
New Insights into Conformationally Restricted Carbonic Anhydrase Inhibitors
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemistry
2.2. In Vitro Carbonic Anhydrase Inhibition
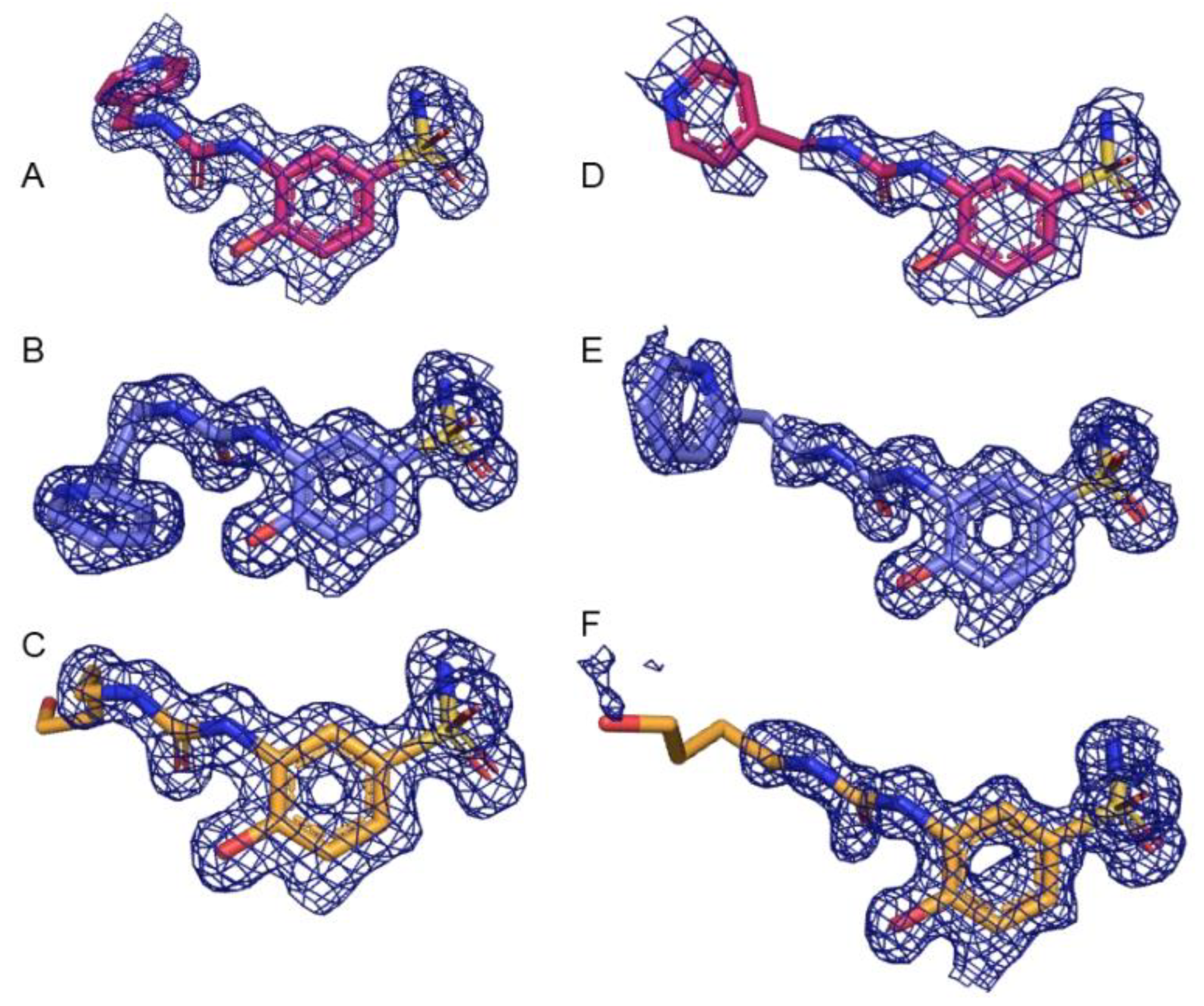
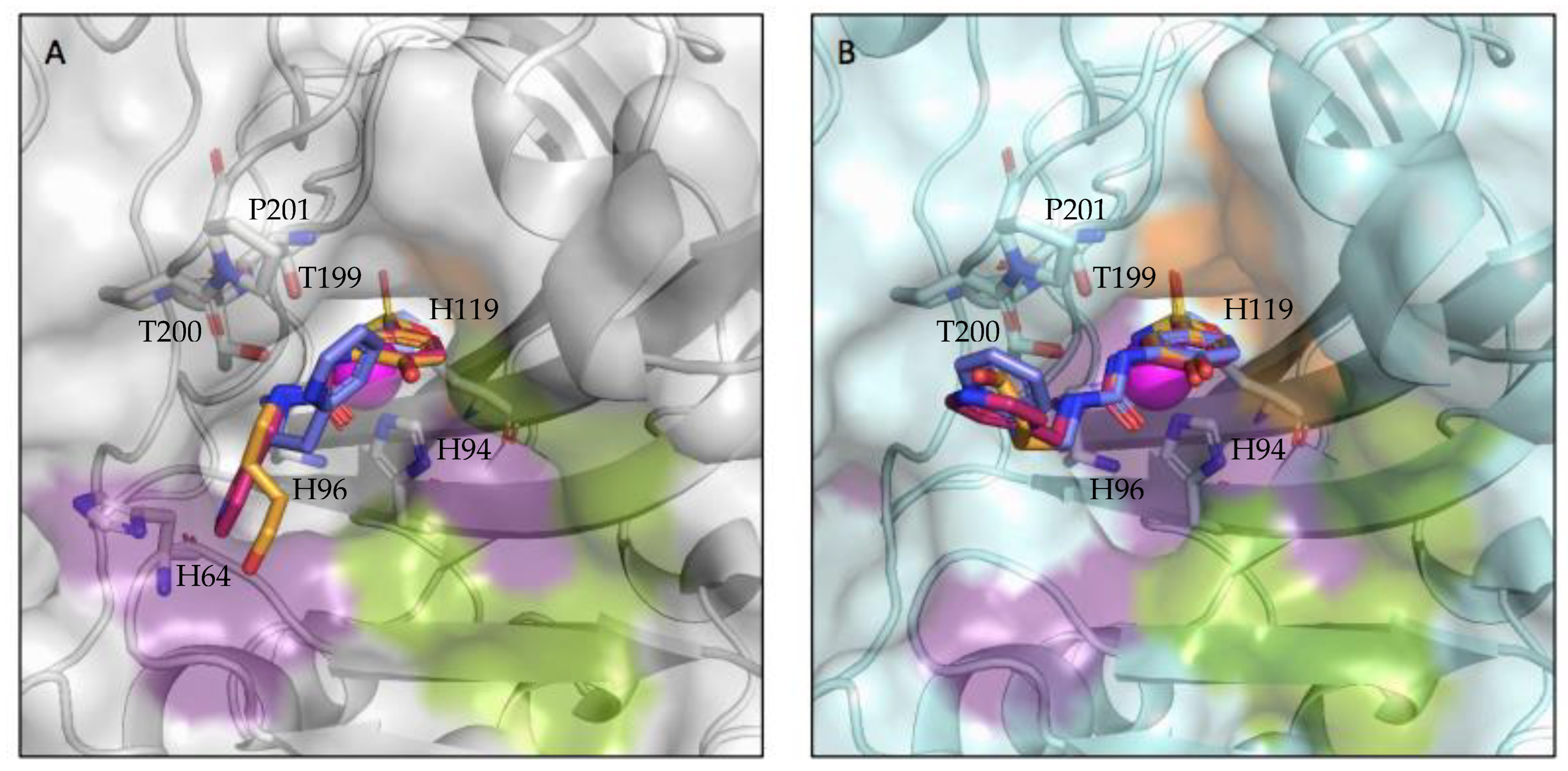
2.3. X-ray Crystallography
3. Experimental Section
3.1. Chemistry
3.2. Carbonic Anhydrase Inhibition
3.3. Carbonic Anhydrase IX-Mimic Design
3.4. Protein Expression and Purification
3.5. Crystal Preparation
3.6. Data Collection and Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bozdag, M.; Carta, F.; Ceruso, M.; Ferraroni, M.; McDonald, P.C.; Dedhar, S.; Supuran, C.T. Discovery of 4-Hydroxy-3-(3-(Phenylureido)Benzenesulfonamides as SLC-0111 Analogues for the Treatment of Hypoxic Tumors Overexpressing Carbonic Anhydrase IX. J. Med. Chem. 2018, 61, 6328–6338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortez-Retamozo, V.; Lauwereys, M.; Hassanzadeh, G.G.; Gobert, M.; Conrath, K.; Muyldermans, S.; De Baetselier, P.; Revets, H. Efficient Tumor Targeting by Single-Domain Antibody Fragments of Camels. Int. J. Cancer 2002, 98, 456–462. [Google Scholar] [CrossRef]
- Testa, C.; Papini, A.M.; Zeidler, R.; Vullo, D.; Carta, F.; Supuran, C.T.; Rovero, P. First Studies on Tumor Associated Carbonic Anhydrases IX and XII Monoclonal Antibodies Conjugated to Small Molecule Inhibitors. J. Enzyme Inhib. Med. Chem. 2022, 37, 592–596. [Google Scholar] [CrossRef] [PubMed]
- Steiner, H.; Jonsson, B.-H.; Lindskog, S. The Catalytic Mechanism of Carbonic Anhydrase. Eur. J. Biochem. 1975, 59, 253–259. [Google Scholar] [CrossRef]
- Frost, S.C.; McKenna, R. Carbonic Anhydrase: Mechanism, Regulation, Links to Disease, and Industrial Applications. In Subcellular Biochemistry; Springer: Berlin/Heidelberg, Germany, 2013; Volume 75. [Google Scholar]
- Alterio, V.; Di Fiore, A.; D’Ambrosio, K.; Supuran, C.T.; De Simone, G. Multiple Binding Modes of Inhibitors to Carbonic Anhydrases: How to Design Specific Drugs Targeting 15 Different Isoforms? Chem. Rev. 2012, 112, 4421–4468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacchiano, F.; Aggarwal, M.; Avvaru, B.S.; Robbins, A.H.; Scozzafava, A.; McKenna, R.; Supuran, C.T. Selective Hydrophobic Pocket Binding Observed within the Carbonic Anhydrase II Active Site Accommodate Different 4-Substituted-Ureido-Benzenesulfonamides and Correlate to Inhibitor Potency. Chem. Commun. 2010, 46, 8371–8373. [Google Scholar] [CrossRef]
- Mboge, M.Y.; Combs, J.; Singh, S.; Andring, J.; Wolff, A.; Tu, C.; Zhang, Z.; McKenna, R.; Frost, S.C. Inhibition of Carbonic Anhydrase Using SLC-149: Support for a Noncatalytic Function of CAIX in Breast Cancer. J. Med. Chem. 2021, 64, 1713–1724. [Google Scholar] [CrossRef]
- Bonardi, A.; Nocentini, A.; Bua, S.; Combs, J.; Lomelino, C.; Andring, J.; Lucarini, L.; Sgambellone, S.; Masini, E.; McKenna, R.; et al. Sulfonamide Inhibitors of Human Carbonic Anhydrases Designed through a Three-Tails Approach: Improving Ligand/Isoform Matching and Selectivity of Action. J. Med. Chem. 2020, 63, 7422–7444. [Google Scholar] [CrossRef] [PubMed]
- Pastorekova, S.; Gillies, R.J. The Role of Carbonic Anhydrase IX in Cancer Development: Links to Hypoxia, Acidosis, and Beyond. Cancer Metastasis Rev. 2019, 38, 65–77. [Google Scholar] [CrossRef]
- Russo, D.; Varricchio, S.; Ilardi, G.; Martino, F.; Di Crescenzo, R.M.; Pignatiello, S.; Scalvenzi, M.; Costa, C.; Mascolo, M.; Merolla, F.; et al. Tissue Expression of Carbonic Anhydrase IX Correlates to More Aggressive Phenotype of Basal Cell Carcinoma. Front. Oncol. 2021, 11, 659332. [Google Scholar] [CrossRef]
- Bozdag, M.; Cravey, L.; Combs, J.; Kota, A.; McKenna, R.; Angeli, A.; Selleri, S.; Carta, F.; Supuran, C.T. Small Molecule Alkoxy Oriented Selectiveness on Human Carbonic Anhydrase II and IX Inhibition. Chem. Med. Chem. 2022, 17, e202200148. [Google Scholar] [CrossRef] [PubMed]
- Berrino, E.; Angeli, A.; Zhdanov, D.D.; Kiryukhina, A.P.; Milaneschi, A.; De Luca, A.; Bozdag, M.; Carradori, S.; Selleri, S.; Bartolucci, G.; et al. Azidothymidine “Clicked” into 1,2,3-Triazoles: First Report on Carbonic Anhydrase–Telomerase Dual-Hybrid Inhibitors. J. Med. Chem. 2020, 63, 7392–7409. [Google Scholar] [CrossRef] [PubMed]
- Khalifah, R.G. The Carbon Dioxide Hydration Activity of Carbonic Anhydrase. I. Stop-Flow Kinetic Studies on the Native Human Isoenzymes B and C. J. Biol. Chem. 1971, 246, 2561–2573. [Google Scholar] [CrossRef] [PubMed]
- Genis, C.; Sippel, K.H.; Case, N.; Cao, W.; Avvaru, B.S.; Tartaglia, L.J.; Govindasamy, L.; Tu, C.; Agbandje-McKenna, M.; Silverman, D.N.; et al. Design of a Carbonic Anhydrase IX Active-Site Mimic To Screen Inhibitors for Possible Anticancer Properties. Biochemistry 2009, 48, 1322–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinard, M.A.; Boone, C.D.; Rife, B.D.; Supuran, C.T.; McKenna, R. Structural Study of Interaction between Brinzolamide and Dorzolamide Inhibition of Human Carbonic Anhydrases. Bioorg. Med. Chem. 2013, 21, 7210–7215. [Google Scholar] [CrossRef] [PubMed]
- Pinard, M.A.; Mahon, B.; McKenna, R. Probing the Surface of Human Carbonic Anhydrase for Clues towards the Design of Isoform Specific Inhibitors. BioMed Res. Int. 2015, 2015, e453543. [Google Scholar] [CrossRef] [Green Version]
- Siffert, W.; Gros, G. Carbonic Anhydrase in Human Platelets. Biochem. J. 1984, 217, 727–730. [Google Scholar] [CrossRef] [Green Version]
- Silverman, D.N.; Lindskog, S. The Catalytic Mechanism of Carbonic Anhydrase: Implications of a Rate-Limiting Protolysis of Water. Acc. Chem. Res. 1988, 21, 30–36. [Google Scholar] [CrossRef]
- Kabsch, W. XDS. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Winn, M.D.; Ballard, C.C.; Cowtan, K.D.; Dodson, E.J.; Emsley, P.; Evans, P.R.; Keegan, R.M.; Krissinel, E.B.; Leslie, A.G.W.; McCoy, A.; et al. Overview of the CCP4 Suite and Current Developments. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 235–242. [Google Scholar] [CrossRef]
- Fisher, S.Z.; Maupin, C.M.; Budayova-Spano, M.; Govindasamy, L.; Tu, C.; Agbandje-McKenna, M.; Silverman, D.N.; Voth, G.A.; McKenna, R. Atomic Crystal and Molecular Dynamics Simulation Structures of Human Carbonic Anhydrase II: Insights into the Proton Transfer Mechanism. Biochemistry 2007, 46, 2930–2937. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.D.; Afonine, P.V.; Bunkóczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.-W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A Comprehensive Python-Based System for Macromolecular Structure Solution. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emsley, P.; Cowtan, K. Coot: Model-Building Tools for Molecular Graphics. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The PyMOL Molecular Graphics System, Version 0.9.4 (2015); Schrodinger, LLC: New York, NY, USA, 2015. Available online: pymol.org(accessed on 9 January 2023).
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple Ligand–Protein Interaction Diagrams for Drug Discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KI * (nM) | |||||
|---|---|---|---|---|---|
| Compound | CAI | CA II | CA IX | CA XII | SI (IX/II) |
| 3a | 583.1 | 3182 | 369.1 | 91.3 | 0.12 |
| 3b | 8654 | 914.5 | 307.9 | 506.6 | 0.34 |
| 3c | 4000 | 8026 | 139.8 | 73.8 | 0.017 |
| 3d | 688.6 | 287.8 | 4116 | 659.1 | 14.3 |
| 3e | 9206 | 2995 | 2142 | 628.8 | 0.71 |
| 3f | 8987 | 7741 | 2153 | 505.3 | 0.28 |
| 3g [10] | 89.5 | 463 | 0.30 | 85.1 | 0.0006 |
| 3h [10] | 571 | 331 | 0.24 | 65.9 | 0.0007 |
| 3i | 697.9 | 405.4 | 204.2 | 33.7 | 0.50 |
| 3k [10] | 798 | 280 | 215 | 47.1 | 0.77 |
| 3j | 315.5 | 75.9 | 34.7 | 238.6 | 0.46 |
| AAZ | 250.0 | 12.1 | 25.8 | 5.7 | 2.1 |
| Compound | 3g | 3h | 3j | |||
|---|---|---|---|---|---|---|
| Protein | CA II | CA IX-Mimic | CA II | CA IX-Mimic | CA II | CA IX-Mimic |
| Resolution Range (Å) | 25.1-1.4 (1.5-1.4) | 28.8-2.0 (2.1-2.0) | 25.1-1.4 (1.5-1.4) | 31.9-1.3 (1.4-1.3) | 30.5-1.4 (1.5-1.4) | 39.8-1.1 (1.2-1.1) |
| Space Group | P 1 21 1 | P 1 21 1 | P 1 21 1 | P 1 21 1 | P 1 21 1 | P 1 21 1 |
| Unit Cell: a, b, c (Å) α, β, γ (°) | 42.0, 40.8, 71.1 90, 104.1, 90 | 42.5, 41.6, 72.5 90, 103.5, 90 | 42.0, 40.9, 71.4 90, 104.2, 90 | 41.9, 41.2, 71.9 90, 103.8, 90 | 42.3, 41.1, 71.9 90, 104.2, 90 | 42.2, 41.4, 72.2 90, 103.6, 90 |
| Total Reflections | 123,074 (11,706) | 30,931 (3,110) | 130,379 (12,291) | 189,486 (17,491) | 134,739 (12,839) | 279,296 (19,725) |
| Redundancy | 3.4 (3.3) | 1.9 (1.9) | 3.3 (3.3) | 3.3 (3.1) | 3.3 (3.3) | 3.3 (2.9) |
| Completeness (%) | 96 (93) | 94 (97) | 97 (93) | 100 (98) | 99 (96) | 92 (71) |
| Mean I/σ (I) | 17.4 (1.9) | 7.6 (2.6) | 15.2 (1.9) | 14.7 (1.8) | 18.6 (1.8) | 10.9 (1.3) |
| R-Merge/R-Symm | 0.04 (0.52) | 0.08 (0.33) | 0.04 (0.59) | 0.04 (0.57) | 0.04 (0.57) | 0.05 (0.76) |
| R-Work | 0.17 (0.30) | 0.19 (0.27) | 0.17 (0.27) | 0.16 (0.29) | 0.16 (0.41) | 0.17 (0.24) |
| R-Free | 0.19 (0.32) | 0.25 (0.35) | 0.19 (0.31) | 0.18 (0.29) | 0.18 (0.43) | 0.17 (0.23) |
| No. of non-H atoms: CA, Inhibitor, water | 2080, 33, 291 | 2042, 31, 36 | 2062, 24, 344 | 2061, 36, 408 | 2062, 29, 324 | 2061, 27, 208 |
| Ramachandran Statistics: Favored, Allowed, Outliers (%) | 97.3, 2.7, 0.0 | 96.1, 3.9, 0.0 | 96.9, 3.1, 0.0 | 96.1, 3.9, 0.0 | 97.6, 2.4, 0.0 | 96.9, 3.1, 0.0 |
| Average B-factors: Overall, CA, Inhibitor, solvent (Å2) | 23.0, 21.9, 30.2, 31.7 | 27.4, 27.2, 40.7, 26.3 | 21.2, 20.0, 17.7, 30.8 | 18.3, 16.3, 27.8, 29.8 | 19.8, 18.1, 31.3, 31.2 | 18.6, 17.1, 25.3, 31.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Combs, J.; Bozdag, M.; Cravey, L.D.; Kota, A.; McKenna, R.; Angeli, A.; Carta, F.; Supuran, C.T. New Insights into Conformationally Restricted Carbonic Anhydrase Inhibitors. Molecules 2023, 28, 890. https://doi.org/10.3390/molecules28020890
Combs J, Bozdag M, Cravey LD, Kota A, McKenna R, Angeli A, Carta F, Supuran CT. New Insights into Conformationally Restricted Carbonic Anhydrase Inhibitors. Molecules. 2023; 28(2):890. https://doi.org/10.3390/molecules28020890
Chicago/Turabian StyleCombs, Jacob, Murat Bozdag, Lochlin D. Cravey, Anusha Kota, Robert McKenna, Andrea Angeli, Fabrizio Carta, and Claudiu T. Supuran. 2023. "New Insights into Conformationally Restricted Carbonic Anhydrase Inhibitors" Molecules 28, no. 2: 890. https://doi.org/10.3390/molecules28020890