
Xanthohumol-Enriched Beer Does Not Exert Antitumorigenic Effects on HeLa Cell Line In Vivo
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
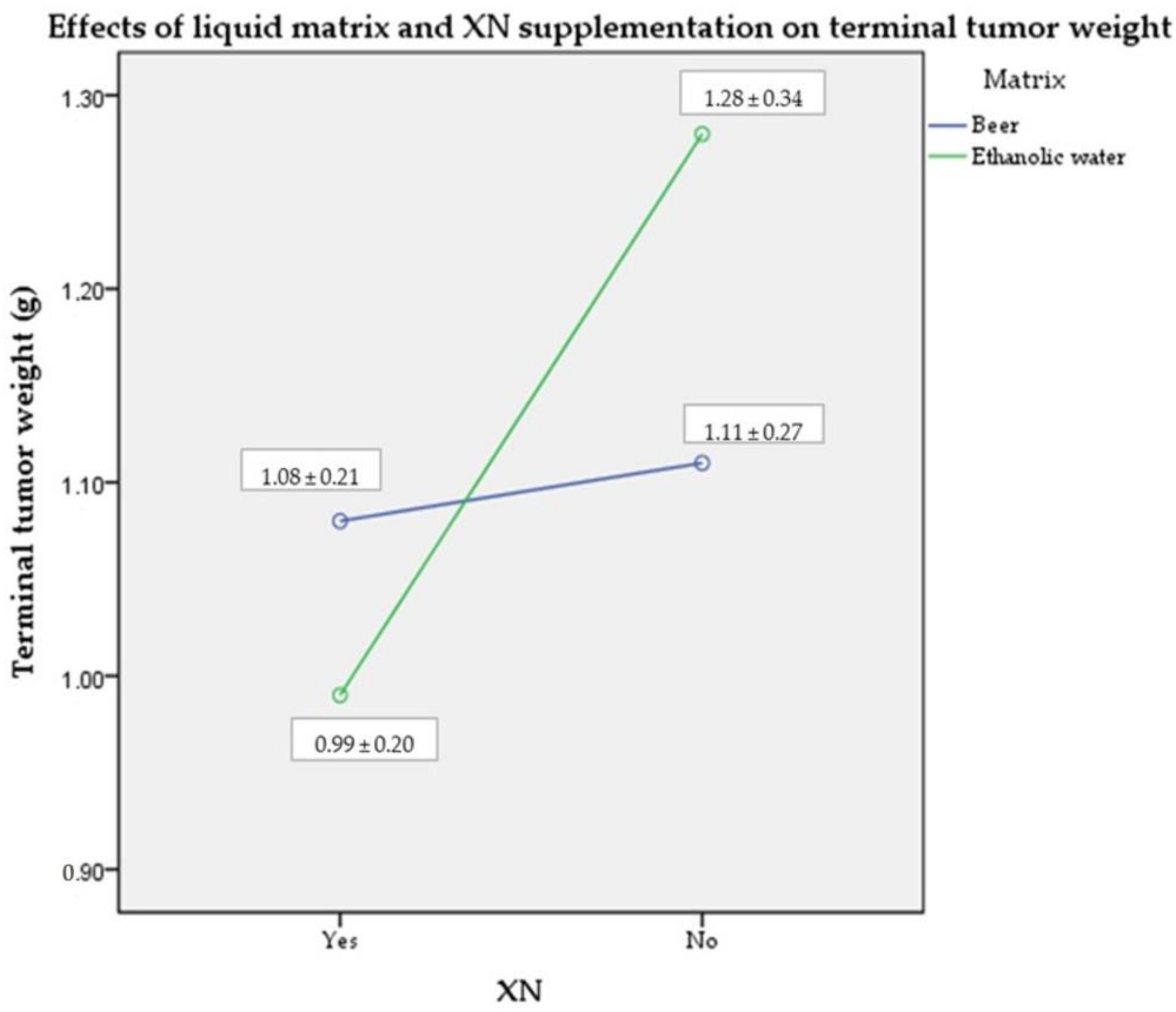
2.1. Terminal Tumor Weight
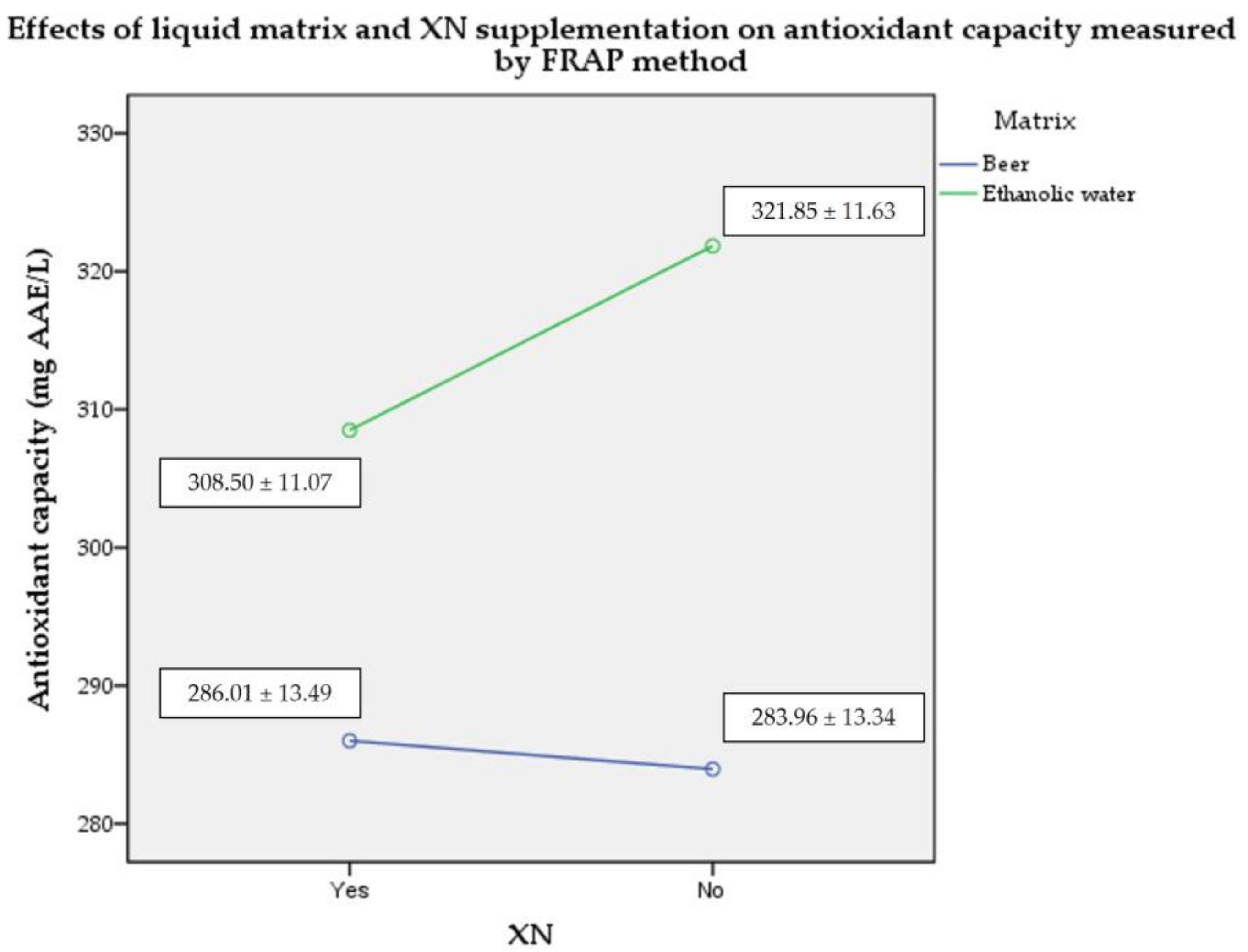
2.2. Plasma Antioxidant Capacity by FRAP Assay
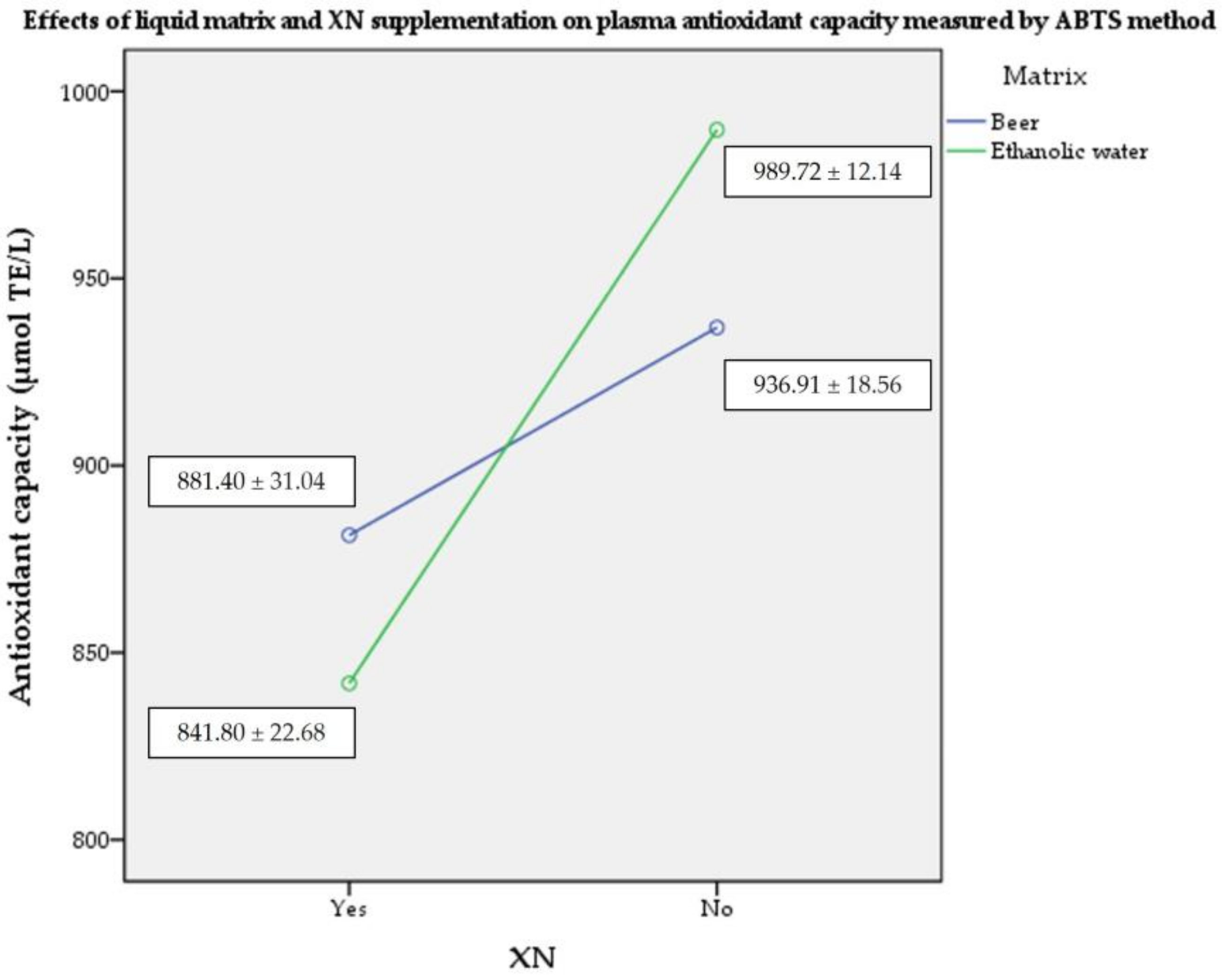
2.3. Plasma Antioxidant Capacity by ABTS Assay
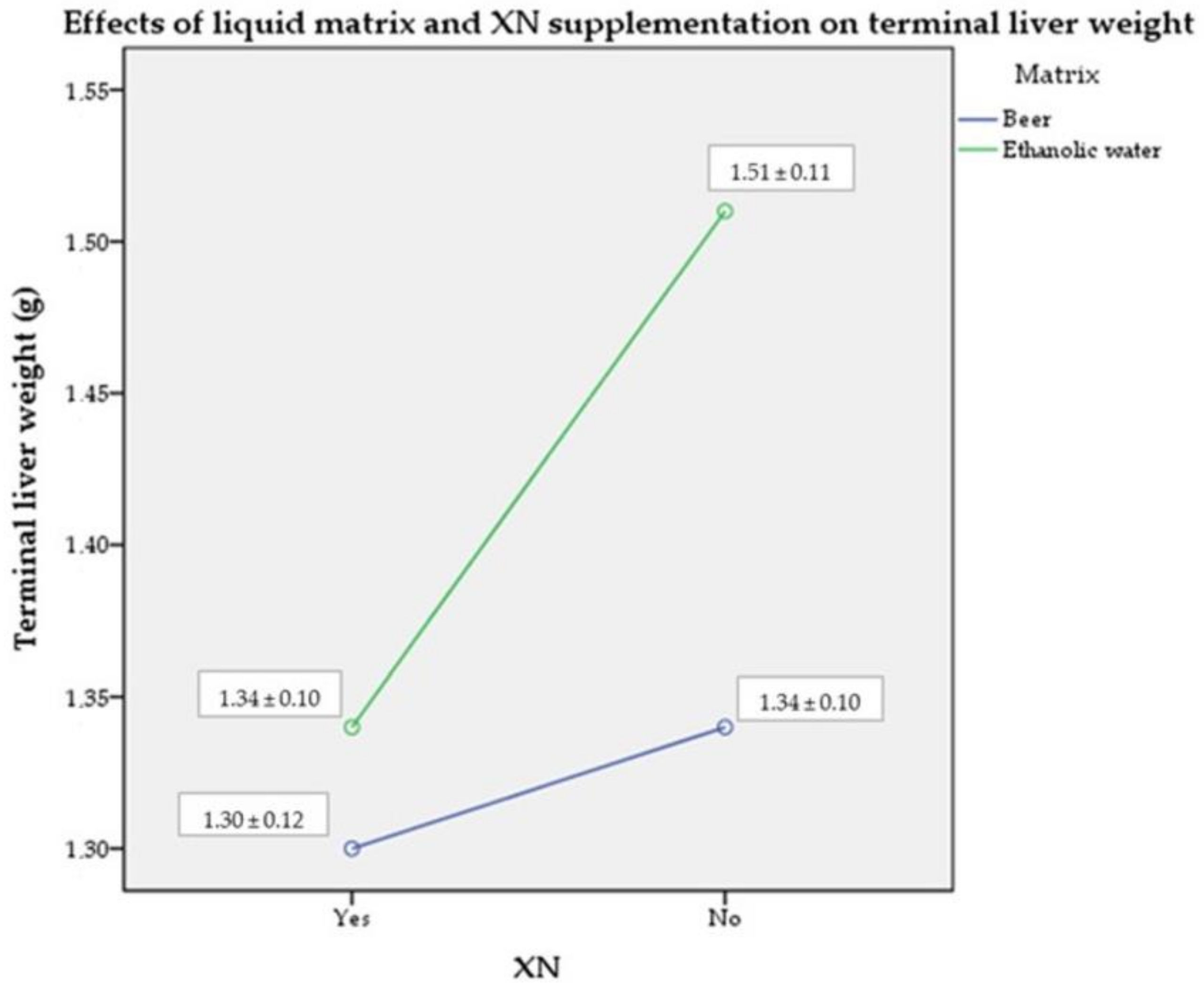
2.4. Terminal Liver Weight
3. Discussion
Limitations of the Study
4. Materials and Methods
4.1. Analytical Procedure of XN Content Measurement
4.2. Study Subjects and Study Design
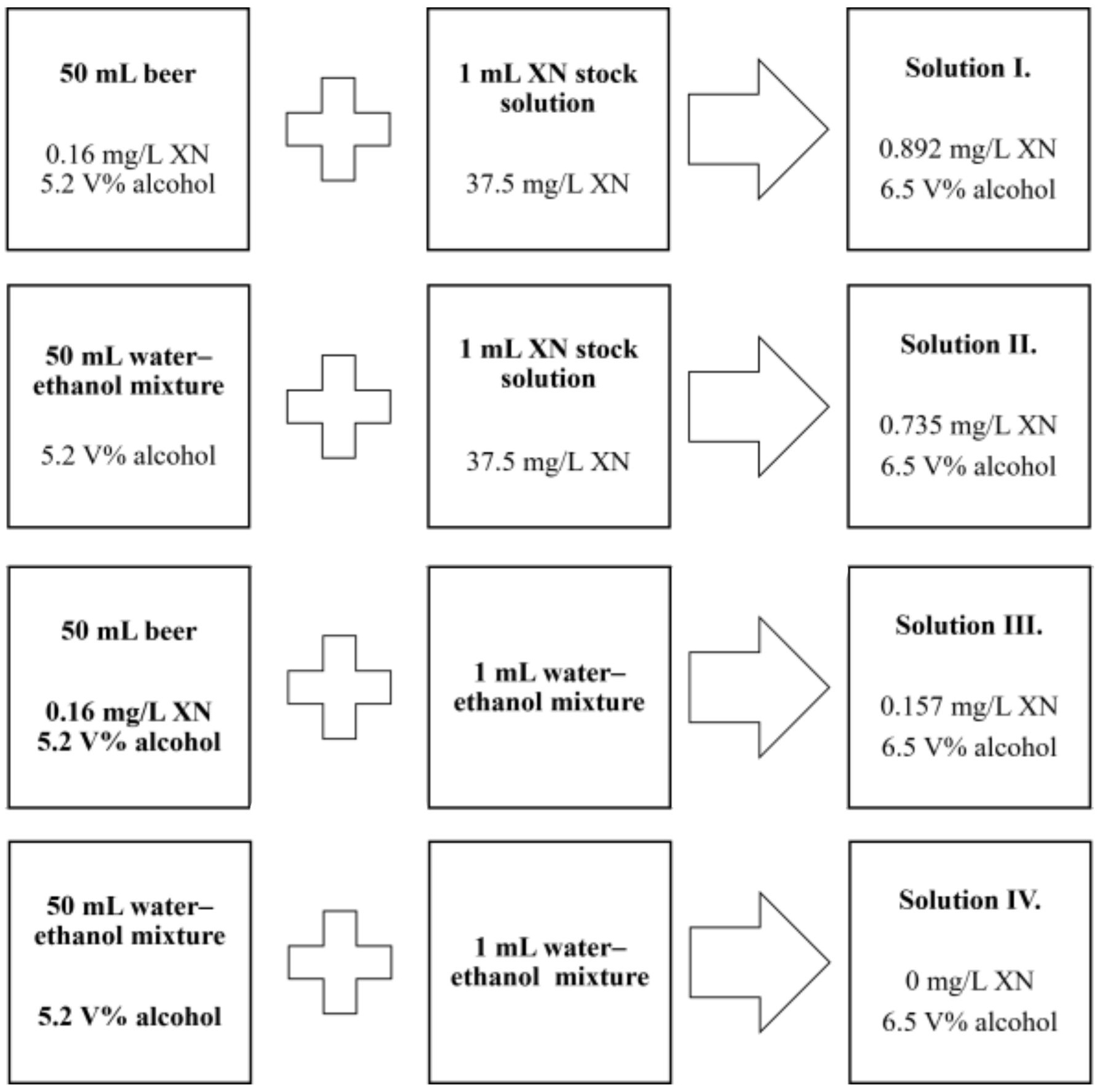
4.3. Study Sample Preparation
4.4. Measurement of Antioxidant Capacity
4.4.1. FRAP Method
4.4.2. ABTS Method
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vesaghhamedani, S.; Ebrahimzadeh, F.; Najafi, E.; Shabgah, O.G.; Askari, E.; Shabgah, A.G.; Mohammadi, H.; Jadidi-Niaragh, F.; Navashenaq, J.G. Xanthohumol: An Underestimated, While Potent and Promising Chemotherapeutic Agent in Cancer Treatment. Prog. Biophys. Mol. Biol. 2022, 172, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Harish, V.; Haque, E.; Śmiech, M.; Taniguchi, H.; Jamieson, S.; Tewari, D.; Bishayee, A. Xanthohumol for Human Malignancies: Chemistry, Pharmacokinetics and Molecular Targets. Int. J. Mol. Sci. 2021, 22, 4478. [Google Scholar] [CrossRef]
- Dokduang, H.; Yongvanit, P.; Namwat, N. Xanthohumol Inhibits STAT3 Activation Pathway Leading to Growth Suppression and Apoptosis Induction in Human Cholangiocarcinoma Cells. Oncol. Rep. 2016, 35, 2065–2072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, K.; Matsuo, Y.; Imafuji, H.; Okubo, T.; Maeda, Y.; Sato, T.; Shamoto, T.; Tsuboi, K.; Morimoto, M.; Takahashi, H.; et al. Xanthohumol Inhibits Angiogenesis by Suppressing Nuclear Factor-ΚB Activation in Pancreatic Cancer. Cancer Sci. 2018, 109, 132–140. [Google Scholar] [CrossRef] [Green Version]
- Shikata, Y.; Yoshimaru, T.; Komatsu, M.; Katoh, H.; Sato, R.; Kanagaki, S.; Okazaki, Y.; Toyokuni, S.; Tashiro, E.; Ishikawa, S.; et al. Protein Kinase A Inhibition Facilitates the Antitumor Activity of Xanthohumol, a Valosin-Containing Protein Inhibitor. Cancer Sci. 2017, 108, 785–794. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Zhou, C.; Liu, F.; Zhang, W.; Chen, J.; Pan, Y.; Ma, L.; Liu, Q.; Du, Y.; Yang, J.; et al. Inhibition of Breast Cancer Cell Survival by Xanthohumol via Modulation of the Notch Signaling Pathway in Vivo and in Vitro. Oncol. Lett. 2018, 15, 908–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venè, R.; Benelli, R.; Minghelli, S.; Astigiano, S.; Tosetti, F. Xanthohumol Impairs Human Prostate Cancer Cell Growth and Invasion and Diminishes the Incidence and Progression of Advanced Tumors in TRAMP Mice. Mol. Med. 2012, 18, 1292–1302. [Google Scholar] [CrossRef]
- Walden, D.; Kunnimalaiyaan, S.; Sokolowski, K.; Gamblin, T.C.; Kunnimalaiyaan, M. Antiproliferative and Apoptotic Effects of Xanthohumol in Cholangiocarcinoma. Oncotarget 2017, 8, 88069–88078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venturelli, S.; Burkard, M.; Biendl, M.; Lauer, U.M.; Frank, J.; Busch, C. Prenylated Chalcones and Flavonoids for the Prevention and Treatment of Cancer. Nutrition 2016, 32, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Éliás, A.J.; Bodor, Z.; Benedek, C. Xanthohumol from Hop (Humulus lupulus L.). In Sustainable and Functional Foods from Plants; Goyal, M.R., Nath, A., Kovács, Z., Eds.; Apple Academic Press: Palm Bay, FL, USA, 2022; ISBN 9781774914540. [Google Scholar]
- Zugravu, C.A.; Bohiltea, R.E.; Salmen, T.; Pogurschi, E.; Otelea, M.R. Antioxidants in Hops: Bioavailability, Health Effects and Perspectives for New Products. Antioxidants 2022, 11, 241. [Google Scholar] [CrossRef]
- Ciriminna, R.; Albanese, L.; Di Stefano, V.; Delisi, R.; Avellone, G.; Meneguzzo, F.; Pagliaro, M. Beer Produced via Hydrodynamic Cavitation Retains Higher Amounts of Xanthohumol and Other Hops Prenylflavonoids. Lwt 2018, 91, 160–167. [Google Scholar] [CrossRef]
- Karabín, M.; Jelínek, L.; Hudcová, T.; Kotlíková, B.; Dostálek, P. New Approach to the Production of Xanthohumol-Enriched Beers. J. Inst. Brew. 2013, 119, 98–102. [Google Scholar] [CrossRef]
- Protsenko, L.; Rudyk, R.; Hryniuk, T.; Vlasenko, A.; Protsenko, A.; Litvynchuk, S.; Ovadenko, O. Beer Enrichment with Biologically Active Hop Compounds. Ukr. Food J. 2018, 7, 65–78. [Google Scholar] [CrossRef]
- Wunderlich, S.; Zürcher, A.; Back, W. Enrichment of Xanthohumol in the Brewing Process. Mol. Nutr. Food Res. 2005, 49, 874–881. [Google Scholar] [CrossRef]
- Lugasi, A. Polyphenol Content and Antioxidant Properties of Beer. Acta Aliment. 2003, 32, 181–192. [Google Scholar] [CrossRef]
- Koren, D.; Kun, S.; Hegyesné Vecseri, B.; Kun-Farkas, G. Study of Antioxidant Activity during the Malting and Brewing Process. J. Food Sci. Technol. 2019, 56, 3801–3809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shishehbor, F.; Shahi, M.M.; Zarei, M.; Saki, A.; Shirani, F.; Zare, M. Effects of Concentrated Pomegranate Juice on Subclinical Inflammation and Cardiometabolic Risk Factors for Type 2 Diabetes : A Quasi- Experimental Study. Int. J. Endocrinol. Metab. 2016, 14, e338835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasowski, B.; Leontowicz, M.; Leontowicz, H.; Katrich, E.; Lojek, A.; Číž, M.; Trakhtenberg, S.; Gorinstein, S. The Influence of Beer with Different Antioxidant Potential on Plasma Lipids, Plasma Antioxidant Capacity, and Bile Excretion of Rats Fed Cholesterol-Containing and Cholesterol-Free Diets. J. Nutr. Biochem. 2004, 15, 527–533. [Google Scholar] [CrossRef]
- Secretan, B.; Straif, K.; Baan, R.; Grosse, Y.; El Ghissassi, F.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Freeman, C.; Galichet, L.; et al. A Review of Human Carcinogens—Part E: Tobacco, Areca Nut, Alcohol, Coal Smoke, and Salted Fish. Lancet Oncol. 2009, 10, 1033–1034. [Google Scholar] [CrossRef]
- Dorn, C.; Kraus, B.; Motyl, M.; Weiss, T.S.; Gehrig, M. Xanthohumol, a Chalcon Derived from Hops, Inhibits Hepatic Inflammation and Fibrosis. Mol. Nutr. Food Res. 2010, 54, 205–213. [Google Scholar] [CrossRef]
- Kanuri, G.; Wagnerberger, S.; Landmann, M.; Prigl, E.; Hellerbrand, C.; Bischoff, S.C.; Bergheim, I. Effect of Acute Beer Ingestion on the Liver : Studies in Female Mice. Eur. J. Nutr. 2015, 54, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Landmann, M.; Wagnerberger, S.; Kanuri, G.; Ziegenhardt, D.; Bergheim, I. Beer Is Less Harmful for the Liver than Plain Ethanol : Studies in Male Mice Using a Binge-Drinking Model. Alcohol Alcohol. 2015, 50, 493–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landmann, M.; Sellmann, C.; Engstler, A.J.; Ziegenhardt, D.; Jung, F.; Brombach, C.; Bergheim, I. Hops (Humulus lupulus) Content in Beer Modulates Effects of Beer on the Liver after Acute Ingestion in Female Mice. Alcohol Alcohol. 2017, 52, 48–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osorio-Paz, I.; Brunauer, R.; Alavez, S. Beer and Its Non-Alcoholic Compounds in Health and Disease. Crit. Rev. Food Sci. Nutr. 2020, 60, 3492–3505. [Google Scholar] [CrossRef]
- Griswold, M.G.; Fullman, N.; Hawley, C.; Arian, N.; Zimsen, S.R.M.; Tymeson, H.D.; Venkateswaran, V.; Tapp, A.D.; Forouzanfar, M.H.; Salama, J.S.; et al. Alcohol Use and Burden for 195 Countries and Territories, 1990–2016: A Systematic Analysis for the Global Burden of Disease Study 2016. Lancet 2018, 392, 1015–1035. [Google Scholar] [CrossRef] [Green Version]
- Ferk, F.; Huber, W.W.; Filipič, M.; Bichler, J.; Haslinger, E.; Mišík, M.; Nersesyan, A.; Grasl-Kraupp, B.; Žegura, B.; Knasmüller, S. Xanthohumol, a Prenylated Flavonoid Contained in Beer, Prevents the Induction of Preneoplastic Lesions and DNA Damage in Liver and Colon Induced by the Heterocyclic Aromatic Amine Amino-3-Methyl-Imidazo [4, 5-f ] Quinoline (IQ). Mutat. Res. / Fundam. Mol. Mech. Mutagen. 2010, 691, 17–22. [Google Scholar] [CrossRef]
- Arjomandnejad, M.; Muhammadnejad, A.; Haddadi, M.; Sherkat-Khameneh, N.; Rismanchi, S.; Amanpour, S.; Muhammadnejad, S. Hela Cell Line Xenograft Tumor as a Suitable Cervical Cancer Model: Growth Kinetic Characterization and Immunohistochemis-Try Array. Arch. Iran. Med. 2014, 17, 273–277. [Google Scholar]
- Rocha-Guzmán, N.E.; Gallegos-Infante, J.A.; González-Laredo, R.F.; Reynoso-Camacho, R.; Ramos-Gómez, M.; Garcia-Gasca, T.; Rodríguez-Muñoz, M.E.; Guzmán-Maldonado, S.H.; Medina-Torres, L.; Lujan-García, B.A. Antioxidant Activity and Genotoxic Effect on HeLa Cells of Phenolic Compounds from Infusions of Quercus Resinosa Leaves. Food Chem. 2009, 115, 1320–1325. [Google Scholar] [CrossRef]
- Ruginǎ, D.; Sconţa, Z.; Leopold, L.; Pintea, A.; Bunea, A.; Socaciu, C. Antioxidant Activities of Chokeberry Extracts and the Cytotoxic Action of Their Anthocyanin Fraction on Hela Human Cervical Tumor Cells. J. Med. Food 2012, 15, 700–706. [Google Scholar] [CrossRef] [Green Version]
- Ferk, F.; Mišík, M.; Nersesyan, A.; Pichler, C.; Jäger, W.; Szekeres, T.; Marculescu, R.; Poulsen, H.E.; Henriksen, T.; Bono, R.; et al. Impact of Xanthohumol (a Prenylated Flavonoid from Hops) on DNA Stability and Other Health-Related Biochemical Parameters: Results of Human Intervention Trials. Mol. Nutr. Food Res. 2016, 60, 773–786. [Google Scholar] [CrossRef] [PubMed]
- Varela-Rey, M.; Woodhoo, A.; Maria-Luz, M.-C.; Mato, J.; Lu, S. Alcohol, {DNA} Methylation, and Cancer. Alcohol Res. 2013, 35, 25–35. [Google Scholar] [CrossRef]
- Ratna, A.; Mandrekar, P. Alcohol and Cancer : Mechanisms and Therapies. Biomolecules 2017, 7, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theodoratou, E.; Timofeeva, M.; Li, X.; Meng, X.; Ioannidis, J.P.A. Nature, Nurture and Cancer Risks : Genetic and Nutritional Contributions to Cancer. Annu. Rev. Nutr. 2017, 37, 293–320. [Google Scholar] [CrossRef]
- Caulfeild, J.; Hong, S.; Teufel, K.; Trevithick, J.R.; Vinson, J.A.; Derksen, T.; Bocksch, L.; Trevithick, C.C.; Rahman, F.; Hirst, M.; et al. Is Ethanol an Important Antioxidant in Alcoholic Beverages Associated with Risk Reduction of Cataract and Atherosclerosis? Redox Rep. 2004, 4, 89–93. [Google Scholar] [CrossRef]
- Arts, M.J.T.J.; Haenen, G.R.M.M.; Voss, H.; Bast, A. Masking of Antioxidant Capacity by the Interaction of Flavonoids with Protein. Food Chem. Toxicol. 2001, 39, 787–791. [Google Scholar] [CrossRef]
- Belščak, A.; Komes, D.; Horžić, D.; Ganić, K.K.; Karlović, D. Comparative Study of Commercially Available Cocoa Products in Terms of Their Bioactive Composition. Food Res. Int. 2009, 42, 707–716. [Google Scholar] [CrossRef]
- Ozdal, T.; Capanoglu, E.; Altay, F. A Review on Protein-Phenolic Interactions and Associated Changes. Food Res. Int. 2013, 51, 954–970. [Google Scholar] [CrossRef]
- du Sert, N.P.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; Emerson, M.; et al. Reporting Animal Research: Explanation and Elaboration for the Arrive Guidelines 2.0. PLoS Biol. 2020, 18, 1–65. [Google Scholar] [CrossRef]
- World Health Organization WHO Expert Committee on Biological Standardization, Annex 3: Reccomendations for the Evaluation of Animal Cell Cultures as Substrates for the Manufacture of Biological Medicinal Products and for the Characterization of Cell Banks. World Health Organ. Tech. Rep. Ser. 2013, 79–187.
- Krofta, K. The Content of Hop Prenylflavonoids in Czech and Foreign Beers. Kvas. Prum. 2010, 56, 2–9. [Google Scholar] [CrossRef] [Green Version]
- Cayman Chemical Company Product Information Xanthohumol. Available online: https://cdn.caymanchem.com/cdn/insert/15399.pdf (accessed on 7 April 2021).
- Enzo Life Sciences Xanthohumol. Available online: https://www.enzolifesciences.com/alx-350-280/xanthohumol/ (accessed on 12 August 2020.).
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of ‘“Antioxidant Power”’. The FRAP Assay. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Re, R.; Pellegrini, N.; Proteggenete, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant Activity Applying an Improved ABTS Radical Cation Decolorization Assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Effect Size—A Quick Guide. Available online: https://www.spss-tutorials.com/effect-size/ (accessed on 3 January 2023).
- Vanhoecke, B.W.; Delporte, F.; Braeckel, E.V.A.V.A.N.; Heyerick, A.; Depypere, H.T.; Nuytinck, M.; Keukeleire, D.D.E.; Bracke, M.E. A Safety Study of Oral Tangeretin and Xanthohumol Administration to Laboratory Mice. In Vivo 2005, 19, 103–107. [Google Scholar] [PubMed]
- Dorn, C.; Bataille, F.; Gaebele, E.; Heilmann, J.; Hellerbrand, C. Xanthohumol Feeding Does Not Impair Organ Function and Homoeostasis in Mice. Food Chem. Toxicol. 2010, 48, 1890–1897. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Éliás, A.J.; Balogh, L.; Brányik, T.; Mák, E.; Csajbókné Csobod, É.; Veresné Bálint, M.; Benedek, C. Xanthohumol-Enriched Beer Does Not Exert Antitumorigenic Effects on HeLa Cell Line In Vivo. Molecules 2023, 28, 1070. https://doi.org/10.3390/molecules28031070
Éliás AJ, Balogh L, Brányik T, Mák E, Csajbókné Csobod É, Veresné Bálint M, Benedek C. Xanthohumol-Enriched Beer Does Not Exert Antitumorigenic Effects on HeLa Cell Line In Vivo. Molecules. 2023; 28(3):1070. https://doi.org/10.3390/molecules28031070
Chicago/Turabian StyleÉliás, Anna Júlia, Lajos Balogh, Tomáš Brányik, Erzsébet Mák, Éva Csajbókné Csobod, Márta Veresné Bálint, and Csilla Benedek. 2023. "Xanthohumol-Enriched Beer Does Not Exert Antitumorigenic Effects on HeLa Cell Line In Vivo" Molecules 28, no. 3: 1070. https://doi.org/10.3390/molecules28031070