
Identifications of Surfactin-Type Biosurfactants Produced by Bacillus Species Isolated from Rhizosphere of Vegetables

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Identification of the Isolated Strains
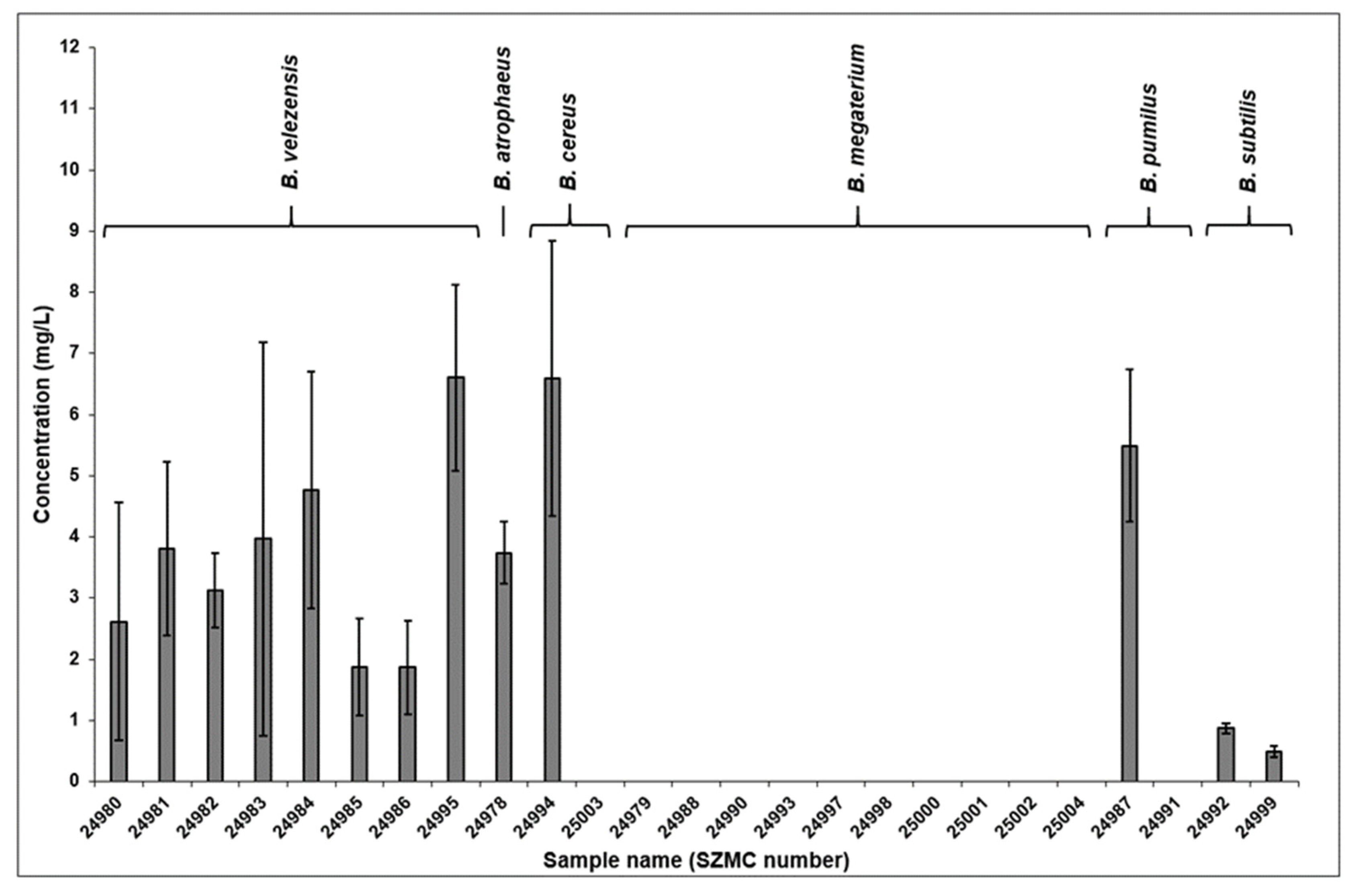
2.2. Quantitative Results of the Total Surfactin Production
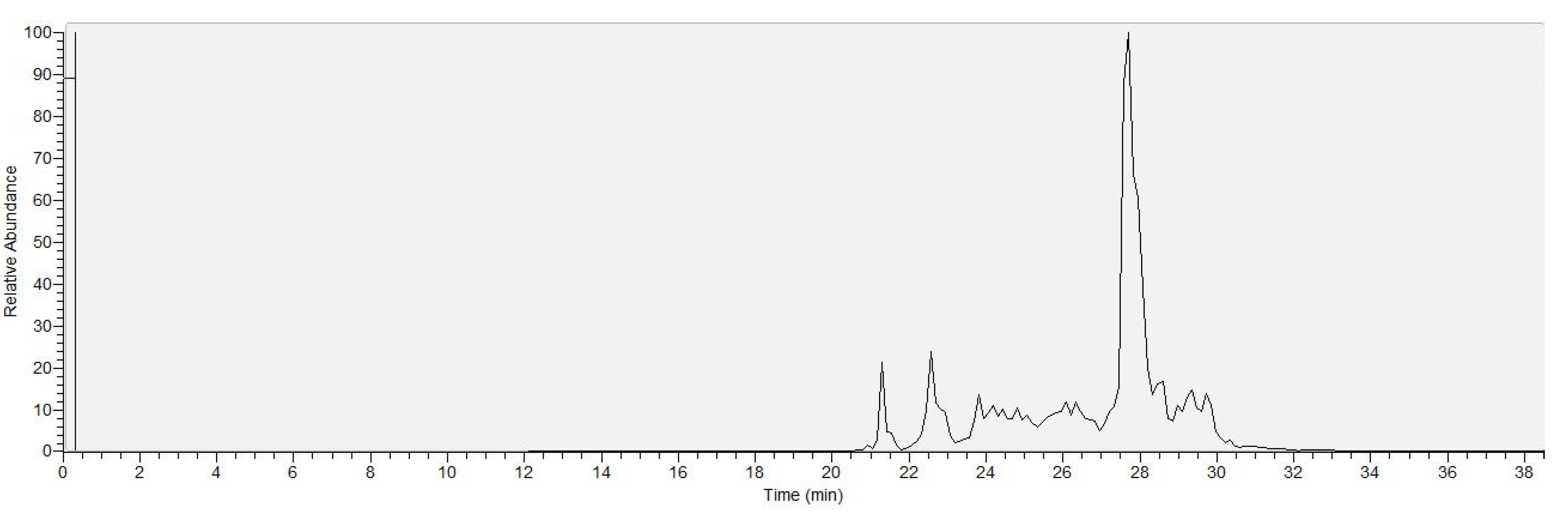
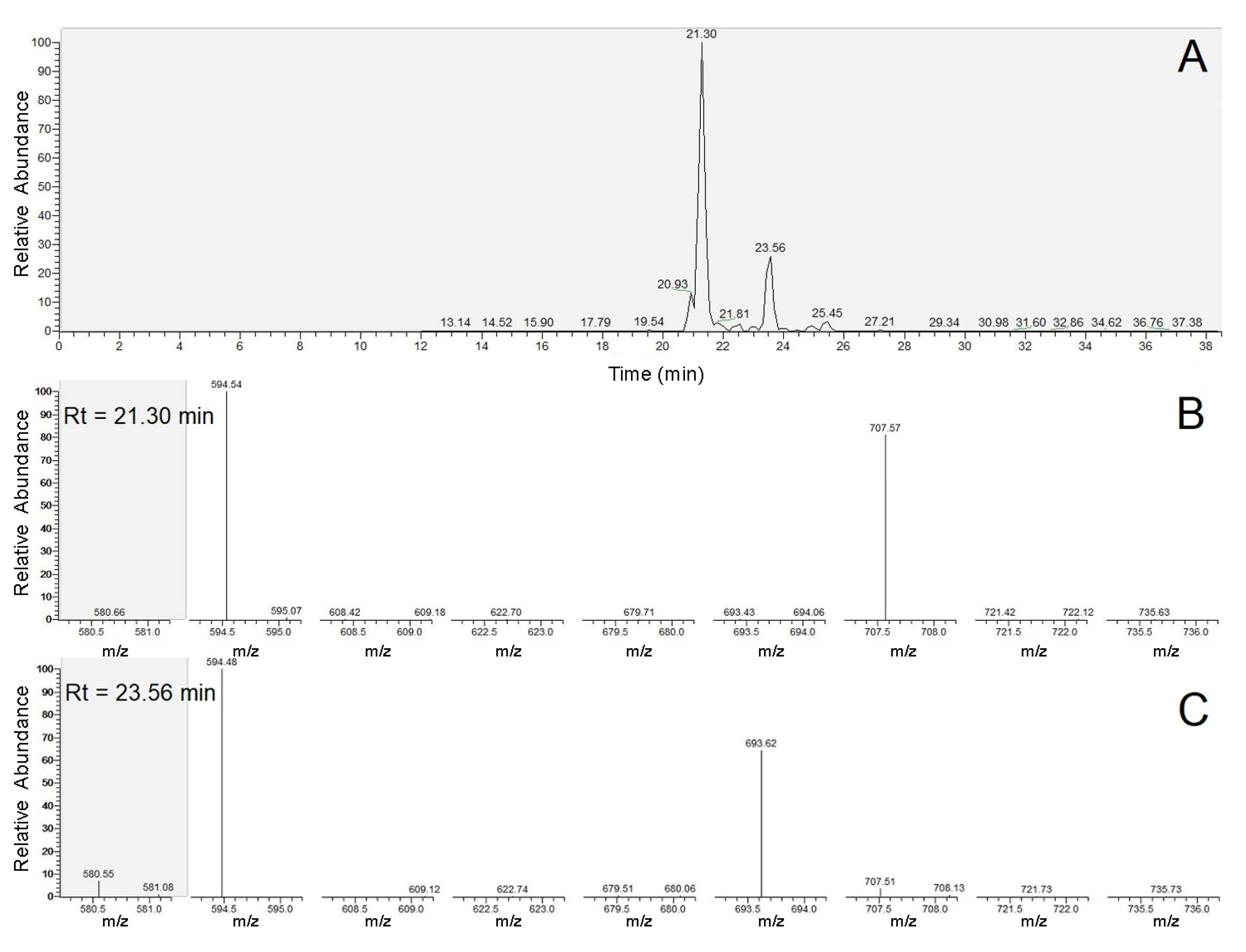
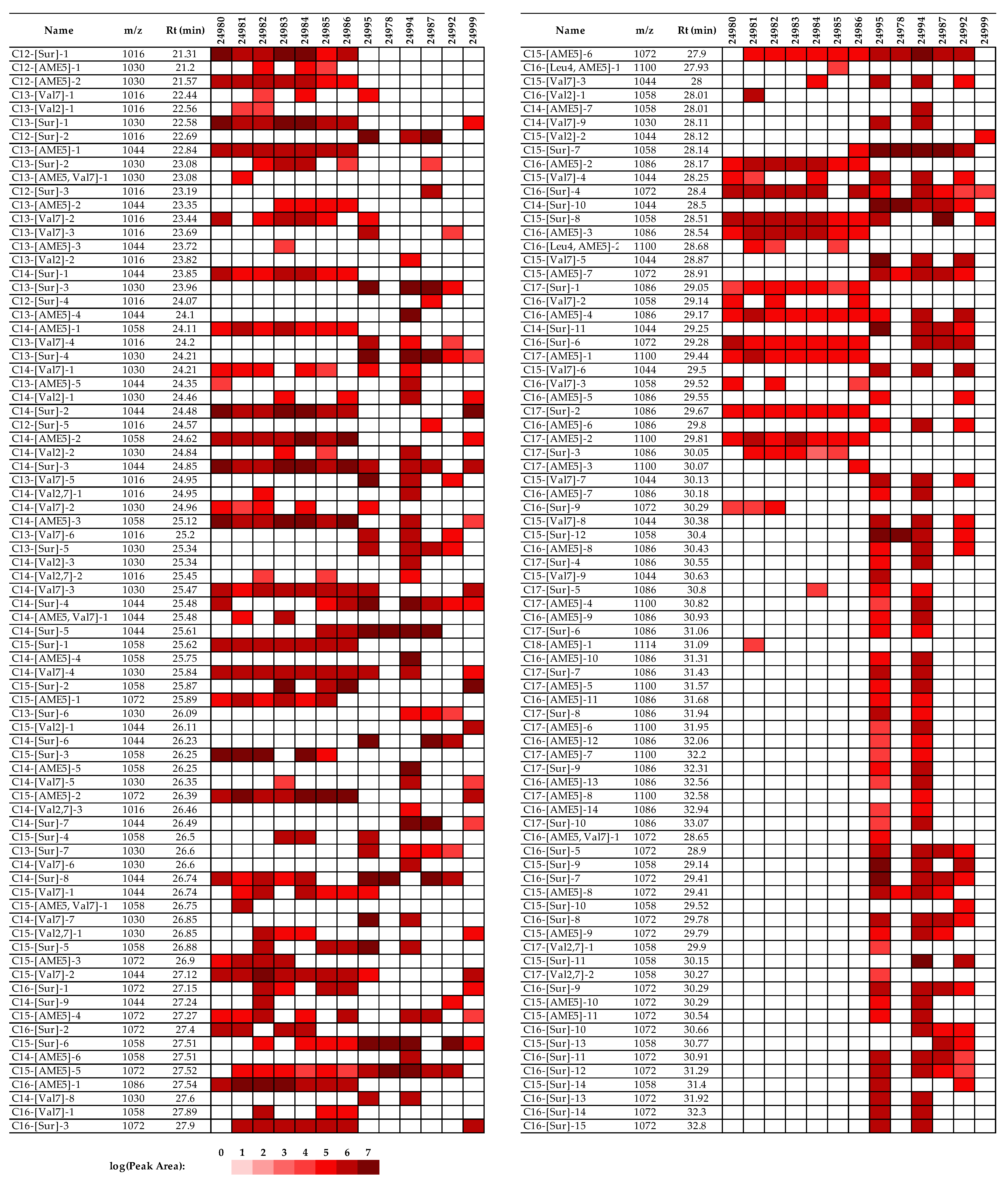
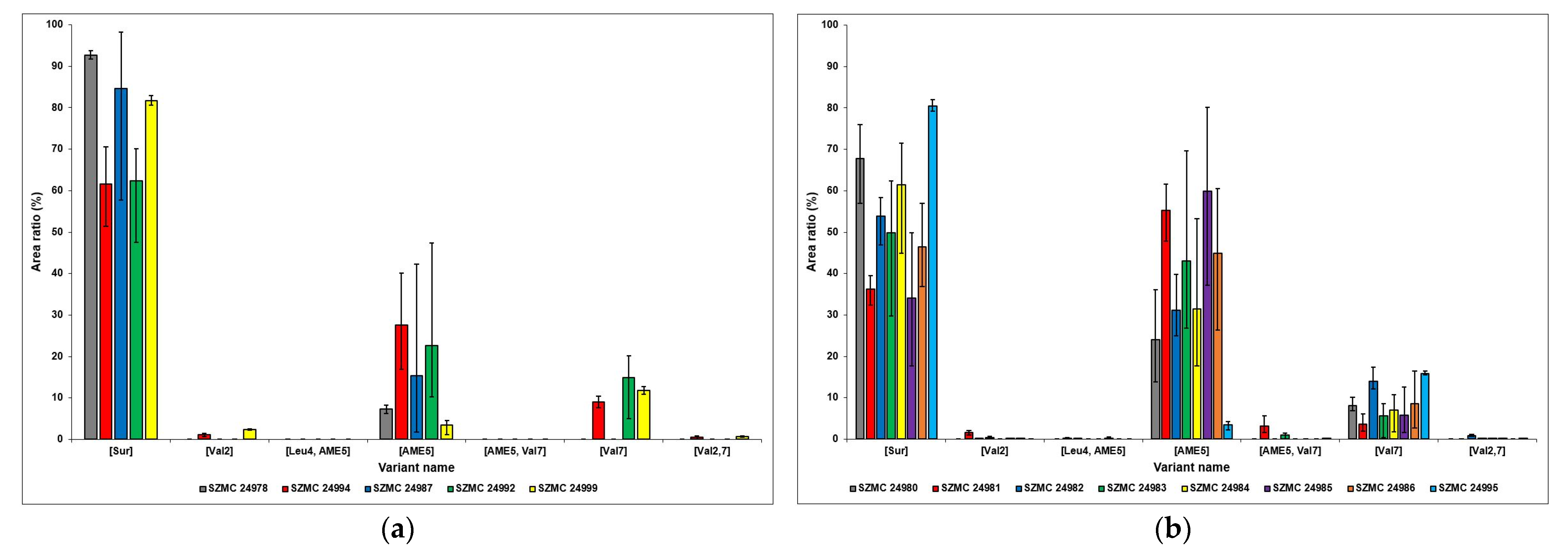
2.3. Identification of the Detected Surfactins
2.4. Comparison of the Surfactin Production Profiles
2.5. The Biocontrol Properties of the Examined Bacillus Isolates
3. Discussion
4. Materials and Methods
4.1. Strains Maintenance
4.2. Nomenclature of Surfactin Variants
4.3. Culture Conditions and Sample Preparation for surfactin Analysis
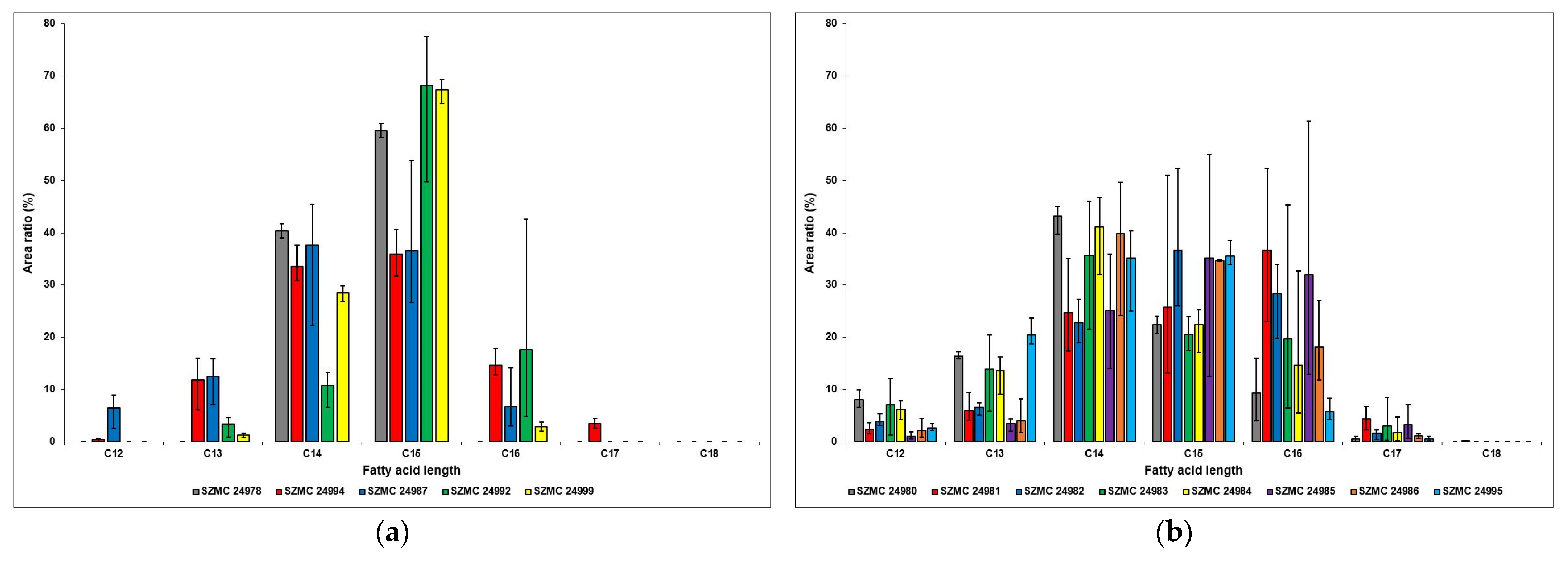
4.4. Identification of Bacillus Isolates by Fatty Acid Methyl Ester (FAME) Analysis
4.5. Analytical Parameters
4.6. Inhibition Assays on Phytopathogen Microorganisms
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Arima, K.; Kakinuma, A.; Tamura, G. Surfactin, a crystalline peptidelipid surfactant produced by Bacillus subtilis: Isolation, characterization and its inhibition of fibrin clot formation. Biochem. Biophys. Res. Commun. 1968, 31, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-C.; Juang, R.-S.; Wei, Y.-H. Applications of a lipopeptide biosurfactant, surfactin, produced by microorganisms. Biochem. Eng. J. 2015, 103, 158–169. [Google Scholar] [CrossRef]
- Nieminen, T.; Rintaluoma, N.; Andersson, M.; Taimisto, A.-M.; Ali-Vehmas, T.; Seppälä, A.; Priha, O.; Salkinoja-Salonen, M. Toxinogenic Bacillus pumilus and Bacillus licheniformis from mastitic milk. Vet. Microbiol. 2007, 124, 329–339. [Google Scholar] [CrossRef] [PubMed]
- From, C.; Hormazabal, V.; Hardy, S.P.; Granum, P.E. Cytotoxicity in Bacillus mojavensis is abolished following loss of surfactin synthesis: Implications for assessment of toxicity and food poisoning potential. Int. J. Food Microbiol. 2007, 117, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Pecci, Y.; Rivardo, F.; Martinotti, M.G.; Allegrone, G. LC/ESI-MS/MS characterisation of lipopeptide biosurfactants produced by the Bacillus licheniformis v9t14 strain. J. Mass Spectrom. 2010, 4, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, F.; Castro, M.; Príncipe, A.; Borioli, G.; Fischer, S.; Mori, G.; Jofré, E. The plant-associated Bacillus amyloliquefaciens strains MEP218 and ARP23 capable of producing the cyclic lipopeptides iturin or surfactin and fengycin are effective in biocontrol of sclerotinia stem rot disease. J. Appl. Microbiol. 2011, 112, 159–174. [Google Scholar] [CrossRef]
- Bonmatin, J.-M.; Laprevote, O.; Peypoux, F. Diversity among microbial cyclic lipopeptides: Iturins and surfactins. Activity-structure relationships to design new bioactive agents. Comb. Chem. High Throughput Screen. 2003, 6, 541–556. [Google Scholar] [CrossRef] [PubMed]
- Vollenbroich, D.; Pauli, G.; Ozel, M.; Vater, J. Antimycoplasma properties and application in cell culture of surfactin, a lipopeptide antibiotic from Bacillus subtilis. Appl. Environ. Microbiol. 1997, 63, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Duarte, C.; Gudiña, E.J.; Lima, C.F.; Rodrigues, L.R. Effects of biosurfactants on the viability and proliferation of human breast cancer cells. AMB Express 2014, 4, 40. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, C.; Dong, B.; Ma, X.; Hou, L.; Cao, X.; Wang, C. Anti-inflammatory activity and mechanism of surfactin in lipopolysaccharide-activated macrophages. Inflammation 2015, 38, 756–764. [Google Scholar] [CrossRef]
- Wang, X.; Hu, W.; Zhu, L.; Yang, Q. Bacillus subtilis and surfactin inhibit the transmissible gastroenteritis virus from entering the intestinal epithelial cells. Biosci. Rep. 2017, 37, BSR20170082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulligan, C.N. Environmental applications for biosurfactants. Environ. Pollut. 2005, 133, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Albino, J.D.; Nambi, I.M. Effect of biosurfactants on the aqueous solubility of PCE and TCE. J. Environ. Sci. Health Part A 2009, 44, 1565–1573. [Google Scholar] [CrossRef] [PubMed]
- Zou, A.; Liu, J.; Garamus, V.M.; Zheng, K.; Willumeit, R.; Mu, B. Interaction between the natural lipopeptide [Glu1, Asp5] surfactin-C15 and hemoglobin in aqueous solution. Biomacromolecules 2010, 11, 593–599. [Google Scholar] [CrossRef]
- Hafeez, F.Y.; Naureen, Z.; Sarwar, A. Surfactin: An emerging biocontrol tool for agriculture sustainability. In Plant Growth Promoting Rhizobacteria for Agricultural Sustainability, 1st ed.; Kumar, A., Meena, V., Eds.; Springer: Singapore, 2019; pp. 203–213. [Google Scholar]
- Bóka, B.; Manczinger, L.; Kecskeméti, A.; Chandrasekaran, M.; Kadaikunnan, S.; Alharbi, N.S.; Vágvölgyi, C.; Szekeres, A. Ion trap mass spectrometry of surfactins produced by Bacillus subtilis SZMC 6179J reveals novel fagmentation features of cyclic lipopeptides. Rapid Commun. Mass Spectrom. 2016, 30, 1581–1590. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.-S.; Gao, H.; Hong, K.; Yu, J.; Jiang, M.-M.; Lin, H.-P.; Ye, W.-C.; Yao, X.-S. Complete assignments of 1H and 13C NMR spectral data of nine surfactin isomers. Magn. Reson. Chem. 2007, 45, 792–796. [Google Scholar] [CrossRef]
- Tang, J.-S.; Zhao, F.; Gao, H.; Dai, Y.; Yao, Z.-H.; Hong, K.; Li, J.; Ye, W.-C.; Yao, X.-S. Characterization and online detection of surfactin isomers based on HPLC-MSn analyses and their inhibitory effects on the overproduction of nitric oxide and the release of TNF-α and IL-6 in LPS-induced macrophages. Mar. Drugs 2010, 8, 2605–2618. [Google Scholar] [CrossRef] [Green Version]
- Moro, G.V.; Almeida, R.T.R.; Napp, A.P.; Porto, C.; Pilau, E.J.; Lüdtke, D.S.; Moro, A.V.; Vainstein, M.H. Identification and ultra-high-performance liquid chromatography coupled with high-resolution mass spectrometry characterization of biosurfactants, including a new surfactin, isolated from oil-contaminated environments. Microb. Biotechnol. 2018, 11, 759–769. [Google Scholar] [CrossRef]
- Kecskeméti, A.; Bartal, A.; Bóka, B.; Kredics, L.; Manczinger, L.; Shine, K.; Alharby, N.S.; Khaled, J.M.; Varga, M.; Vágvölgyi, C.; et al. High-frequency occurrence of surfactin monomethyl isoforms in the ferment broth of a Bacillus subtilis strain revealed by ion trap mass spectrometry. Molecules 2018, 23, 2224. [Google Scholar] [CrossRef] [Green Version]
- Slivinski, C.T.; Mallmann, E.; de Araújo, J.M.; Mitchell, D.A.; Krieger, N. Production of surfactin by Bacillus pumilus UFPEDA 448 in solid-state fermentation using a medium based on okara with sugarcane bagasse as a bulking agent. Process Biochem. 2012, 47, 1848–1855. [Google Scholar] [CrossRef]
- Joshi, S.J.; Al-Wahaibi, Y.M.; Al-Bahry, S.N.; Elshafie, A.E.; Al-Bemani, A.S.; Al-Mandhari, M.S. Production, characterization, and application of Bacillus licheniformis W16 biosurfactant in enhancing oil recovery. Front. Microbiol. 2016, 7, 1853. [Google Scholar] [CrossRef] [PubMed]
- Bartal, A.; Vigneshwari, A.; Bóka, B.; Vörös, M.; Takács, I.; Kredics, L.; Manczinger, L.; Varga, M.; Vágvölgyi, C.; Szekeres, A. Effects of different cultivation parameters on the production of surfactin variants by a Bacillus subtilis strain. Molecules 2018, 23, 2675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czinkóczky, R.; Németh, Á. The effect of pH on biosurfactant production by Bacillus subtilis DSM10. Hung. J. Ind. Chem. 2020, 48, 37–43. [Google Scholar] [CrossRef]
- Huynh, T.; Vörös, M.; Kedves, O.; Turbat, A.; Sipos, G.; Leitgeb, B.; Kredics, L.; Vágvölgyi, C.; Szekeres, A. Discrimination between the two closely related species of the operational group B. amyloliquefaciens based on whole-cell fatty acid profiling. Microorganisms 2022, 10, 418. [Google Scholar] [CrossRef]
- Diomandé, S.E.; Nguyen-The, C.; Guinebretière, M.H.; Broussolle, V.; Brillard, J. Role of fatty acids in Bacillus environmental adaptation. Front. Microbiol. 2015, 6, 813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foysal, M.J.; Lisa, A.K. Isolation and characterization of Bacillus sp. strain BC01 from soil displaying potent antagonistic activity against plant and fish pathogenic fungi and bacteria. J. Genet. Eng. Biotechnol. 2018, 16, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Zalila-Kolsi, I.; Ben Mahmoud, A.; Ali, H.; Sellami, S.; Nasfi, Z.; Tounsi, S.; Jamoussi, K. Antagonist effects of Bacillus spp. strains against Fusarium graminearum for protection of durum wheat (Triticum turgidum L. subsp. durum). Microbiol. Res. 2016, 192, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Dubey, R.C.; Maheshwari, D.K. Bacillus strains isolated from rhizosphere showed plant growth promoting and antagonistic activity against phytopathogens. Microbiol. Res. 2012, 167, 493–499. [Google Scholar] [CrossRef]
- Vágvölgyi, C.; Sajben-Nagy, E.; Bóka, B.; Vörös, M.; Berki, A.; Palágyi, A.; Krisch, J.; Skrbić, B.; Durišić-Mladenović, N.; Manczinger, L. Isolation and characterization of antagonistic Bacillus strains capable to degrade ethylenethiourea. Curr. Microbiol. 2013, 66, 243–250. [Google Scholar] [CrossRef]
- Fan, B.; Blom, J.; Klenk, H.P.; Borriss, R. Bacillus amyloliquefaciens, Bacillus velezensis, and Bacillus siamensis form an “Operational group B. amyloliquefaciens” within the B. subtilis species complex. Front. Microbiol. 2017, 8, 22. [Google Scholar] [CrossRef]
- Ruiz-García, C.; Béjar, V.; Martínez-Checa, F.; Llamas, I.; Quesada, E. Bacillus velezensis sp. nov., a surfactant-producing bacterium isolated from the river Vélez in Málaga, southern Spain. Int. J. Syst. Evol. Microbiol. 2005, 55, 191–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grady, E.N.; MacDonald, J.; Ho, M.T.; Weselowski, B.; McDowell, T.; Solomon, O.; Renaud, J.; Yuan, Z.C. Characterization and complete genome analysis of the surfactin-producing, plant-protecting bacterium Bacillus velezensis 9D-6. BMC Microbiol. 2019, 19, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Zhao, D.; Qi, G.; Mao, Z.; Hu, X.; Du, B.; Liu, K.; Ding, Y. Effects of Bacillus velezensis FKM10 for promoting the growth of Malus hupehensis Rehd. and inhibiting Fusarium verticillioides. Front. Microbiol. 2020, 10, 2889. [Google Scholar] [CrossRef]
- Liu, G.; Kong, Y.; Fan, Y.; Geng, C.; Peng, D.; Sun, M. Data on genome analysis of Bacillus velezensis LS69. Data Brief 2017, 13, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Grangemard, I.; Peypoux, F.; Wallach, J.; Das, B.C.; Labbé, H.; Caille, A.; Genest, M.; Maget-Dana, R.; Ptak, M.; Bonmatin, J.-M. Lipopeptides with improved properties: Structure by NMR, purification by HPLC and structure–activity relationships of new isoleucyl-rich surfactins. J. Pept. Sci. 1997, 3, 145–154. [Google Scholar] [CrossRef]
- Roepstorff, P.; Fohlman, J. Proposal for a common nomenclature for sequence ions in mass spectra of peptides. Biol. Mass Spectrom. 1984, 11, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Biemann, K. Sequencing of peptides by tandem mass spectrometry and high-energy collision-induced dissociation. Methods Enzymol. 1990, 193, 455–479. [Google Scholar]
- Hue, N.; Serani, L.; Laprévote, O. Structural investigation of cyclic peptidolipids from Bacillus subtilis by high-energy tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2001, 15, 203–209. [Google Scholar] [CrossRef]
- Besson, F.; Chevanet, C.; Michel, G. Influence of the culture medium on the production of iturin A by Bacillus subtilis. J. Gen. Microbiol. 1987, 133, 767–772. [Google Scholar] [CrossRef] [Green Version]
- MIDI Inc. The Sherlock Chromatographic Analysis System Operating Manual, 6th ed.; MIDI, Inc.: Newark, DE, USA, 2018. [Google Scholar]
- Büchner, R.; Vörös, M.; Allaga, H.; Varga, A.; Bartal, A.; Szekeres, A.; Varga, S.; Bajzát, J.; Bakos-Barczi, N.; Misz, A.; et al. Selection and characterization of a Bacillus strain for potential application in industrial production of white button mushroom (Agaricus bisporus). Agronomy 2022, 12, 467. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture Collection Number 1 | Source | Identification |
|---|---|---|
| SZMC 24978 | Totovo selo, Serbia, soil sample, tomato | B. atrophaeus |
| SZMC 24979 | Szeged, Hungary, soil sample, tomato | B. megaterium |
| SZMC 24980 | Szeged, Hungary soil sample, pepper | B. velezensis |
| SZMC 24981 | Szeged, Hungary soil sample, pepper | B. velezensis |
| SZMC 24982 | Szeged, Hungary soil sample, pepper | B. velezensis |
| SZMC 24983 | Szeged, Hungary soil sample, pepper | B. velezensis |
| SZMC 24984 | Szeged, Hungary soil sample, pepper | B. velezensis |
| SZMC 24985 | Szeged, Hungary soil sample, pepper | B. velezensis |
| SZMC 24986 | Szeged, Hungary soil sample, tomato | B. velezensis |
| SZMC 24987 | Cantavir, Serbia, tomato | B. pumilus |
| SZMC 24988 | Szeged, Hungary soil sample, tomato | B. megaterium |
| SZMC 24990 | Szeged, Hungary soil sample, tomato | B. megaterium |
| SZMC 24991 | Szeged, Hungary soil sample, tomato | B. pumilus |
| SZMC 24992 | Cantavir, Serbia, pepper | B. subtilis |
| SZMC 24993 | Szeged, Hungary soil sample, pepper | B. megaterium |
| SZMC 24994 | Cantavir, Serbia, tomato | B. cereus |
| SZMC 24995 | Cantavir, Serbia, tomato | B. velezensis |
| SZMC 24997 | Szeged, Hungary soil sample, carrot | B. megaterium |
| SZMC 24998 | Szeged, Hungary soil sample, carrot | B. megaterium |
| SZMC 24999 | Szeged, Hungary soil sample, carrot | B. subtilis |
| SZMC 25000 | Szeged, Hungary soil sample, paprika | B. megaterium |
| SZMC 25001 | Madaras, Hungary, pepper | B. megaterium |
| SZMC 25002 | Szeged, Hungary soil sample, paprika | B. megaterium |
| SZMC 25003 | Szeged, Hungary soil sample, paprika | B. cereus |
| SZMC 25004 | Szeged, Hungary soil sample, sweet potato | B. megaterium |
| Features | B. atrophaeus | B. cereus (GC-A) 1 | B. megaterium (GC-A) 1 | B. pumilus (GC-B) 1 | B. subtilis | B. velezensis |
|---|---|---|---|---|---|---|
| 13:0 iso | - | 9.69 | - | 0.50 | - | 0.93 |
| 14:0 iso | 1.37 | 3.61 | 4.31 | 0.79 | 0.85 | 1.05 |
| 14:0 | - | 2.91 | 1.72 | 1.23 | - | 3.04 |
| 15:0 iso | 13.53 | 34.78 | 38.02 | 50.83 | 24.24 | 30.25 |
| 15:0 anteiso | 46.28 | 3.93 | 41.56 | 23.48 | 39.18 | 32.54 |
| 16:0 iso | 4.11 | 5.56 | 0.62 | 1.44 | 2.37 | 1.56 |
| 16:1 w11c | 1.82 | - | 3.85 | 1.88 | 1.74 | 1.71 |
| 16:0 | 2.97 | 4.97 | 2.70 | 4.15 | 3.39 | 13.00 |
| 17:1 iso w10c | 1.81 | 4.69 | 0.57 | 2.26 | 2.44 | 0.82 |
| 17:0 iso | 6.29 | 9.00 | 1.86 | 8.12 | 10.94 | 8.18 |
| 17:1 iso w5c | - | 4.50 | - | - | - | - |
| 17:0 anteiso | 17.92 | 0.96 | 3.25 | 4.56 | 12.34 | 5.43 |
| Species | Strain Number | P. syringae | E. amylovora | E. carotovora | X. campestris | A. tumefaciens |
|---|---|---|---|---|---|---|
| Inhibition Zones * | ||||||
| B. atrophaeus | SZMC 24978 | - | ++ | + | + | + |
| B. cereus | SZMC 25003 | - | + | - | - | - |
| B. megaterium | SZMC 24979 | - | +++ | + | ++ | + |
| SZMC 24988 | - | - | - | - | + | |
| SZMC 25000 | - | + | - | - | - | |
| B. pumilus | SZMC 24991 | - | - | - | - | + |
| B. subtilis | SZMC 24992 | - | + | + | + | + |
| SZMC 24999 | - | + | - | - | - | |
| B. velezensis | SZMC 24980 | + | ++ | + | ++ | + |
| SZMC 24981 | + | +++ | + | ++ | + | |
| SZMC 24982 | + | +++ | + | ++ | + | |
| SZMC 24983 | + | +++ | + | ++ | + | |
| SZMC 24984 | + | +++ | + | +++ | + | |
| SZMC 24985 | - | +++ | + | ++ | + | |
| SZMC 24986 | + | +++ | - | + | + | |
| SZMC 24995 | + | + | + | - | + | |
| Species | Strain Number | F. graminearum | B. cinerea | F. solani | F. culmorum | C. gloeosporioides |
|---|---|---|---|---|---|---|
| Inhibition Rates * | ||||||
| B. atrophaeus | SZMC 24978 | - | +++ | - | - | - |
| B. cereus | SZMC 25003 | - | ++ | - | - | + |
| B. megaterium | SZMC 24979 | ++ | - | ++ | - | - |
| B. pumilus | SZMC 24987 | - | ++ | - | - | - |
| B. subtilis | SZMC 24992 | ++ | +++ | ++ | ++ | - |
| SZMC 24999 | ++ | +++ | ++ | ++ | +++ | |
| B. velezensis | SZMC 24980 | + | +++ | ++ | ++ | ++ |
| SZMC 24981 | ++ | +++ | ++ | +++ | ++ | |
| SZMC 24982 | ++ | +++ | ++ | ++ | ++ | |
| SZMC 24983 | ++ | +++ | + | +++ | ++ | |
| SZMC 24984 | ++ | +++ | ++ | ++ | ++ | |
| SZMC 24985 | ++ | +++ | ++ | ++ | ++ | |
| SZMC 24986 | ++ | +++ | ++ | ++ | ++ | |
| SZMC 24995 | ++ | +++ | ++ | +++ | ++ | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartal, A.; Huynh, T.; Kecskeméti, A.; Vörös, M.; Kedves, O.; Allaga, H.; Varga, M.; Kredics, L.; Vágvölgyi, C.; Szekeres, A. Identifications of Surfactin-Type Biosurfactants Produced by Bacillus Species Isolated from Rhizosphere of Vegetables. Molecules 2023, 28, 1172. https://doi.org/10.3390/molecules28031172
Bartal A, Huynh T, Kecskeméti A, Vörös M, Kedves O, Allaga H, Varga M, Kredics L, Vágvölgyi C, Szekeres A. Identifications of Surfactin-Type Biosurfactants Produced by Bacillus Species Isolated from Rhizosphere of Vegetables. Molecules. 2023; 28(3):1172. https://doi.org/10.3390/molecules28031172
Chicago/Turabian StyleBartal, Attila, Thu Huynh, Anita Kecskeméti, Mónika Vörös, Orsolya Kedves, Henrietta Allaga, Mónika Varga, László Kredics, Csaba Vágvölgyi, and András Szekeres. 2023. "Identifications of Surfactin-Type Biosurfactants Produced by Bacillus Species Isolated from Rhizosphere of Vegetables" Molecules 28, no. 3: 1172. https://doi.org/10.3390/molecules28031172