomonas reinhardtii to survival in nature. Elife 2018, 7, e39233. [Google Scholar] [CrossRef]
- Harris, E.H. Introduction to Chlamydomonas and Its Laboratory Use. The Chlamydomonas Sourcebook; Harris, E.H., Stern, D.B., Witman, G.B., Eds.; Academic Press: Oxford, UK, 2009. [Google Scholar]
- Pröschold, T.; Harris, E.H.; Coleman, A.W. Portrait of a species: Chlamydomonas reinhardtii. Genetics 2005, 170, 1601–1610. [Google Scholar] [CrossRef] [Green Version]
- Merchant, S.S.; Prochnik, S.E.; Vallon, O.; Harris, E.H.; Karpowicz, S.J.; Witman , G.B.; Terry, A.; Salamov, A.; Fritz-Laylin, L.K.; Maréchal-Drouard, L.; et al. The Chlamydomonas genome reveals the evolution of key animal and plant functions. Science 2007, 318, 245–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rochaix, J.D. Chlamydomonas reinhardtii as the photosynthetic yeast. Annul. Rev. Genet. 1995, 29, 209–230. [Google Scholar] [CrossRef] [PubMed]
- Blaby, I.K.; Blaby-Haas, C.E.; Tourasse, N.; Hom, E.F.Y.; Lopez, D.; Aksoy, M.; Grossman, A.; Umen, J.; Dutcher, S.; Porter, M.; et al. The Chlamydomonas genome project: A decade on. Trends Plant Sci. 2014, 19, 672–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef] [PubMed]
- Bule, M.H.; Ahmed, I.; Maqbool, F.; Bilal, M.; Iqbal, H.M. Microalgae as a source of high- value bioactive compounds. Front. Biosci. 2018, 10, 197–216. [Google Scholar]
- Jayshree, A.; Jayashree, S.; Thangaraju, N. Chlorella vulgaris and Chlamydomonas reinhardtii: Effective antioxidant, antibacterial and anticancer mediators. Indian J. Pharm. Sci. 2016, 78, 575–581. [Google Scholar] [CrossRef] [Green Version]
- Annamalai, J.; Nallamuthu, T. Antioxidant potential phytochemicals from methanol extract of Chlorella vulgaris and Chlamydomonas reinhardtii. J. Algal Biomass Util 2014, 5, 60–67. [Google Scholar]
- Darwish, R.; Gedi, M.A.; Akepach, P.; Assaye, H.; Zaky, A.S.; Gray, D.A. Chlamydomonas reinhardtii Is a Potential Food Supplement with the Capacity to Outperform Chlorella and Spirulina. Appl. Sci. 2020, 10, 6736. [Google Scholar] [CrossRef]
- Scranton, M.A.; Ostrand, J.T.; Fields, F.J.; Mayfield, S.P. Chlamydomonas as a model for biofuels and bio-products production. Plant J. 2015, 82, 523–531. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Tan, Y.; Wang, W.; Bai, W.; Fan, J.; Huang, J.; Wan, M.; Li, Y. Efficient heterotrophic cultivation of Chlamydomonas Reinhardtii. J. Appl. Phycol. 2018, 31, 1545–1554. [Google Scholar] [CrossRef]
- Dreesen, I.A.; Charpin-El Hamri, G.; Fussenegger, M. Heat-stable oral alga-based vaccine protects mice from Staphylococcus aureus infection. J. Biotechnol. 2010, 145, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Gregory, J.A.; Topol, A.B.; Doerner, D.Z.; Mayfield, S.P. Alga-produced cholera toxin- Pfs25 fusion proteins as oral vaccines. Appl. Environ. Microbiol. 2013, 79, 3917–3925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres-Tiji, Y.; Fields, F.J.; Mayfield, S.P. Microalgae as a future food source. Biotechnol. Adv. 2020, 41, 107536. [Google Scholar] [CrossRef]
- Caporgno, M.P.; Mathys, A. Trends in microalgae incorporation into innovative food products with potential health benefits. Front. Nut. 2018, 5, 58. [Google Scholar] [CrossRef] [PubMed]
- Lordan, S.; Ross, R.P.; Stanton, C. Marine bioactives as functional food ingredients: Potential to reduce the incidence of chronic diseases. Mar. Drugs 2011, 9, 1056–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuño, K.; Villarruel-López, A.; Puebla-Pérez, A.M.; Romero-Velarde, E.; Puebla- Mora, A.G.; Ascencio, F. Effects of the marine microalgae Isochrysis galbana and Nannochloropsis oculata in diabetic rats. J. Funct. Foods 2013, 5, 106–115. [Google Scholar] [CrossRef]
- Deng, R.; Chow, T.J. Hypolipidemic, antioxidant, and antiinflammatory activities of microalgae spirulina. Cardiovasc. Ther. 2010, 28, e33–e45. [Google Scholar] [CrossRef] [Green Version]
- Fallah, A.A.; Sarmast, E.; Dehkordi, S.H.; Engardeh, J.; Mahmoodnia, L.; Khaledifar, A.; Jafari, T. Effect of Chlorella supplementation on cardiovascular risk factors: A meta-analysis of randomized controlled trials. Clin. Nutr. 2018, 37, 1892–1901. [Google Scholar] [CrossRef] [PubMed]
- Fields, F.J.; Lejzerowicz, F.; Schroeder, D.; Ngoi, S.M.; Tran, M.; McDonald, D.; Jiang, L.; Chang, J.T.; Knight, R.; Mayfield, S. Effects of the microalgae Chlamydomonas on gastrointestinal health. J. Funct. Foods 2020, 65, 103738. [Google Scholar] [CrossRef]
- Bhowmick, S.; Mazumdar, A.; Moulick, A.; Adam, V. Algal metabolites: An inevitable substitute for antibiotics. Biotechnol. Adv. 2020, 43, 107571. [Google Scholar] [CrossRef]
- Vishwakarma, J.; Vavilala, S.L. Evaluating the antibacterial and antibiofilm potential of sulphated polysaccharides extracted from green algae Chlamydomonas reinhardtii. J. Appl. Microbiol. 2019, 127, 1004–1017. [Google Scholar] [CrossRef] [PubMed]
- Kamble, P.; Cheriyamundath, S.; Lopus, M.; Sirisha, V.L. Chemical characteristics, antioxidant and anticancer potential of sulfated polysaccharides from Chlamydomonas reinhardtii. J. Appl. Phycol. 2018, 30, 1641–1653. [Google Scholar] [CrossRef]
- Mitchell, S.F.; Trainor, F.R.; Rich, P.H.; Goulden, C.E. Growth of Daphnia Magna in the Laboratory in Relation to the Nutritional State of Its Food Species, Chlamydomonas Reinhardtii. J. Plankton Res. 1992, 14, 379–391. [Google Scholar] [CrossRef]
- Davies, J.P.; Grossman, A.R. The use of Chlamydomonas (Chlorophyta: Volvocales) as a model algal system for genome studies and the elucidation of photosynthetic processes. J. Phycol. 1998, 34, 907–917. [Google Scholar] [CrossRef]
- Vukavic, T. Intestinal Absorption of IgA in the Newborn. J. Pediatr. Gastroenterol. Nutr. 1983, 2, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Liu, J.; Jiang, Y.; Chen, F. Chlorella Species as Hosts for Genetic Engineering and Expression of Heterologous Proteins: Progress, Challenge and Perspective. Biotechnol. J. 2016, 11, 1244–1261. [Google Scholar] [CrossRef] [PubMed]
- Lönnerdal, B. Infant Formula and Infant Nutrition: Bioactive Proteins of Human Milk and Implications for Composition of Infant Formulas. Am. J. Clin. Nutr. 2014, 99, 712S–717S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as Sources of Carotenoids. Mar. Drugs 2011, 9, 625–644. [Google Scholar] [CrossRef]
- Hou, Q.; Qiu, S.; Liu, Q.; Tian, J.; Hu, Z.; Ni, J. Selenoprotein-Transgenic Chlamydomonas Reinhardtii. Nutrition 2013, 5, 624–636. [Google Scholar] [CrossRef]
- Burgess, J.G. New and emerging analytical techniques for marine biotechnology. Curr. Opin. Biotechnol. 2012, 23, 29–33. [Google Scholar] [CrossRef]
- Bafana, A. Characterization and optimization of production of exopolysaccharide from Chlamydomonas reinhardtii. Carbohydr. Polym. 2013, 95, 746–752. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Montes, E. Polysaccharides: Sources, Characteristics, Properties, and Their Application in Biodegradable Films. Polysaccharides 2022, 3, 480–501. [Google Scholar] [CrossRef]
- Sousa, A.M.M.; Rocha, C.M.R.; Gonçalves, M.P. Agar. In Handbook of Hydrocolloids; Phillips, G.O., Williams, P.A., Eds.; Elsevier Ltd.: Amsterdam, The Netherlands, 2021; pp. 731–765. [Google Scholar]
- Morais, M.G.; Santos, T.D.; Moraes, L.; Vaz, B.S.; Morais, E.G.; Costa, J.A.V. Exopolysaccharides from microalgae: Production in a biorefinery framework and potential applications. Bioresour. Technol. Rep. 2022, 18, 101006. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, L.; Ren, Y.; Chen, F. Characterization of exopolysaccharides produced by microalgae with antitumor activity on human colon cancer cells. Int. J. Biol. Macromol 2019, 128, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M.A.; Beardall, J.; Raven, J.A. The Physiology of Microalgae, Developments in Applied Phycology 6; Springer International Publishing: Cham, Switzerland, 2016. [Google Scholar]
- Barclay, R.W.; Lewin, R.A. Microalgal polysaccharide production for the conditioning of agricultural soils. Plant Soil 1985, 88, 159–169. [Google Scholar] [CrossRef]
- Choi, S.P.; Nguyen, M.T.; Sim, S.J. Enzymatic pretreatment of Chlamydomonas reinhardtii biomass for ethanol production. Bioresour. Technol. 2010, 101, 5330–5336. [Google Scholar] [CrossRef]
- Ball, S.G.; Dirick, L.; Decq, A.; Martiat, J.C.; Matagne, R. Physiology of starch storage in the monocellular alga Chlamydomonas reinhardtii. Plant Sci 1990, 66, 1–9. [Google Scholar] [CrossRef]
- Melis, A. Photosynthetic H2 metabolism in Chlamydomonas reinhardtii (unicellular green algae). Planta 2007, 226, 1075–1086. [Google Scholar] [CrossRef]
- Raposo, M.F.J.; de Morais, A.M.B.; de Morais, R.M.S.C. Marine polysaccharides from algae with potential biomedical applications. Mar. Drugs 2015, 13, 2967–3028. [Google Scholar] [CrossRef] [Green Version]
- Wijesekara, I.; Pangestuti, R.; Kim, S.K. Biological activities and potential health benefits of sulphated polysaccharides derived from marine algae. Carbohyd. Polym 2011, 84, 14–21. [Google Scholar] [CrossRef]
- Costa, L.S.; Fidelis, G.P.; Cordeiro, S.L.; Oliveira, R.M.; Sabry, D.A.; Câmara, R.B.G.; Nobre, L.T.D.B.; Costa, M.S.S.P.; Almeida-Lima, J.; Farias, E.H.C.; et al. Biological activities of sulfated polysaccharides from tropical seaweeds. Biomed Pharm. 2010, 64, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Morais, A.M.M.B.; Alves, A.; Kumla, D.; Morais, R.M.S.C. Pharmaceutical and Biomedical Potential of Sulphated Polysaccharides from Algae. Polysaccharides of Microbial Origin; Oliveira, J.M., Radhouani, H., Reis, R.L., Eds.; Springer: Cham, Switzerland, 2021; pp. 1–28. [Google Scholar]
- Camara, R.B.; Day, L.S.; Fidelis, G.P.; Nobre, L.T.; Dantas-Santos, N.; Cordiro, S.L.; Costa, M.S.; Alves, L.G.; Rocha, H.A. Heterofucans from the brown seaweed Canistrocarpus cervicornis with anticoagulant and antioxidant activities. Mar. Drugs 2011, 24, 124–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, J.A.G.; Neto, E.M.; Teixeira, L.A.C.; Maula, P.R.C.; Mourao, P.A.S.; Benevides, N.M.B. Structural features and inactivation of coagulation proteases of a sulfated polysaccharidic fraction from Caulerpa cupressoides varlycopodium (Caulerpaceae, Chlorophyta.). Acta Sci. Technol. 2013, 35, 611–619. [Google Scholar] [CrossRef]
- Faggio, C.; Pagano, M.; Dottorem, A.; Genovese, G.; Morabito, M. Evaluation of anticoagulant activity of two algal polysaccharides. Nat. Prod. Res 2016, 17, 1934–1937. [Google Scholar] [CrossRef]
- Necas, J.; Bartosikova, L. Carrageenan: A review. Vet. Med. 2013, 58, 187–205. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, A.; Moghadamtousi, S.Z.; Abubakar, S.; Zandi, K. Antiviral potential of algae polysaccharides isolated from marine sources: A Review. Biomed Res. Int. 2015, 1, 825203. [Google Scholar] [CrossRef] [Green Version]
- Hu, T.D.; Liu, D.; Chen, Y.; Wu, J.; Wang, S. Antioxidant activity of sulfated polysaccharide fractions extracted from Undaria pinnitafida in vitro. Int. J. Biol. Macromol. 2010, 46, 193–198. [Google Scholar] [CrossRef]
- Costa, L.S.; Fidelis, G.P.; Telles, C.B.S.; Dantas-Santos, N.; Camara, R.B.G.S.; Cordeiro, L.; Pereira Costa, M.S.S.; Almeida-Lima, J.; Melo-Silveira, R.F.; Oliveira, R.M.; et al. Antioxidant and anti-proliferative activities of heterofucans from the seaweed Sargassum filipendula. Mar. Drugs 2011, 9, 952–966. [Google Scholar] [CrossRef] [Green Version]
- Souza, B.W.S.; Cerqueira, M.A.; Bourbon, A.I.; Pinheiro, A.C.; Martins, J.T.; Teixeira, J.A.; Coimbra, M.A.; Vicente, A.A. Chemical characterization and antioxidant activity of sulfated polysaccharide from the red seaweed Gracilaria birdiae. Food Hydrocoll. 2012, 27, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Hu, S.; Nie, S.; Yu, Q.; Xie, M. Reviews on mechanisms of In vitro antioxidant activity of polysaccharides. Oxid. Med. Cell. Longev. 2016, 64, 1–13. [Google Scholar]
- Luo, M.; Shao, B.; Nie, W.; Wei, X.W.; Li, Y.L.; Wang, B.L.; He, Z.Y.; Liang, X.; Ye, T.H.; Wei, Y.Q. Antitumor and adjuvant activity of λ-carrageenan by stimulating immune response in cancer immunotherapy. Sci.Rep. 2015, 5, 11062. [Google Scholar] [CrossRef] [PubMed]
- Lowenthal, R.M.; Fitton, J.H. Are seaweed-derived fucoidans possible future anti-cancer agents? J. Appl. Phycol. 2015, 27, 2075–2077. [Google Scholar] [CrossRef]
- Shao, P.; Chen, X.; Sun, P. In vitro antioxidant and antitumor activities of different sulfated polysaccharides isolated from three algae. Int. J. Biol. Macromol. 2013, 62, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Zaporozhets, T.S.; Ermakova, S.V.; Zvyagintseva, T.N.; Besednova, N.N. Antitumor effects of sulphated polysaccharides produced from marine algae. Biol. Bull. Rev 2014, 4, 122–132. [Google Scholar] [CrossRef]
- Rodrigues, J.A.G.; de Queiroz, I.N.L.; Gomes Quinderé, A.L.; Vairo, B.C.; de Souza Mourão, P.A.; Benevides, N.M.B. An antithrombin-dependent sulfated polysaccharide isolated from the green alga Caulerpa cupressoides has in vivo anti- and prothrombotic effects. Ciência Rural 2011, 41, 634–639. [Google Scholar] [CrossRef] [Green Version]
- Amorim, R.D.; Rodrigues, J.A.; Holanda, M.L.; Quinderé, A.L.; Paula, R.C.; Melo, V.M.; Benevides, N.M. Antimicrobial effect of a crude sulfated polysaccharide from the red seaweed Gracilaria ornate. Braz. Arch. Biol. Technol. 2012, 55, 171–181. [Google Scholar] [CrossRef]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Miyashita, K. Dietary combination of fucoxanthin and fish oil attenuates the weight gain of white adipose tissue and decreases blood glucose in obese/diabetic KK-Ay Mice. J. Agric. Food Chem. 2007, 55, 7701–7706. [Google Scholar] [CrossRef] [PubMed]
- Tsukui, T.; Konno, K.; Hosokawa, M.; Maeda, H.; Sashima, T.; Miyashita, K. Fucoxanthin and fucoxanthinol enhance the amount of docosahexaenoic acid in the liver of KKAy obese/diabetic mice. J. Agric. Food Chem. 2007, 55, 5025–5029. [Google Scholar] [CrossRef]
- Kong, C.S.; Kim, J.; Ahn, B.N.; Vo, T.S.; Yoon, N.Y.; Kim, S.K. 1-(3, 5-Dihydroxyphenoxy)-7-(2, 4, 6- trihydroxyphenoxy)-2, 4, 9-trihydroxydibenzo-1, 4-dioxin inhibits adipocyte differentiation of 3T3-L1 fibroblasts. Mar. Biotechnol. 2001, 12, 299–307. [Google Scholar] [CrossRef]
- Marques, C.T.; de Azevedo, T.C.G.; Nascimento, M.S.; Medeiros, V.P.; Alves, L.G.; Benevides, N.M.B.; Rocha, H.A.O.; Leite, E.L. Sulfated fucans extracted from algae Padina gymnospora have Anti-inflammatory effect. Rev. Bras. Farmacogn. 2012, 22, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Coura, C.O.; Souza, R.B.; Rodrigues, J.A.; Vanderlei, E.D.; de Araújo, I.W.; Ribeiro, N.A.; Frota, A.F.; Ribeiro, K.A.; Chaves, H.V.; Pereira, K.M.; et al. Mechanisms involved in the anti-inflammatory action of a polysulfated fraction from Gracilaria cornea in rats. PLoS ONE 2015, 10, e0119319. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, J.G.; Rodrigues, J.A.G.; de Sousa Oliveira Vanderlei, E.; Basto Souza, R.; Quinderé, A.L.G.; Coura, C.O.; de Araújo, I.W.F.; Chaves, H.V.; Bezerra, M.M.; Benevides, N.M.B. Peripheral Antinociception and Anti-Inflammatory Effects of Sulphated Polysaccharides from the Alga Caulerpa Mexicana. Basic Clin. Pharm. Toxicol. 2014, 115, 335–342. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Yang, Y.; Yang, G.; Yu, L.J. Studies on antibacterial activity and antibacterial mechanism of a novel polysaccharide from Streptomyces virginia H03. Food Control 2010, 21, 1257–1262. [Google Scholar] [CrossRef]
- Choudhary, S.; Save, S.N.; Vavilala, S.L. Unravelling the inhibitory activity of Chlamydomonas reinhardtii sulfated polysaccharides against α-Synuclein fibrillation. Sci. Rep. 2018, 8, 5692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somerville, C.; Browse, J.; Jaworski, J.; Ohlrogge, J. Lipids. In Biochemistry and Molecular Biology of Plants, 2nd ed.; Buchanan, B.B., Gruissem, W., Jones, R.L., Eds.; American Society of Plant Physiologists: Rockville, MD, USA, 2000. [Google Scholar]
- Lu, C.; Napier, J.A.; Clemente, T.E.; Cahoon, E.B. New frontiers in oilseed biotechnology: Meeting the global demand for vegetable oils for food, feed, biofuel, and industrial applications. Curr. Opin. Biotechnol. 2011, 22, 252–259. [Google Scholar] [CrossRef]
- Breuer, G.; Lamers, P.P.; Martens, D.E.; Draaisma, R.B.; Wijffels, R.H. The impact of nitrogen starvation on the dynamics of triacylglycerol accumulation in nine microalgae strains. Bioresour. Technol. 2012, 124, 217–226. [Google Scholar] [CrossRef]
- Affudeen, C.L.W.; Loh, S.H.; Aziz, A.; Takahashi, K.; Efendy, A.W.M.; Cha, T.S. Double-high in palmitic and oleic acids accumulation in a non-model green microalga, Messastrum gracile SE-MC4 under nitrate-repletion and -starvation cultivations. Sci. Rep. 2021, 11, 381. [Google Scholar] [CrossRef]
- Riediger, N.D.; Othman, R.A.; Suh, M.; Moghadasian, M.H. A systemic review of the roles of n-3 fatty acids in health and disease. J. Am. Diet. Assoc. 2009, 109, 668–679. [Google Scholar] [CrossRef]
- Khozin-Goldberg, I.; Leu, S.; Boussiba, S. Microalgae as a source for VLC-PUFA production. Lipids Plant Algae Dev. 2016, 86, 471–510. [Google Scholar]
- Calder, P.C. Omega-3 polyunsaturated fatty acids and inflammatory processes: Nutrition or pharmacology? Br. J. Clin. Pharmacol. 2013, 75, 645–662. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C. Polyunsaturated fatty acids and inflammatory processes: New twists in an old tale. Biochimie 2009, 91, 791–795. [Google Scholar] [CrossRef]
- Ferreri, C.; Chatgilialoglu, C. Membrane Lipidomics for Personalized Health; John Wiley & Sons.: Hoboken, NJ, USA, 2015; ISBN 978-1-118-54032-9. [Google Scholar]
- Shahidi, F.; Ambigaipalan, P. Omega-3 polyunsaturated fatty acids and their health benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 345–381. [Google Scholar] [CrossRef]
- Solovchenko, A.E. Physiological role of neutral lipid accumulation in eukaryotic microalgae under stresses. Russ. J. Plant Physiol. 2012, 59, 167–176. [Google Scholar] [CrossRef]
- Mulgund, A. Increasing Lipid Accumulation in Microalgae through Environmental Manipulation, Metabolic and Genetic Engineering: A Review in the Energy NEXUS framework. Energy Nexus 2022, 5, 100054. [Google Scholar] [CrossRef]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef] [Green Version]
- Li-Beisson, Y.; Beisson, F.; Riekhof, W. Metabolism of acyl-lipids in Chlamydomonas reinhardtii. Plant J. 2015, 82, 504–522. [Google Scholar] [CrossRef]
- Figueroa-Torres, G.M.; Pittman, J.K.; Theodoropoulos, C. Optimisation of Microalgal Cultivation via Nutrient-Enhanced Strategies: The Biorefinery Paradigm. Biotechnol. Biofuels 2021, 14, 1–16. [Google Scholar] [CrossRef]
- Ferreri, C.; Masi, A.; Sansone, A.; Giacometti, G.; Larocca, A.V.; Menounou, G.; Scanferlato, R.; Tortorella, S.; Rota, D.; Conti, M.; et al. Fatty acids in membranes as homeostatic, metabolic and nutritional biomarkers: Recent advancements in analytics and diagnostics. Diagnostics 2017, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Day, A.; Goldschmidt-Clermont, M. The chloroplast transformation toolbox: Selectable markers and marker removal. Plant Biotechnol. J. 2011, 9, 540–553. [Google Scholar] [CrossRef] [Green Version]
- Michelet, L.; Lefebvre-Legendre, L.; Burr, S.E.; Rochaix, J.D.; Goldschmidt-Clermont, M. Enhanced chloroplast transgene expression in a nuclear mutant of Chlamydomonas. Plant Biotechnol. J. 2011, 9, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Rochaix, J.D. The three genomes of Chlamydomonas. In Discoveries in Photosynthesis, Govindjee; Beatty, J.T., Gest, H., Allen, J.F., Eds.; Springer: Dordrecht, The Netherlands, 2005; Volume 20, pp. 1047–1055. [Google Scholar]
- Borowiak, D.; Krzywonos, M. Bioenergy, biofuels, lipids and pigments—Research trends in the use of microalgae grown in photobioreactors. Energies 2022, 15, 5357. [Google Scholar] [CrossRef]
- De Luca, M.; Pappalardo, I.; Limongi, A.R.; Viviano, E.; Radice, R.P.; Todisco, S.; Martelli, G.; Infantino, V.; Vassallo, A. Lipids from microalgae for cosmetic applications. Cosmetics 2021, 8, 52. [Google Scholar] [CrossRef]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The promising future of microalgae: Current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Factories 2018, 17, 36. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.P.; Wang, M.; Wang, C. Nuclear transformation of Chlamydomonas reinhardtii: A review. Biochimie 2021, 181, 1–11. [Google Scholar] [CrossRef]
- Cutolo, E.A.; Mandalà, G.; Dall’Osto, L.; Bassi, R. Harnessing the Algal Chloroplast for Heterologous Protein Production. Microorganisms 2022, 10, 743. [Google Scholar] [CrossRef]
- Braun-Galleani, S.; Baganz, F.; Purton, S. Improving recombinant protein production in the Chlamydomonas reinhardtii chloroplast using vivid Verde Fluorescent Protein as a reporter. Biotechnol. J. 2015, 10, 1289–1297. [Google Scholar] [CrossRef] [Green Version]
- Tran, M.; Zhou, B.; Pettersson, P.L.; Gonzalez, M.J.; Mayfield, S.P. Synthesis and assembly of a full-length human monoclonal antibody in algal chloroplasts. Biotechnol. Bioeng. 2009, 104, 663–673. [Google Scholar] [CrossRef]
- Tran, M.; Van, C.; Barrera, D.J.; Pettersson, P.L.; Peinado, C.D.; Bui, J.; Mayfield, S.P. Production of unique immunotoxin cancer therapeutics in algal chloroplasts. Proc. Natl. Acad. Sci. USA 2013, 110, E15–E22. [Google Scholar] [CrossRef] [Green Version]
- Chávez, M.N.; Schenck, T.L.; Hopfner, U.; Centeno-Cerdas, C.; Somlai-Schweiger, I.; Schwarz, C.; Machens, H.G.; Heikenwalder, M.; Bono, M.R.; Allende, M.L.; et al. Towards autotrophic tissue engineering: Photosynthetic gene therapy for regeneration. Biomaterials 2016, 75, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Schroda, M. The Chlamydomonas genome reveals its secrets: Chaperone genes and the potential roles of their gene products in the chloroplast. Photosynth. Res. 2004, 82, 221–240. [Google Scholar] [CrossRef]
- Breiman, A.; Fawcett, T.W.; Ghirardi, M.L.; Mattoo, A.K. Plant organelles contain distinct peptidylprolyl cis, trans-isomerases. J. Biol. Chem. 1992, 267, 21293–21296. [Google Scholar] [CrossRef] [PubMed]
- Ramana, K.V.; Xavier, J.R.; Sharma, R.K. Recent trends in pharmaceutical biotechnology. Pharm. Biotechnol. Curr. Res. 2017, 1, 5. [Google Scholar]
- Yan, N.; Fan, C.; Chen, Y.; Hu, Z. The potential for microalgae as bioreactors to produce pharmaceuticals. Int. J. Mol. Sci. 2016, 17, 962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacheco, S.E.C.; Hankamer, B.; Oey, M. Optimising light conditions increases recombinant protein production in Chlamydomonas reinhardtii chloroplasts. Algal Res. 2018, 32, 329–340. [Google Scholar] [CrossRef] [Green Version]
- Stoffels, L.; Finlan, A.; Mannall, G.; Purton, S.; Parker, B. Downstream processing of Chlamydomonas reinhardtii TN72 for recombinant protein recovery. Front. Bioeng. Biotechnol. 2019, 7, 383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almaraz-Delgado, A.L.; Flores-Uribe, J.; Pérez-España, V.H.; Salgado-Manjarrez, E.; Badillo-Corona, J.A. Production of therapeutic proteins in the chloroplast of Chlamydomonas reinhardtii. AMB Express 2014, 4, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.; Qian, K.; Su, N.; Chang, H.; Liu, J.; Shen, G. Foot-and-mouth disease virus VP1 protein fused with cholera toxin B subunit expressed in Chlamydomonas reinhardtii chloroplast. Biotechnol. Lett. 2003, 25, 1087–1092. [Google Scholar] [CrossRef]
- Mayfield, S.P.; Franklin, S.E.; Lerner, R.A. Expression and assembly of a fully active antibody in algae. Proc. Natl. Acad. Sci. USA 2003, 100, 438–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Chen, F.; Li, D.; Zhang, Z.; Liu, Y.; Zheng, D.; Liu Yanxin, L.; Dexian, Z.; Yong, W.; Shen, G. Expression of human soluble TRAIL in Chlamydomonas reinhardtii chloroplast. Chin. Sci. Bull. 2006, 51, 1703–1709. [Google Scholar] [CrossRef]
- Manuell, A.L.; Beligni, M.V.; Elder, J.H.; Siefker, D.T.; Tran, M.; Weber, A.; McDonal, T.L.; Mayfield, S.P. Robust expression of a bioactive mammalian protein in Chlamydomonas chloroplast. Plant Biotechnol. J. 2007, 5, 402–412. [Google Scholar] [CrossRef] [PubMed]
- He, D.M.; Qian, K.X.; Shen, G.F.; Zhang, Z.F.; Li, Y.N.; Su, Z.L.; Shao, H.B. Recombination and expression of classical swine fever virus (CSFV) structural protein E2 gene in Chlamydomonas reinhardtii chroloplasts. Colloids Surf. B Biointerfaces 2007, 55, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Brandsma, M.; Tremblay, R.; Maxwell, D.; Jevnikar, A.M.; Huner, N.; Ma, S. A novel expression platform for the production of diabetes-associated autoantigen human glutamic acid decarboxylase (hGAD65). BMC Biotechnol. 2008, 8, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surzycki, R.; Greenham, K.; Kitayama, K.; Dibal, F.; Wagner, R.; Rochaix, J.D.; Ajam, T.; Surzycki, S. Factors effecting expression of vaccines in microalgae. Biologicals 2009, 37, 133–138. [Google Scholar] [CrossRef]
- Rasala, B.A.; Muto, M.; Lee, P.A.; Jager, M.; Cardoso, R.M.; Behnke, C.A.; Kirk, C.A.; Hokanson, R.; Crea, M.; Mendez, S.P.; et al. Production of therapeutic proteins in algae, analysis of expression of seven human proteins in the chloroplast of Chlamydomonas reinhardtii. Plant Biotechnol. J. 2010, 8, 719–733. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.M.; Kim, S.Y.; Li, K.F.; Yoon, B.H.; Choe, S.; Kuo, M.M.C. Transgenic microalgae expressing Escherichia coli AppA phytase as feed additive to reduce phytate excretion in the manure of young broiler chicks. Appl. Microbiol. Biot. 2011, 91, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Gregory, J.A.; Li, F.; Tomosada, L.M.; Cox, C.J.; Topol, A.B.; Vinetz, J.M.; Mayfield, S. Algae-produced Pfs25 elicits antibodies that inhibit malaria transmission. PLoS ONE 2012, 7, e37179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, M.; Henry, R.E.; Siefker, D.; Van, C.; Newkirk, G.; Kim, J.; Bui, J.; Mayfield, S.P.M.; Tran, R.E. Production of anti-cancer immunotoxins in algae: Ribosome inactivating proteins as fusion partners. Biotechnol. Bioeng. 2013, 110, 2826–2835. [Google Scholar] [CrossRef]
- Rasala, B.A.; Mayfield, S.P. The microalga Chlamydomonas reinhardtii as a platform for the production of human protein therapeutics. Bioeng. Bugs 2011, 2, 50–54. [Google Scholar] [CrossRef] [Green Version]
- Pang, X.; Tong, Y.; Xue, W.; Yang, Y.F.; Chen, X.; Liu, J.; Chen, D. Expression and characterization of recombinant human lactoferrin in edible alga Chlamydomonas reinhardtii. Biosci. Biotechnol. Biochem. 2019, 83, 851–859. [Google Scholar] [CrossRef]
- Grossman, A.R.; Lohr, M.; Im, C.S. Chlamydomonas reinhardtii in the landscape of pigments. Annu. Rev. Genet. 2004, 38, 119–173. [Google Scholar] [CrossRef]
- Koyande, A.K.; Chew, K.W.; Rambabu, K.; Tao, Y.; Chu, D.T.; Show, P.L. Microalgae: A potential alternative to health supplementation for humans. Food Sci. Hum. Wellness 2019, 8, 16–24. [Google Scholar] [CrossRef]
- Gille, A.; Trautmann, A.; Posten, C.; Briviba, K. Bioaccessibility of carotenoids from Chlorella vulgaris and Chlamydomonas reinhardtii. Int. J. Food Sci. Nutr 2016, 67, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Rathod, J.P.; Vira, C.; Lali, A.M.; Prakash, G. Metabolic engineering of Chlamydomonas reinhardtii for enhanced β-carotene and lutein production. Appl. Biochem. Biotechnol. 2010, 190, 1457–1469. [Google Scholar] [CrossRef] [PubMed]
- Song, I.; Kim, J.; Baek, K.; Choi, Y.; Shin, B.; Jin, E. The generation of metabolic changes for the production of high-purity zeaxanthin mediated by CRISPR-Cas9 in Chlamydomonas reinhardtii. Microb. Cell Factories 2020, 19, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cordero, B.F.; Couso, I.; León, R.; Rodríguez, H.; Vargas, M.A. Enhancement of carotenoids biosynthesis in Chlamydomonas reinhardtii by nuclear transformation using a phytoene synthase gene isolated from Chlorella zofingiensis. Appl. Microbiol. Biotechnol. 2011, 91, 341–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Ma, R.; Liu, X.; Ho, S.H.; Xie, Y.; Chen, J. Strategies related to light quality and temperature to improve lutein production of marine microalga Chlamydomonas sp. Bioprocess Biosyst. Eng. 2019, 42, 435–443. [Google Scholar] [CrossRef]
- Tran, Q.G.; Cho, K.; Kim, U.; Yun, J.H.; Cho, D.H.; Heo, J.; Park, S.B.; Kim, J.W.; Lee, Y.J.; Ramanan, R.; et al. Enhancement of β-carotene production by regulating the autophagy- carotenoid biosynthesis seesaw in Chlamydomonas reinhardtii. Bioresour. Technol. 2019, 292, 121937. [Google Scholar] [CrossRef]
- Scott, J.D.; Chalker-Scott, L.; Foreman, A.E.; D’Angelo, M. Daphnia Pulex Fed UVB- Irradiated Chlamydomonas Reinhardtii Show Decreased Survival and Fecundity. Photochem. Photobioliol. 1999, 70, 308–313. [Google Scholar] [CrossRef]
- Gophen, M. Feeding of Daphnia on Chlamydomonas and Chlorobium. Nature 1977, 265, 271–273. [Google Scholar] [CrossRef]
- Taub, F.B.; Dollar, A.M. The nutritional inadequacy of Chlorella and Chlamydomonas as food for Daphnia Pulex. Limnol. Oceanogr. 1968, 13, 607–617. [Google Scholar] [CrossRef]
- Weers, P.M.M.; Gulati, R.D. Gulati, Growth and Reproduction of Daphnia Galeata in Response to Changes in Fatty Acids, Phosphorus, and Nitrogen in Chlamydomonas Reinhardtii. Limnol. Oceanogr. 1997, 42, 1584–1589. [Google Scholar] [CrossRef] [Green Version]
- DeMott, W.R. Feeding Selectivities and Relative Ingestion Rates of Daphnia and Bosmina. Limnol. Oceanogr. 1982, 27, 518–527. [Google Scholar] [CrossRef]
- Triton Algae Innovations Inc. Notice to US Food and Drug Administration of the Conclusion that the Intended Use of Chlamydomonas reinhardtii (THN 6) Dried Biomass Powder is Generally Recognized as Safe. Available online: https://www.fda.gov/media/128921/download (accessed on 1 December 2022).
- Murbach, T.S.; Glávits, R.; Endres, J.R.; Hirka, G.; Vértesi, A.; Béres, E.; Szakonyiné, I.P. Toxicological Evaluation of Chlamydomonas Reinhardtii, a Green Algae. Int. J. Toxicol. 2018, 37, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Becker, E.W. Micro-Algae as a Source of Protein. Biotechnol. Adv. 2007, 25, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Boyle, N.R.; Morgan, J.A. Flux Balance Analysis of Primary Metabolism in Chlamydomonas Reinhardtii. BMC Syst. Biol. 2009, 3, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, M.J.; Harrison, S.T.L. Lipid Productivity as a Key Characteristic for Choosing Algal Species for Biodiesel Production. J. Appl. Phycol. 2008, 21, 493–507. [Google Scholar] [CrossRef]
- Knap, A.H.; Michaels, A.; Close, A.R.; Ducklow, H.; Dickson, A.G. Protocols for the Joint Global Ocean Flux Study (JGOFS) Core Measurements. Intergovernmental Oceanographic Commission; (Intergovernmental Oceanographic Commission Manuals and Guides: 29), (JGOFS Report; 19); UNESCO-IOC: Paris, France, 1994; 170p. [Google Scholar]
- Baosheng, G.E.; Xiangfa, W.; Fang, H.; Jie, L. Method for Overproducing Hemoglobin in Algae and Compositions Therefrom. Chinese Patent CN111213579A, 02 February 2019. [Google Scholar]
- Wang, X.; Innovations, T.A. Green Algae as a Platform for Protein Production: Food, Feed, and Nutritional Supplements; Montreal, QC, Canada, 2017. Available online: https://www.bio.org/sites/default/files/legacy/bioorg/docs/0830AM-Xun%20Wang.pdf (accessed on 1 December 2022).
- Tran, M.; Deaton, J.; Adams, B.; Mayfield, M.; Longo, A.; Gonzalez, O.; Hansen, J.; Wang, X.; Schroeder, D. Compositions and Methods for Incorporating Heme from Algae in Edible Products. U.S. Patent 20210401008A1, 30 December 2021. [Google Scholar]
- Tran, M.; Deaton, J.; Adams, B.; Mayfield, M.; Longo, A.; Gonzalez, O.; Hansen, J.; Wang, X.; Schroeder, D. Method of Purification of Recombinant Osteopontin from Microalgae. U.S. Patent 20190276508A1, 12 September 2019. [Google Scholar]
- Tran, M.; Deaton, J.; Adams, B.; Mayfield, M.; Longo, A.; Gonzalez, O.; Hansen, J.; Wang, X.; Schroeder, D. Methods for Overproducing Protoporphyrin Ix in Algae and Compositions Therefrom. U.S. Patent 20210386088 A1, 16 December 2021. [Google Scholar]
- Diego, S.; Adams, B.; Diego, S.; Deaton, J.; Diego, S.; Hansen, J.; Diego, S.; Longo, A.; Diego, S.; Mayfield, M.; et al. Production of Iron—Complexed Proteins. U.S. Patent 20200332249 A1, 22 October 2020. [Google Scholar]
- Peña, D.A.; Gasser, B.; Zanghellini, J.; Steiger, M.G.; Mattanovich, D. Metabolic Engineering of Pichia Pastoris. Metab. Eng. 2018, 50, 2–15. [Google Scholar] [CrossRef]
- Fraser, R.Z.; Shitut, M.; Agrawal, P.; Mendes, O.; Klapholz, S. Safety Evaluation of Soy Leghemoglobin Protein Preparation Derived From Pichia Pastoris, Intended for Use as a Flavor Catalyst in Plant-Based Meat. Int. J. Toxicol. 2018, 37, 241–262. [Google Scholar] [CrossRef] [Green Version]
- Sproles, A.E.; Fields, F.J.; Smalley, T.N.; Le, C.H.; Badary, A.; Mayfield, S.P. Recent advancements in the genetic engineering of microalgae. Algal Res. 2021, 53, 102158. [Google Scholar] [CrossRef]
- Crozet, P.; Navarro, F.J.; Willmund, F.; Mehrshahi, P.; Bakowski, K.; Lauersen, K.J.; Pérez-Pérez, M.-E.; Auroy, P.; Gorchs Rovira, A.; Sauret-Gueto, S.; et al. Birth of a photosynthetic chassis: A MoClo toolkit enabling synthetic biology in the microalga Chlamydomonas reinhardtii. ACS Synth. Biol. 2018, 7, 2074–2086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiner, I.; Atar, S.; Schweitzer, S.; Eilenberg, H.; Feldman, Y.; Avitan, M.; Blau, M.; Danon, A.; Tuller, T.; Yacoby, I. Enhancing heterologous expression in Chlamydomonas reinhardtii by transcript sequence optimization. Plant J. 2018, 94, 22–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Martinez, E.M.; Fimognari, L.; Sakuragi, Y. High-yield secretion of recombinant proteins from the microalga Chlamydomonas reinhardtii. Plant Biotechnol. J. 2017, 15, 1214–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasala, B.A.; Mayfield, S.P. Photosynthetic biomanufacturing in green algae; production of recombinant proteins for industrial, nutritional, and medical uses. Photosynth. Res. 2015, 123, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Schroda, M.; Blöcker, D.; Beck, C.F. The HSP70A promoter as a tool for the improved expression of transgenes in Chlamydomonas. Plant J. 2000, 21, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Perozeni, F.; Stella, G.R.; Ballottari, M. LHCSR expression under HSP70/RBCS2 promoter as a strategy to increase productivity in microalgae. Int. J. Mol. Sci. 2018, 19, 155. [Google Scholar] [CrossRef] [Green Version]
- Xue, B.; Dong, C.M.; Hu, H.H.; Dong, B.; Fan, Z.C. Chlamydomonas reinhardtii-expressed multimer of ToAMP4 inhibits the growth of bacteria of both Gram-positive and Gram- negative. Process Biochem. 2020, 91, 311–318. [Google Scholar] [CrossRef]
- Kwon, K.C.; Lamb, A.; Fox, D.; Jegathese, S.J.P. An evaluation of microalgae as a recombinant protein oral delivery platform for fish using green fluorescent protein (GFP). Fish Shellfish Immunol. 2019, 87, 414–420. [Google Scholar] [CrossRef]
- Shamriz, S.; Ofoghi, H. Expression of recombinant PfCelTOS antigen in the chloroplast of Chlamydomonas reinhardtii and its potential use in detection of malaria. Mol. Biotechnol. 2019, 61, 102–110. [Google Scholar] [CrossRef]
- Rosales-Mendoza, S.; Solís-Andrade, K.I.; Márquez-Escobar, V.A.; González-Ortega, O.; Bañuelos-Hernandez, B. Current advances in the algae-made biopharmaceuticals field. Expert Opin. Biol. Ther. 2020, 20, 751–766. [Google Scholar] [CrossRef]
- Griesbeck, C.; Kobl, I.; Heitzer, M. Chlamydomonas reinhardtii. Mol. Biotechnol. 2006, 34, 213–223. [Google Scholar] [CrossRef]
- Perozeni, F.; Cazzaniga, S.; Baier, T.; Zanoni, F.; Zoccatelli, G.; Lauersen, K.J.; Wobbe, L.; Ballottari, M. Turning a green alga red: Engineering astaxanthin biosynthesis by intragenic pseudogene revival in Chlamydomonas reinhardtii. Plant Biotechnol. J. 2020, 18, 2053–2067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F., III. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Antonacci, A.; Scognamiglio, V. Biotechnological advances in the design of algae-based biosensors. Trends Biotechnol. 2020, 38, 334–347. [Google Scholar] [CrossRef] [PubMed]
- Ghribi, M.; Nouemssi, S.B.; Meddeb-Mouelhi, F.; Desgagné-Penix, I. Genome editing by CRISPR-Cas: A game change in the genetic manipulation of Chlamydomonas. Life 2020, 10, 295. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.E.; Lim, J.M.; Koh, H.G.; Kim, E.K.; Kang, N.K.; Jeon, S.; Kwon, S.; Shin, W.S.; Lee, B.; Hwangbo, K.; et al. CRISPR/Cas9-induced knockout and knock-in mutations in Chlamydomonas reinhardtii. Sci. Rep. 2016, 6, 27810. [Google Scholar] [CrossRef] [PubMed]
- Baek, K.; Yu, J.; Jeong, J.; Sim, S.J.; Bae, S.; Jin, E. Photoautotrophic production of macular pigment in a Chlamydomonas reinhardtii strain generated by using DNA-free CRISPR-Cas9 RNP-mediated mutagenesis. Biotechnol. Bioeng. 2018, 115, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Rasala, B.A.; Chao, S.S.; Pier, M.; Barrera, D.J.; Mayfield, S.P. Enhanced Genetic Tools for Engineering Multigene Traits into Green Algae. PLoS ONE 2014, 9, e94028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scaife, M.A.; Nguyen, G.T.D.T.; Rico, J.; Lambert, D.; Helliwell, K.E.; Smith, A.G. Establishing Chlamydomonas Reinhardtii as an Industrial Biotechnology Host. Plant J. 2015, 82, 532–546. [Google Scholar] [CrossRef]
- Giordano, M.; Wang, Q. Microalgae for Industrial Purposes. Biomass and Green Chemistry; Chapter 6; Springer: Berlin/Heidelberg, Switzerland, 2017; ISBN 978-3-319-66735-5. [Google Scholar]
- Anderson, M.S.; Muff, T.J.; Georgianna, D.R.; Mayfield, S.P. Towards a Synthetic Nuclear Transcription System in Green Algae: Characterization of Chlamydomonas Reinhardtii Nuclear Transcription Factors and Identification of Targeted Promoters. Algal Res. 2017, 22, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Barrera, D.J.; Mayfield, S.P. High-Value Recombinant Protein Production in Microalgae. In Handbook of Microalgal Culture: Biotechnology and Applied Phycology, 2nd ed.; Wiley & Sons: Oxford, UK,, 2013; pp. 532–544. ISBN 9781118567166. [Google Scholar]
- Fields, F.J.; Ostrand, J.T.; Mayfield, S.P. Fed-Batch Mixotrophic Cultivation of Chlamydomonas Reinhardtii for High-Density Cultures. Algal Res. 2018, 33, 109–117. [Google Scholar] [CrossRef]
- Mayfield, S.P.; Manuell, A.L.; Chen, S.; Wu, J.; Tran, M.; Siefker, D.; Muto, M.; Marin-Navarro, J. Chlamydomonas Reinhardtii Chloroplasts as Protein Factories. Curr. Opin. Biotechnol. 2007, 18, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Franklin, S.; Ngo, B.; Efuet, E.; Mayfield, S.P.; Franklin, S.; Ngo, B.; Efuet, E.; Mayfield, S.P. Development of a GFP Reporter Gene for Chlamydomonas Reinhardtii Chloroplast. Plant J. 2002, 30, 733–744. [Google Scholar] [CrossRef] [Green Version]
- Loera-Quezada, M.M.; Leyva-González, M.A.; Velázquez-Juárez, G.; Sanchez-Calderón, L.; Do Nascimento, M.; López-Arredondo, D.; Herrera-Estrella, L. A Novel Genetic Engineering Platform for the Effective Management of Biological Contaminants for the Production of Microalgae. Plant Biotechnol. J. 2016, 14, 2066–2076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takouridis, S.J.; Tribe, D.E.; Gras, S.L.; Martin, G.J.O. The Selective Breeding of the Freshwater Microalga Chlamydomonas Reinhardtii for Growth in Salinity. Bioresour. Technol. 2015, 184, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Li, L.Z.S.; Chang, S.; Wang, W.; Zhang, Z.; Wang, J.; Zhao, G.; Qi, L.; Xu, W. Characterization of Cell Growth and Photobiological H2 Production of Chlamydomonas Reinhardtii in ASSF Industry Wastewater. Int. J. Hydrogen Energy 2014, 39, 13462–13467. [Google Scholar] [CrossRef]
- Tran, M.; Chang, J.T.; Hansen, J.; Mayfield, M.; Mayfield, S.P.; Rasala, B.A.; Wang, X. Inventors; Triton Algae Innovations, Assignee. Method of Treating Alimentary Canal Conditions. Method of Treating Alimentary Canal Conditions. U.S. Patent 16/609,398, 18 June 2020. [Google Scholar]
- Koller, M.; Muhr, A.; Braunegg, G. Microalgae as Versatile Cellular Factories for Valued Products. Algal Res. 2014, 6, 52–63. [Google Scholar] [CrossRef]
- Pulz, O.; Gross, W. Valuable Products from Biotechnology of Microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef]
- Gunasekaran, B.; Gothandam, K.M. A review on edible vaccines and their prospects. Braz. J. Med. Biol. Res. 2020, 53, e8749. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sugar Monomers (in Alphabetical Order) | Ref. |
|---|---|---|
| C. mexicana (or C. oblonga) 1 | Ara, Fuc,2 Gal, Glu,2 Man, Rha, Rib, Xyl, UA | [49] |
| C. reinhardtii | Ara, Gal, Glu,2 Man, Rha | [50] |
| C. sajao (or Labochamys segnis) 1,3 | Ara, Gal,2 Glu, Man, Rha, Xyl, UA | [49] |
| FA (DW) | C. reinhardtii | Chlorella | Spirulina | |||
|---|---|---|---|---|---|---|
| mg/g- | % | mg/g- | % | mg/g- | % | |
| C16:0 | 16.7± 0.8 | 23.8 ± 0.2 | 11.0 ± 0.3 | 22.2 ± 0.1 | 25.9 ± 4.6 | 57.9 ± 0.4 |
| C16:1 n-7 | 1.9 ± 0.1 | 2.7 ± 0.1 | 6.5 ± 0.1 | 13.0 ± 0.1 | 0.1 ± 0.0 | 0.1 ± 0.0 |
| C16:4 n-3 | 3.9 ± 0.1 | 5.5 ± 0.3 | 0.0 ± 0.0 | 0.1 ± 0.0 | 0.0 ± 0.0 | 0.1 ± 0.0 |
| C18:0 | 1.6 ± 0.2 | 2.3 ± 0.3 | 1.4 ± 0.0 | 2.8 ± 0.1 | 0.7± 0.0 | 1.5 ± 0.1 |
| C18:1 n-9c | 10.3 ± 1.1 | 14.7 ± 1.6 | 3.5 ± 0.1 | 7.0 ± 0.2 | 0.9 ± 0.2 | 2.0 ± 0.2 |
| C18:2 n-6c | 2.7 ± 0.2 | 3.8 ± 0.2 | 15.6 ± 0.3 | 31.4 ± 0.3 | 8.5 ± 1.6 | 19.0 ± 0.2 |
| C18:3 n-6 (GLA) | 2.9 ± 0.2 | 4.1 ± 0.3 | 0.0 ± 0.0 | 0.1 ± 0.0 | 8.7 ± 1.6 | 19.5 ± 0.2 |
| C18:3 n-3 | 29.8 ± 1.9 | 42.4 ± 1.2 | 11.6 ± 0.2 | 23.4 ± 0.1 | 0.1 ± 0.0 | 0.1 ± 0.0 |
| C20:4 n-6 | 0.6 ± 0.1 | 0.9 ± 0.0 | 0.0 ± 0.0 | 0.1 ± 0.0 | 8.7 ± 1.6 | 19.5 ± 0.2 |
| ƩSFA | 18.3 | 26.02 | 12.4 | 25.0 | 26.6 | 59.4 |
| ƩUSFA | 52.1 | 74.0 | 37.3 | 75.0 | 18.5 | 40.6 |
| Ʃn-3 FA | 33.7 | 47.9 | 11.6 | 23.4 | 0.1 | 0.2 |
| Ʃn-6 FA | 6.2 | 8.7 | 15.7 | 31.6 | 17.2 | 38.3 |
| n-6/n-3 | 0.2 | 0.2 | 1.4 | 1.4 | und | und |
| Recombinant Therapeutic Protein | Yield | Ref. |
|---|---|---|
| VP1-CTB; protein VP1 from the foot and mouth disease virus (FMDV) fused to cholera toxin B (CTB) | 3–4% total soluble protein (TSP) | [115] |
| HSV-lsc; large single chain (lsc) antibody directed against glycoprotein D protein from the herpes simplex virus (HSV) | Not reported | [116] |
| TRAIL; tumor necrosis factor-related apoptosis-inducing ligand | 0.43–0.67% TSP | [117] |
| M-SAA; mammary-associated serum amyloid | 3–5% TSP | [118] |
| CSFV-E2; classical swine fever virus (CSFV) structural protein E2 | 1.5–2% TSP | [119] |
| Human glutamic acid decarboxylase (hGAD65) | 0.25–0.3% TSP | [120] |
| IBDV-VP2; infectious burial disease virus VP2 protein | 4–0.8% total cell protein (TCP) | [121] |
| IHNV-G; infectious hematopoietic necrosis virus | <0.5% TCP | |
| IPNV-VP2; infectious pancreatic necrosis virus | <0.3% TCP | |
| VP2 protein | 1–0.1% TCP | |
| IPNV-VP2 SBC; infectious pancreatic necrosis virus | 1–0.2% TCP | |
| Quorum sensing-regulated gene (LecA) p57 | <0.5 TCP | |
| PCV2; porcine circovirus type 2 | 0.9–0.2% TCP | |
| VP-2C | <0.5% TCP | |
| VP28 | 21–0.2% TCP | |
| HC-83K7C; heavy chain human monoclonal antibody against anthrax protective antigen 83 (PA83) | 0.01% dry weight | [105] |
| LC-83K7C; light chain human monoclonal antibody against anthrax PA83 | ||
| CTB-D2; D2 fibronectin-binding domain of staphylococcus aureus fused to the cholera toxin B subunit | 0.7% TSP | [23] |
| 14FN3; domain 14 of human fibronectin | 3–0.15% TSP | [122] |
| VEGF; human vascular endothelial growth factor | 2–0.1% TSP | |
| HMGB1; high mobility group protein B1 | 2.5–1% TSP | |
| acrV2 and vapA2; antigens from the fish pathogen aeromonas salmonicida | 0.8% and 0.3% TSPrespectively | [97] |
| Escherichia coli phytase gene (appA) | Not Detected | [123] |
| Pfs25 and Pfs28; surface proteins from plasmodium falciparum | 0.5% and 0.2%TSP respectively | [124] |
| αCD22PE40; monomeric immunotoxin consisting of the single chain antibody that recognizes the CD22 surface protein from B- cells, fused to domains II and III of exotoxin A (PE40) from pseudomonas aeruginosa | 0.3–0.4% TSP | [106] |
| αCD22HCH23PE40; dimeric version of αCD22PE40 | 0.2–0.3% TSP | |
| CtxB-Pfs25; plasmodium falciparum surface protein 25 fused to the β subunit of the cholera toxin from vibrio cholera | 0.09% TSP | [24] |
| αCD22Gel; single-chain antibody targeting the CD22 receptor from B-cells, fused to the eukaryotic ribosome-inactivating protein, gelonin, from gelonium multiflorm | 0.2–0.3% TSP | [112] |
| αCD22CH23Gel; dimeric version of αCD22Gel | 0.1–0.2% TSP |
| Physical Characteristics | Specification | Batch TAI-1215-01 | Batch TAI-0316-01 | Method |
|---|---|---|---|---|
| Appearance | Green powder | Green powder | a | Visual inspection |
| Water content | ≤10% | 5.6% | 15.57% | AOAC Variable |
| Composition | ||||
| Protein | 30–70% | 36.0% | 39.9% | AOAC 990.03 |
| Fat | ≤10% | 2.0% | 4.23% | AOAC 945.16 |
| Fiber | 1–25% | 7.3% | 3.6% | AOAC 991.43 |
| Ash | ≤5% | 4.8% | 1.87% | AOAC 942.05 |
| Chlorophyll | ≤25% | 0.49% | a | [146] |
| Heavy metals | ||||
| Arsenic, cadmium, lead, Mercury | ≤0.2 ppm | <0.1 ppm, 0.1 ppm, <0.1 ppm, <0.1 ppm | a | USP<233>, EPA 7471 |
| Microbiological tests | ||||
| Total aerobic microbial | ≤1000 CFU/g | ND | a | AOAC 990.12 |
| Total yeast and mold | ≤1000 CFU/g | 190 CFU/g | a | BAM Ch. 18 |
| Total coliforms | ≤100 CFU/g | ND | a | AOAC 991.14 |
| E. coli | Negative (absent/1 g) | Negative | a | AOAC 991.14 |
| Salmonella | Negative (absent/25 g) | Negative | a | AOAC 030301 |
| Staphylococcus | Negative (absent/1 g) | Negative | a | AOAC 2003.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masi, A.; Leonelli, F.; Scognamiglio, V.; Gasperuzzo, G.; Antonacci, A.; Terzidis, M.A. Chlamydomonas reinhardtii: A Factory of Nutraceutical and Food Supplements for Human Health. Molecules 2023, 28, 1185. https://doi.org/10.3390/molecules28031185
Masi A, Leonelli F, Scognamiglio V, Gasperuzzo G, Antonacci A, Terzidis MA. Chlamydomonas reinhardtii: A Factory of Nutraceutical and Food Supplements for Human Health. Molecules. 2023; 28(3):1185. https://doi.org/10.3390/molecules28031185
Chicago/Turabian StyleMasi, Annalisa, Francesca Leonelli, Viviana Scognamiglio, Giulia Gasperuzzo, Amina Antonacci, and Michael A. Terzidis. 2023. "Chlamydomonas reinhardtii: A Factory of Nutraceutical and Food Supplements for Human Health" Molecules 28, no. 3: 1185. https://doi.org/10.3390/molecules28031185