
Ca2+-Sensitive Potassium Channels

1
Biomedical Research Administration, Research Centre, King Fahad Medical City, Riyadh 12231, Saudi Arabia
2
Clinical Pharmacy Administration, King Fahad Medical City, Riyadh 12231, Saudi Arabia
*
Author to whom correspondence should be addressed.
Molecules 2023, 28(2), 885; https://doi.org/10.3390/molecules28020885
Submission received: 23 December 2022
/
Revised: 7 January 2023
/
Accepted: 10 January 2023
/
Published: 16 January 2023
Abstract
:The Ca2+ ion is used ubiquitously as an intracellular signaling molecule due to its high external and low internal concentration. Many Ca2+-sensing ion channel proteins have evolved to receive and propagate Ca2+ signals. Among them are the Ca2+-activated potassium channels, a large family of potassium channels activated by rises in cytosolic calcium in response to Ca2+ influx via Ca2+-permeable channels that open during the action potential or Ca2+ release from the endoplasmic reticulum. The Ca2+ sensitivity of these channels allows internal Ca2+ to regulate the electrical activity of the cell membrane. Activating these potassium channels controls many physiological processes, from the firing properties of neurons to the control of transmitter release. This review will discuss what is understood about the Ca2+ sensitivity of the two best-studied groups of Ca2+-sensitive potassium channels: large-conductance Ca2+-activated K+ channels, KCa1.1, and small/intermediate-conductance Ca2+-activated K+ channels, KCa2.x/KCa3.1.
1. Introduction
Ca2+ is essential for the survival and functioning of all living cells [1]. It is an important signaling molecule that crosses membranes, acting as a homeostatic regulator for both intracellular and extracellular fluid [2]. Ca2+ regulates a wide range of functions in all types of cells, involving several ion channels [1]. Ca2+ ions offer ultimate properties in size, charge, availability, and ionization potential, allowing them to flow rapidly, yet simultaneously, bind deeply. Thus calcium might be considered the prominent intracellular signaling molecule, as it is involved in numerous physiological processes such as neurotransmission, muscle contraction, the regulation of gene expression, fertilization, and mitosis [3,4,5]. Unique among these Ca2+ regulatory processes are the Ca2+ sensitive ion channels or Ca2+-activated ion channels. The gating of these channels is modulated by Ca2+-sensing proteins such as calmodulin CaM [3,6]. CaM is characterized by its ability to bind and release Ca2+ over the physiological Ca2+ concentrations. It undergoes a significant conformational change in Ca2+ binding and consequently regulates these specific ion channels to modify their function. Therefore, the physiological roles and functions of these channels can be affected by changes in intracellular Ca2+ concentration. Some Ca2+-sensitive ion channels include Ca2+-activated anion (Cl−) channels, Ca2+-regulated nonselective cation channels, and Ca2+-activated K+ channels. Here we will elaborate on the Ca2+-sensing mechanisms of the Ca2+-activated K+ channels.
Potassium channels are the most diverse and abundantly expressed ion channels in living organisms. These channels are expressed in the most excitable and non-excitable cells. They perform various vital functions and can be classified into several different groups. Among these groups are the Ca2+-activated potassium channels (KCa channels), a large family of potassium channels activated by rises in cytosolic Ca2+ [7,8]. Over the last two decades, advances have been made in KCa channel research, including the channels’ functions, expression, pharmacology, and genetic mutations associated with channelopathies. The family of KCa channels shares a typical functional role by coupling the increase in intracellular Ca2+ concentration to the hyperpolarization of the membrane potential. Thus, KCa channels are critical for maintaining K+ homeostasis and cell volume. Additionally, KCa channels modulate several physiological processes, from neuron firing properties to transmitter release control [8].
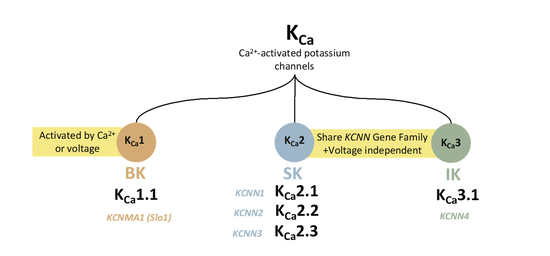
KCa channels can be divided into three main subfamilies based on their single-channel conductance, SK or KCa2 (small conductance; ∼4–14 pS), IK or KCa3.1 (intermediate conductance; ∼32–39 pS), and BK or KCa1.1 (large conductance; ∼200–300 pS) channels [8,9,10]. K Ca1.1 channels are activated by intracellular Ca2+ and membrane voltage synergistically. However, K Ca2x and K Ca3.1 are gated solely by internal Ca2+ ions and are more sensitive to Ca2+ than K Ca1.1 channels. KCa2 includes three small-conductance KCa2-channel subtypes, KCa2.1 (SK1), KCa2.2 (SK2), and KCa2.3 (SK3) [8,11,12]. The Ca2+-binding sites and Ca2+-binding affinities are significantly different among these channel groups. KCa1.1 directly binds Ca2+ and has a very low Ca2+-binding affinity associated with the regulator of conductance of K+ (RCK) Ca2+-binding domains (1–11 μM) [7,8]. In contrast, KCa2.x and KCa3.1 channels share the calmodulin-mediated gating mechanism with a high Ca2+-binding affinity due to the constitutive binding of calmodulin (300–600 nM) [13,14,15]. KCa1.1 and KCa2/3 channels show minimal sequence homology. The latter’s S4 segment comprises fewer charged residues than the S4 segment of KCa1.1, which results in a lack of voltage dependence in KCa2/3 channels, enabling them to remain open at negative membrane potentials and thus hyperpolarize the membrane toward values near the K+ equilibrium potential [11,14]. Each type of KCa channel shows distinct pharmacology, and the activity of each hyperpolarizes the membrane potential. Here we will discuss these unique potassium ion channels and their sensitivity to Ca2+, aiming to understand the mechanisms behind Ca2+-dependent regulation.
2. Ca2+-Activated KCa1.1 Channels
Large-conductance Ca2+-activated K+ (BKCa) channels are activated under dual control by both calcium and the membrane voltage (membrane depolarization). The calcium dependence of these channels changes steeply according to membrane potentials, with the calcium Kd in the micromolar range at resting membrane potentials (~–60 mV) but in the nanomolar range at depolarized potentials (+20 to +40 mV). Remarkably, these channels can open in the absence of calcium, and it seems that the effects of calcium and membrane potential are independent processes that both increase the open probability. While the voltage dependence of these channels is quite weak compared to the other purely voltage-gated potassium channels, the voltage dependence of KCa1.1 enables them to act as coincidence detectors, which is central to their physiological function [7]. These channels are expressed on the plasma membrane as tetramers of α subunits. The gene encoding the α subunit (slo; KCNMA1) of KCa1.1 channels (KCa1.1; slo1) was cloned in the early 1990s from Drosophila. KCa1.1 channels are widely distributed in various cells and regulate Ca2+ influx and many Ca2+-dependent physiological processes [8,16,17].
2.1. Expression and Physiology of KCa1.1 Channels
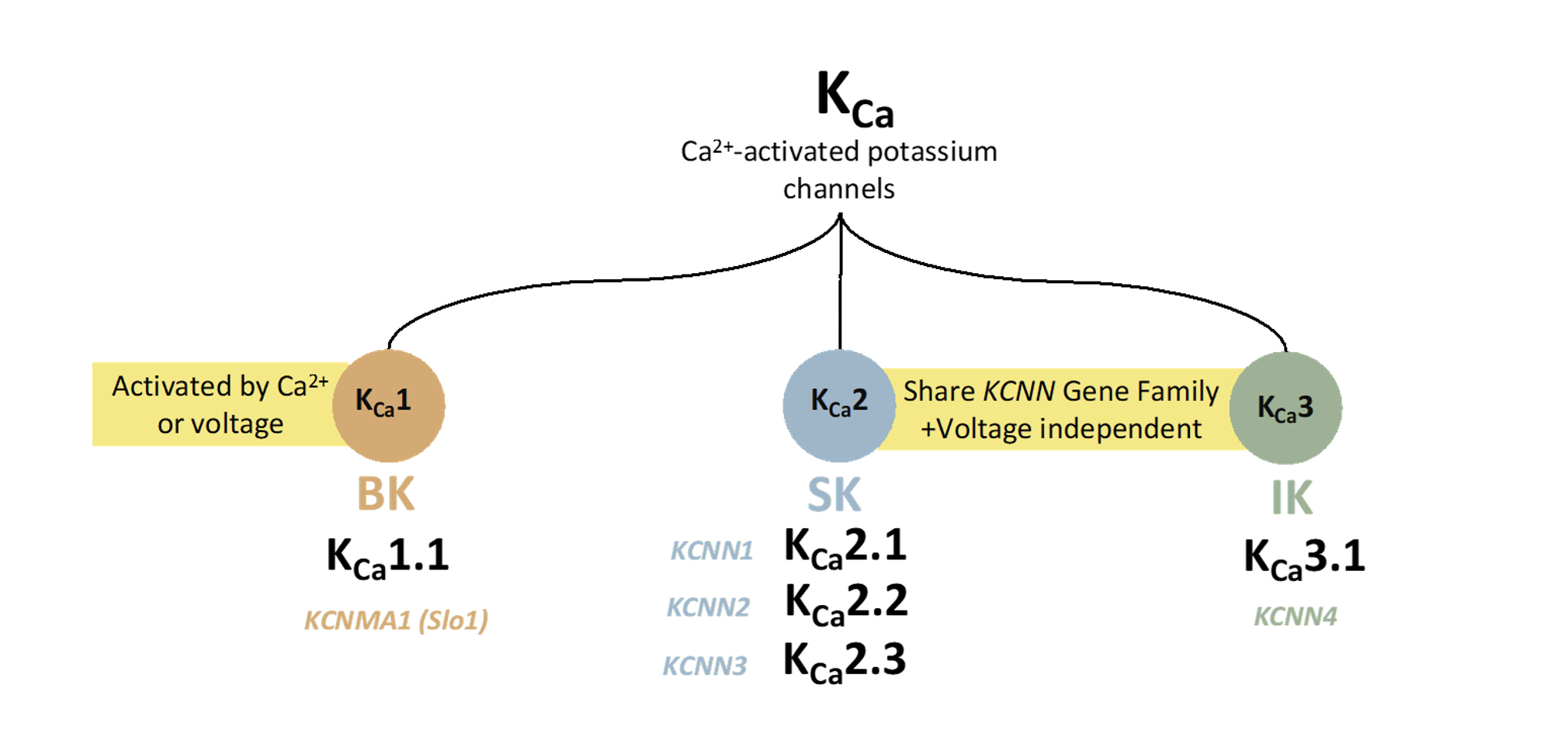
In neurons, the activation of KCa1.1 channels repolarizes the membrane and reduces Ca2+ influx into the cells [7,18,19]. KCa1.1 channels participate in numerous physiological processes via the modulation of membrane excitability and Ca2+ homeostasis [18]. Examples include modulating neurotransmitter release, regulating vascular and respiratory tone, neurovascular coupling [7], endocrine secretion, and urinary bladder tone.
KCa1.1 channels control hormonal secretion by altering the duration and frequency of action potentials [20,21]. The main functions of KCa1.1 channels in neurons are to generate fast afterhyperpolarization after an action potential [22]. In neurovascular coupling, KCa1.1 channels umpire most of the dilation and the entire vasoconstriction in astrocytic end feet [18,23]. In retinal circulation, KCa1.1channels are contributed to the regulation of retinal blood flow via the action of several vasodilators in the endothelium and vascular smooth muscle. A recent study has shown that administering a KCa1.1 channel activator (BMS-191011) to male Wistar rats improved retinal circulation [18,24,25]. In the urinary system, KCa1.1 channels have been reported in various renal cell types, such as urinary bladder smooth muscle cells [26], glomerular mesangial cells [27], afferent arterioles [28], and podocytes in the Bowman’s capsule [29].
It has been demonstrated that KCa1.1 channels provide negative feedback, preventing contractions induced by agonists. Moreover, several segments of the distal convoluted tubules of the nephron have been shown to express KCa1.1 channels that are suggested to contribute to the volume regulation in the distal convoluted tubules of the nephron [26,28].
The list continues with numerous studies identifying enormous regulatory physiological mechanisms. KCa1.1 channels may serve as a potential target (Figure 1).
2.2. KCa1.1 Channels Structure
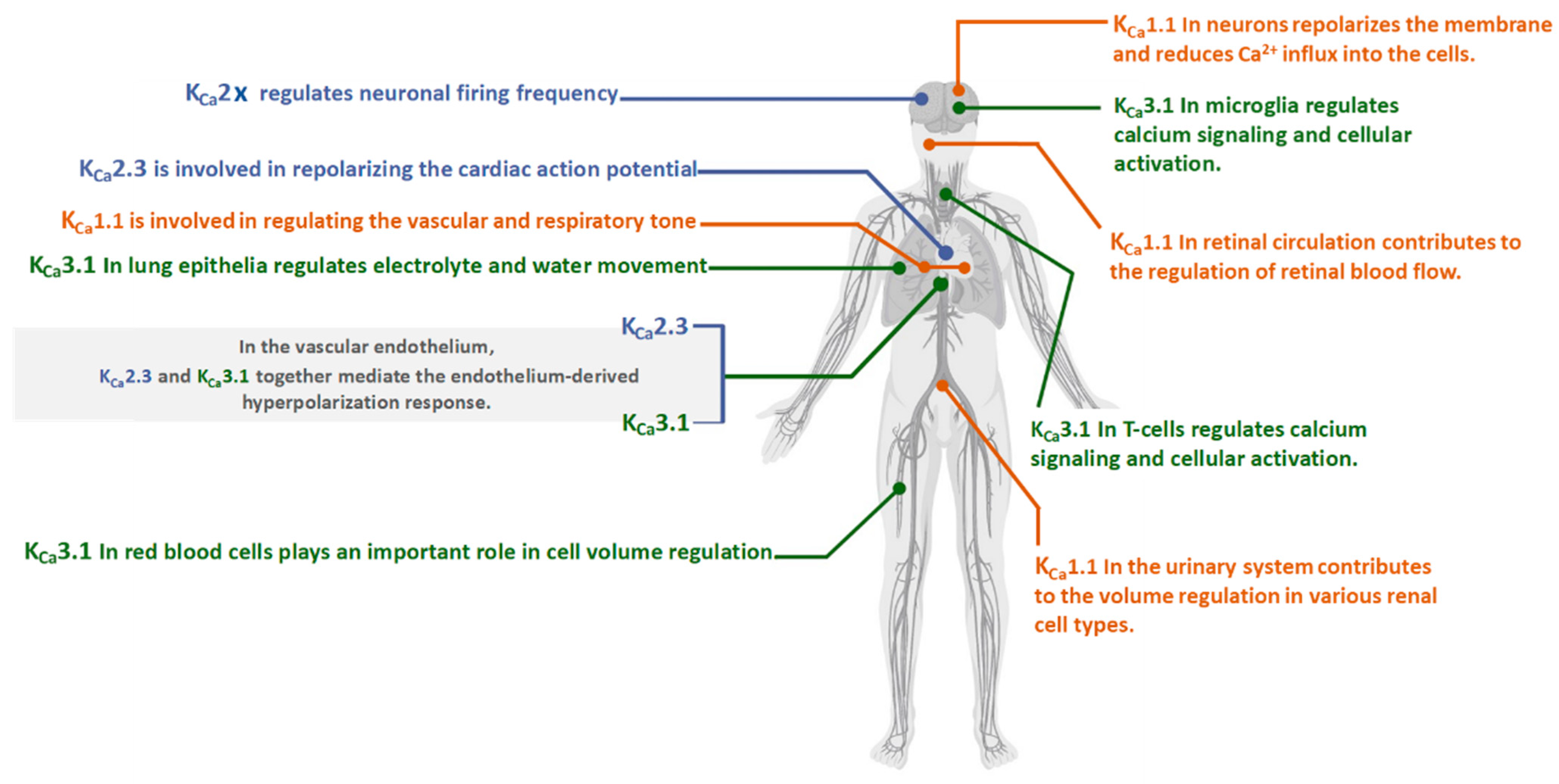
KCa1.1 channels are composed of four pore-forming subunits (α) encoded by the Slo1 gene or KCNMA1 in mammals. The Slo1 gene undergoes alternative splicing leading to a high degree of functional diversity in the KCa1.1 channels [16,30]. Each α (Slo1) subunit contains three main domains: a voltage sensor domain (S0–S4), a pore-gate domain (S5–S6), and a long intracellular C-terminal cytosolic region, which functions as a Ca2+ sensor domain. KCa1.1 channels are also co-assembling and prominently impacted by diverse auxiliary subunits, including β (β1-4), γ (γ1-4), and LINGO1 subunits that give rise to the existence of distinct KCa1.1 channel phenotypes with diverse functionality [16,29,31,32].
The Ca2+ sensor domain comprises two non-identical domains (i.e., RCK1 and RCK2), which contain high-affinity binding Ca2+ sites (Ca2+ bowl) that have been implicated in the direct gating of the channel [16,33,34]. The KCa1.1-β-subunits have been shown to affect the Ca2+ sensitivity of the KCa1.1 channel gating [35,36]. Thus, due to these structural features, the KCa1.1 channel is characterized by unique biophysical properties, including ion permeation, gating, and modulation by diverse ligands and intracellular molecules.
Cryogenic electron microscopy (cryo-EM) [37] has begun to provide crucial structural and biophysical insights into the KCa1.1 channel gating. Structure–function studies of KCa1.1 channels are needed to provide a better understanding of these channels in search of novel compounds to treat diverse KCa1.1-associated pathologies [17,38] (Figure 2A).
2.3. Channelopathies of KCa1.1 Channels
Channelopathies are conditions caused by pathogenic alterations in ion channel activity that disrupt homeostasis and physiological functions [39,40]. With advances in whole-exome sequencing (WES), various monogenetic channelopathies are now diagnosable. However, the molecular basis of these mutations and how they produce clinical phenotypes remains unclear. Because of the small number of patients and the lack of genetic pedigree analysis, we have a limited understanding of channelopathies [41]. Alterations in the activity of KCa1.1 channels were demonstrated in several channelopathies. Human KCNMA1 mutations are primarily linked with neurological conditions, such as seizures, developmental delay, movement disorders, and intellectual disability. KCNMA1 mutations are also associated with several other pathologies, including diabetes [42], atherosclerosis [43], hypertension [44], and cardiac hypertrophy [45]. These mutations involve gain-of-function (GOF) and loss-of-function (LOF) alterations in KCa1.1 channel activity, together with several variants of unknown significance (VUS). Evidence suggests that the mutation alterations of the KCa1.1 channel may associate with semi-distinct patient symptoms, such as paroxysmal nonkinesigenic dyskinesia (PNKD) with GOF and ataxia with LOF. KCa1.1 channel dysfunction can also lead to urinary incontinence and an overactive bladder [46]. Data has already suggested that the deletion of the KCa1.1 gene has been associated with progressive hearing loss [47]. Other results [48] indicated that losing the KCa1.1 channel leads to erectile dysfunction. Though most KCNMA1 mutations are de novo in origins, additional evidence is needed to establish causality in most cases. Additionally, GOF and LOF in KCa1.1 channels are linked with overlapping symptoms. However, it is unclear whether selective agonists or antagonists that correct the level of KCa1.1 channel activity will produce the desired outcome on neuronal activity [39,40].
3. Ca2+-Activated KCa 2.x Channels
KCa2.x channels are members of the voltage-insensitive calcium-activated potassium channel family that are stimulated by the elevation of the cytosolic calcium concentration. Upon activation, KCa2.x channels allow K+ ions to leave the cell as a function of the difference between the depolarized cell and the K+ equilibrium potentials [49]. KCa2.x subunits are encoded by the KCNN1 (KCa2.1; SK1), KCNN2 (KCa2.2; SK2), and KCNN3 (KCa2.3; SK3) genes [50], while IKα (KCa3.1; SK4) are encoded by the KCNN4 gene [51].
3.1. Expression and Physiology of KCa 2.x Channels
The three subtypes of KCa2.x channels (KCa2.1–3) are expressed in many areas of the central nervous system and are involved in afterhyperpolarization upon activation. In the cerebellum, KCa2.2 is the primary channel subtype expressed in Purkinje cells and has a crucial role in the Purkinje cell peacemaking [52,53]. Furthermore, KCa2.x channels are expressed on the neuronal plasma membrane and are suggested to exert neuroprotective effects by modulating the firing pattern of dopaminergic neurons. In dopaminergic neurons, KCa2.x channels were determined on the membrane of mitochondria and, upon activation, were suggested to prevent mitochondrial dysfunction [54]. In the cardiovascular system, KCa2.x subtypes are expressed in the atrial and ventricular walls and play essential roles in regulating the activity of atrial myocytes. KCa2.3 and KCa3.1 are predominant subtypes expressed in endothelial cells and play a crucial role in vasodilation, which is mediated by an endothelium-derived hyperpolarizing factor (EDHF) [55,56]. The diversity of KCa2.x physiological roles are widened due to numerous splicing variants in different tissues (Figure 1) [57].
3.2. KCa 2.x Channels Structure
All three KCa2.x subtypes and KCa3.1 form tetrameric channels and are between 553 and 580 amino acids long. The α subunits of these channels comprise six transmembrane helices (S1–S6), the pore region between helices S5 and S6), and cytosolic N- and C-terminal domains. Their biophysical characteristics are their independence from membrane voltage because they have lost most of the positively charged residues commonly associated with voltage-dependent gating [58]. Low intracellular Ca2+ concentrations activate KCa2.x channels through a unique calmodulin (CaM) gating mechanism. (Figure 2B).
3.3. Channelopathies of KCa 2.x Channels
The discovery of mutations in genes encoding K+ channel subunits has provided unique insights into the pathophysiology of human disorders affecting the central nervous system, heart, kidney, and other organs [59]. Decreased or increased activity of KCa channels caused by loss-of-function (LOF) and gain-of-function (GOF) variants in the corresponding genes, respectively, underlies a broad spectrum of human channelopathies [60,61]. There are no known genetic disorders associated with KCa2.1 channels in humans. While neurodevelopmental disorders such as cerebellar ataxia and tremor are associated with loss-of-function KCa2.2 mutations [59]. The GOF mutations of KCa2.3 are linked with the Zimmermann–Laband syndrome (ZLS) [60,62,63,64] and have also been associated with an idiopathic non-cirrhotic portal hypertension (INCPH) [60,62,65], In one study [66], CAG repeat polymorphism in KCNN3 was linked to schizophrenia. On the other hand, another study [67] found that a mutation of the KCa2.3 channel gene (L283fs287X) results in the deletion of the protein N-terminal region identified in schizophrenic patients. These resulting mutant KCa2.3 channels were found to dominantly suppress KCa2.3 channel currents [67] (LOF mutation). It has been reported that GOF mutations of KCa3.1 are linked with a subset of hereditary xerocytosis (HX) (OMIM 194380), which is an autosomal dominant congenital hemolytic anemia characterized by erythrocyte dehydration [60,68,69].
4. Ca2+- Sensitivity of KCa Channels
The intracellular Ca2+ concentration and membrane potential are important metabolic parameters for all organisms. Ca2+ is involved in numerous physiological processes, including muscle contraction, the regulation of gene expression, neurotransmission, mitosis, and fertilization.
Ca2+-sensitive ion channels allow for crosstalk between chemical and electrical systems and the feedback control of Ca2+ entry into the cells [5,10]. The unique Ca2+-activated potassium channels are best known for enabling changes in intracellular Ca2+ concentration to influence neuronal firing patterns and the strength of muscle contraction. KCa2.x/KCa3.1 are voltage-independent and are activated by low intracellular Ca2+ concentrations. Remarkably, KCa 2.x and KCa3.1 display similar Ca2+ sensing properties (Ca2+ dose-response relationships) and respond rapidly to changes in Ca2+ with time constants of 5–15 ms [13].
Ca2+-binding protein calmodulin (CaM) serves as the KCa2.x and KCa3.1 channel’s Ca2+ sensor [10,70] (Figure 2B). KCa1.1 channels are both Ca2+ and voltage-activated, and their sensitivity to voltage and intracellular Ca2+ are prominently influenced by their association with auxiliary and non-pore-forming modulatory β (β1-4), γ (γ1-4), and LINGO1 subunits [32,71,72]. Site-directed-mutagenesis experiments have led to understanding the KCa1.1 channel’s Ca2+ binding sites. The KCa1.1 structure (Figure 2A) displayed shows that each α subunit contains a pair of RCK domains in the C-terminus portion where the Ca2+ bowl resides. The KCa1.1 Ca2+ gating interface is made by the intersubunit assembly interface, and mutational analyses studies of the putative interacting residues in human BK channels discovered that E955 is a determinant of Ca2+ sensitivity through intersubunit electrostatic interactions. These findings proved that the intersubunit assembly interface contains molecular determinants of Ca2+ sensitivity in KCa1.1 channels [73].
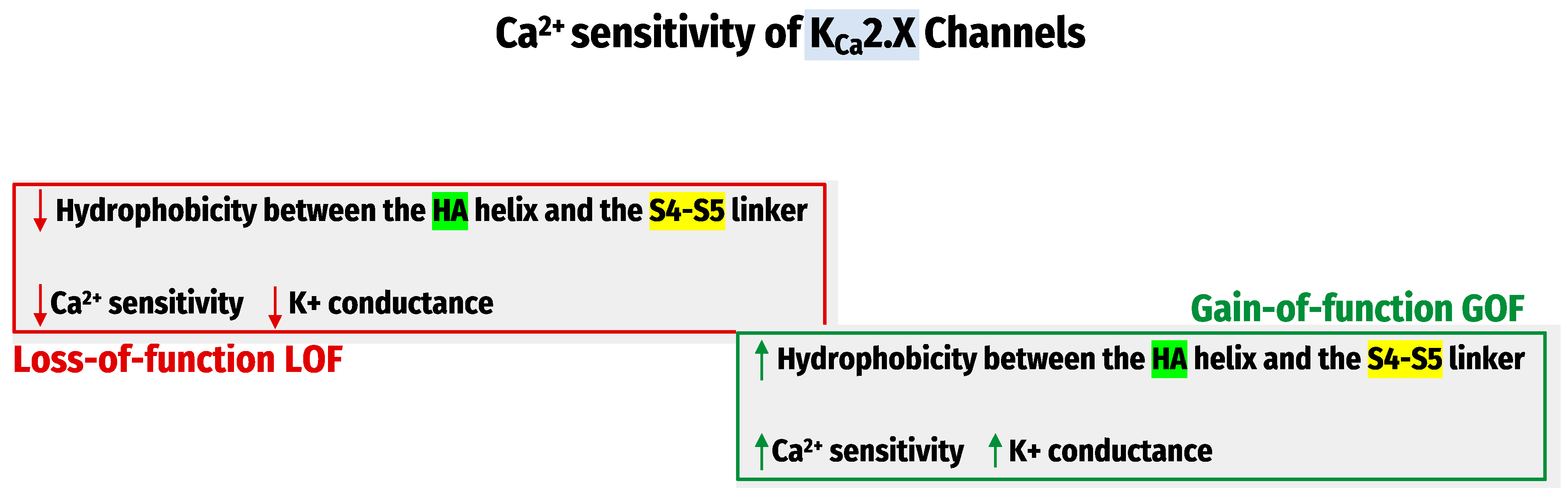
Recently, the Cryo-electron-microscopy structures of the entire KCa1.1 channel [74] have significantly advanced our understanding of Ca2+ sensing mechanisms. Therefore, a tremendous effort has been devoted to developing small molecules targeting KCa channels. These small molecules could modulate KCa channels positively or negatively by influencing the apparent Ca2+ sensitivity of these channels. We have recently investigated how the KCa2.x channel’s apparent Ca2+ sensitivity is regulated and found that the hydrophobic interactions between the HA helix and S4-S5 linker regulate the KCa2.x channel’s apparent Ca2+ sensitivity. The methods utilized were site-directed mutagenesis, patch-clamp recordings, and molecular dynamic (MD) simulations. The observations determined that mutations that decrease hydrophobicity at the HA-S4–S5 interface led to Ca2+ hyposensitivity of KCa2.x channels. The mutation that increases hydrophobicity results in hypersensitivity to Ca2+ [75] (Figure 3). These studies are based on the apparent Ca2+ sensitivity of the amino acid sequences of KCa2.x, which disclosed several differences between their channel subtypes. More studies are needed for the potential drug development for channelopathy disorders.
Genetic mutations encoding KCa channels change their apparent Ca2+ sensitivity and lead to subsequent LOF, a decrease in Ca2+ sensitivity, or GOF, and Ca2+ hypersensitivity of these channels have been directly linked to many human diseases. Loss-of-function KCa2.2 mutations are associated with neurodevelopmental disorders, such as ataxias and tremors [76,77]. Gain-of-function of KCa2.3 mutations is associated with the Zimmermann–Laband syndrome (ZLS) [78,79] and idiopathic non-cirrhotic portal hypertension (INCPH) [80]. In contrast, GOF KCa3.1 mutations are linked with hereditary xerocytosis (HX) [81].
Human KCNMA1-linked channelopathy mutations are primarily associated with neurological conditions, including movement disorders, seizures, and intellectual disability. These mutations comprise GOF and LOF alterations in KCa1.1 channel activity and some variants of unknown significance (VUS). Over the past decade, a great effort has been devoted to the Ca2+ sensing mechanisms of Ca2+-activated K+ channels. Each channel type uses a unique molecular mechanism to regulate its Ca2+ gating. Site-directed mutagenesis, in silico modeling, and electrophysiology have proven to make powerful tools in studying Ca2+-dependent gating that leads to the development of more selective biophysical and pharmacological approaches for the KCa channelopathy.
The hydrophobic interactions between the HA helix and S4–S5 linker regulate the KCa2.x channel’s apparent Ca2+ sensitivity.
5. Summary
Sensitive KCa channels reviewed as Table 1.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to thank the research center at king Fahad medical city. Special thanks go to Ali Alfaiz for his support and contribution. We are grateful to Rawan Orfali and Toleen Alrijraji for their technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nordin, B.E.C. Calcium in health and disease. Interrelat. Between Essent. Met. Ions Hum. Dis. 1997, 14, 81–137. [Google Scholar]
- Williams, R.J.P. Calcium: Outside/inside homeostasis and signalling. Biochim. Et Biophys. Acta BBA-Mol. Cell Res. 1998, 1448, 153–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clapham, D.E. Calcium signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [PubMed] [Green Version]
- Carafoli, P.I.B.E.; Carafoli, E.; Klee, C.B. Calcium as a Cellular Regulator; Oxford University Press: Oxford, UK, 1999. [Google Scholar]
- Carafoli, E. Calcium–a universal carrier of biological signals. FEBS J. 2005, 272, 1073–1089. [Google Scholar] [CrossRef] [PubMed]
- Celio, M.R.; Pauls, T.L.; Schwaller, B. Guidebook to the Calcium-Binding Proteins; Sambrook & Tooze Publication at Oxford University Press: Oxford, UK; New York, NY, USA, 1996. [Google Scholar]
- Faber, E.S.L.; Sah, P. Calcium-Activated Potassium Channels: Multiple Contributions to Neuronal Function. Neuroscientist 2003, 9, 181–194. [Google Scholar] [CrossRef]
- Shah, K.R.; Guan, X.; Yan, J. Structural and Functional Coupling of Calcium-Activated BK Channels and Calcium-Permeable Channels Within Nanodomain Signaling Complexes. Frontiers in Physiology. 2022. Available online: https://www.frontiersin.org/articles/10.3389/fphys.2021.796540 (accessed on 16 August 2022).
- Ishii, T.M.; Silvia, C.; Hirschberg, B. A human intermediate conductance calcium-activated potassium channel. Proc. Natl. Acad. Sci. USA 1997, 94, 11651–11656. [Google Scholar] [CrossRef] [Green Version]
- Cox, D.H. Ca(2+)-regulated ion channels. BMB Rep. 2011, 44, 635–646. [Google Scholar] [CrossRef] [Green Version]
- Wulff, H.; Köhler, R. Endothelial Small- and Intermediate-Conductance KCa Channels: An Update on Their Pharmacology and Usefulness as Cardiovascular Targets. J. Cardiovasc. Pharmacol. 2013, 61, 102–112. [Google Scholar] [CrossRef] [Green Version]
- Nam, Y.-W.; Cui, M.; El-Sayed, N.S. Subtype-selective positive modulation of KCa2 channels depends on the HA/HB helices. Br. J. Pharmacol. 2022, 179, 460–472. [Google Scholar] [CrossRef]
- Xia, X.M.; Fakler, B.; Rivard, A. Mechanism of calcium gating in small-conductance calcium-activated potassium channels. Nature 1998, 395, 503–507. [Google Scholar] [CrossRef]
- Wei, A.D.; Gutman, G.A.; Aldrich, R. International Union of Pharmacology. LII. Nomenclature and molecular relationships of calcium-activated potassium channels. Pharmacol. Rev. 2005, 57, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Fanger, C.M.; Ghanshani, S.; Logsdon, N.J. Calmodulin mediates calcium-dependent activation of the intermediate conductance KCa channel, IKCa1. J. Biol. Chem. 1999, 274, 5746–5754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.-N. Large-conductance Ca2+- activated K+ channels:physiological role and pharmacology. Curr. Med. Chem. 2003, 10, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Latorre, R.; Brauchi, S. Large conductance Ca2+-activated K+ (BK) channel: Activation by Ca2+ and voltage. Biol. Res. 2006, 39, 385–401. [Google Scholar] [CrossRef] [PubMed]
- Vetri, F.; Choudhury, M.S.R.; Pelligrino, D.A. BKCa channels as physiological regulators: A focused review. JRLCR 2014, 7, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Gu, N.; Vervaeke, K.; Storm, J.F. BK potassium channels facilitate high-frequency firing and cause early spike frequency adaptation in rat CA1 hippocampal pyramidal cells. J. Physiol. 2007, 580, 859–882. [Google Scholar] [CrossRef] [PubMed]
- N’Gouemo, P. BKCa channel dysfunction in neurological diseases. Front. Physiol. 2014, 5, 373. [Google Scholar] [PubMed] [Green Version]
- Contet, C.; Goulding, S.P.; Kuljis, D.A. BK Channels in the Central Nervous System. Int. Rev. Neurobiol. 2016, 128, 281–342. [Google Scholar]
- Bond, C.T.; Herson, P.S.; Strassmaier, T. Small conductance Ca2+-activated K+ channel knock-out mice reveal the identity of calcium-dependent afterhyperpolarization currents. J. Neurosci. 2004, 24, 5301–5306. [Google Scholar] [CrossRef] [Green Version]
- Vetri, F.; Qi, M.; Xu, H. Impairment of neurovascular coupling in Type 1 Diabetes Mellitus in rats is prevented by pancreatic islet transplantation and reversed by a semi-selective PKC inhibitor. Brain Res. 2017, 1655, 48–54. [Google Scholar] [CrossRef] [Green Version]
- Nelson, M.T.; Cheng, H.; Rubart, M. Relaxation of Arterial Smooth Muscle by Calcium Sparks. Science 1995, 270, 633–637. [Google Scholar] [CrossRef] [PubMed]
- McGahon, M.K.; Dash, D.P.; Arora, A. Diabetes downregulates large-conductance Ca2+-activated potassium beta 1 channel subunit in retinal arteriolar smooth muscle. Circ. Res. 2007, 100, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Herrera, G.M.; Heppner, T.J.; Nelson, M.T. Regulation of urinary bladder smooth muscle contractions by ryanodine receptors and BK and SK channels. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R60–R68. [Google Scholar] [CrossRef] [PubMed]
- Stockand, J.D.; Sansom, S.C. Large Ca2+-activated K+ channels responsive to angiotensin II in cultured human mesangial cells. Am. J. Physiol. Ren. Physiol. 1994, 267, C1080–C1086. [Google Scholar] [CrossRef]
- Fallet, R.W.; Bast, J.P.; Fujiwara, K. Influence of Ca2+-activated K+ channels on rat renal arteriolar responses to depolarizing agonists. Am. J. Physiol. Renal. Physiol. 2001, 280, F583–F591. [Google Scholar] [CrossRef]
- Kim, E.Y.; Ridgway, L.D.; Zou, S. Alternatively spliced C-terminal domains regulate the surface expression of large conductance calcium-activated potassium (BKCa) channels. Neuroscience 2007, 146, 1652–1661. [Google Scholar] [CrossRef] [Green Version]
- Tseng-Crank, J.; Foster, C.D.; Krause, J.D. Cloning, expression, and distribution of functionally distinct Ca2+-activated K+ channel isoforms from human brain. Neuron 1994, 13, 1315–1330. [Google Scholar] [CrossRef]
- Chen, L.; Jeffries, O.; Rowe, I.C.M. Membrane Trafficking of Large Conductance Calcium-activated Potassium Channels Is Regulated by Alternative Splicing of a Transplantable, Acidic Trafficking Motif in the RCK1-RCK2 Linker *. J. Biol. Chem. 2010, 285, 23265–23275. [Google Scholar] [CrossRef] [Green Version]
- Dudem, S.; Large, R.J.; Kulkarni, S. LINGO1 is a regulatory subunit of large conductance, Ca2+-activated potassium channels. Proc. Natl. Acad. Sci. USA 2020, 117, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Filosa, J.A.; Bonev, A.D.; Straub, S.V. Local potassium signaling couples neuronal activity to vasodilation in the brain. Nat. Neurosci. 2006, 9, 1397–1403. [Google Scholar] [CrossRef]
- Zarei, M.M.; Song, M.; Wilson, R.J. Endocytic trafficking signals in KCNMB2 regulate surface expression of a voltage and Ca2+-Activated K+ channel. Neuroscience 2007, 147, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Brenner, R.; Peréz, G.J.; Bonev, A.D. Vasoregulation by the β1 subunit of the calcium-activated potassium channel. Nature 2000, 407, 870–876. [Google Scholar] [CrossRef]
- McManus, O.B.; Helms, L.M.; Pallanck, L. Functional role of the beta subunit of high conductance calcium-activated potassium channels. Neuron 1995, 14, 645–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, X.; Hite, R.K.; MacKinnon, R. Cryo-EM structure of the open high conductance Ca2+-activated K+ channel. Nature 2017, 541, 46–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hite, R.K.; Tao, X.; MacKinnon, R. Structural basis for gating the high-conductance Ca2+-activated K+ channel. Nature 2017, 541, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, C.S.; Moldenhauer, H.J.; Park, S.M. KCNMA1-linked channelopathy. J. Gen. Physiol. 2019, 151, 1173–1189. [Google Scholar] [CrossRef]
- Meredith, A.L. Genetic Methods for Studying Ion Channel Function in Physiology and Disease. In Handbook of Ion Channels; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Kim, J.-B. Channelopathies. Korean J. Pediatr. 2014, 57, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Gutterman, D.D. The coronary circulation in diabetes: Influence of reactive oxygen species on K+ channel-mediated vasodilation. Vascul. Pharmacol. 2002, 38, 43–49. [Google Scholar] [CrossRef]
- Wiecha, J.; Schläger, B.; Voisard, R. Ca(2+)-activated K+ channels in human smooth muscle cells of coronary atherosclerotic plaques and coronary media segments. Basic Res. Cardiol. 1997, 92, 233–239. [Google Scholar] [CrossRef]
- Yang, Y.; Li, P.-Y.; Cheng, J. Function of BKCa channels is reduced in human vascular smooth muscle cells from Han Chinese patients with hypertension. Hypertension 2013, 61, 519–525. [Google Scholar] [CrossRef] [Green Version]
- Kim, N.; Chung, J.; Kim, E. Changes in the Ca2+-activated K+ channels of the coronary artery during left ventricular hypertrophy. Circ. Res. 2003, 93, 541–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meredith, A.L.; Thorneloe, K.S.; Werner, M.E. Overactive Bladder and Incontinence in the Absence of the BK Large Conductance Ca2+-activated K+ Channel *. J. Biol. Chem. 2004, 279, 36746–36752. [Google Scholar] [CrossRef] [PubMed]
- Rüttiger, L.; Sausbier, M.; Zimmermann, U. Deletion of the Ca2+-activated potassium (BK) α-subunit but not the BKβ1-subunit leads to progressive hearing loss. Proc. Natl. Acad. Sci. USA 2004, 101, 12922–12927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, M.E.; Zvara, P.; Meredith, A.L. Erectile dysfunction in mice lacking the large-conductance calcium-activated potassium (BK) channel. J. Physiol. 2005, 567, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Tighilet, B.; Bourdet, A.; Péricat, D. SK Channels Modulation Accelerates Equilibrium Recovery in Unilateral Vestibular Neurectomized Rats. Pharmaceuticals 2021, 14, 1226. [Google Scholar] [CrossRef]
- Subtype-Selective Positive Modulation of KCa2.3 Channels Increases Cilia Length | ACS Chemical Biology. Available online: https://pubs.acs.org/doi/10.1021/acschembio.2c004692344–2354 (accessed on 12 January 2023).
- Wang, J.; Morishima, S.; Okada, Y. IK channels are involved in the regulatory volume decrease in human epithelial cells. Am. J. Physiol. Cell Physiol. 2003, 284, C77–C84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Womack, M.D.; Khodakhah, K. Somatic and Dendritic Small-Conductance Calcium-Activated Potassium Channels Regulate the Output of Cerebellar Purkinje Neurons. J. Neurosci. 2003, 23, 2600–2607. [Google Scholar] [CrossRef]
- Cui, M.; Qin, G.; Yu, K. Targeting the Small- and Intermediate-Conductance Ca2+-Activated Potassium Channels: The Drug-Binding Pocket at the Channel/Calmodulin Interface. NSG 2014, 22, 65–78. [Google Scholar]
- Dolga, A.M.; de Andrade, A.; Meissner, L. Subcellular expression and neuroprotective effects of SK channels in human dopaminergic neurons. Cell Death Dis. 2014, 5, e999. [Google Scholar] [CrossRef] [Green Version]
- Sankaranarayanan, A.; Raman, G.; Busch, C. Naphtho [1,2-d]thiazol-2-ylamine (SKA-31), a new activator of KCa2 and KCa3.1 potassium channels, potentiates the endothelium-derived hyperpolarizing factor response and lowers blood pressure. Mol. Pharmacol. 2009, 75, 281–295. [Google Scholar] [CrossRef]
- McNeish, A.J.; Dora, K.A.; Garland, C.J. Possible role for K+ in endothelium-derived hyperpolarizing factor-linked dilatation in rat middle cerebral artery. Stroke 2005, 36, 1526–1532. [Google Scholar] [CrossRef] [Green Version]
- Pharmacology of Small- and Intermediate-Conductance Calcium-Activated Potassium Channels. Available online: https://www.annualreviews.org/doi/epdf/10.1146/annurev-pharmtox-010919-023420 (accessed on 24 July 2022).
- Wulff, H.; Zhorov, B.S. K + Channel Modulators for the Treatment of Neurological Disorders and Autoimmune Diseases. Chem. Rev. 2008, 108, 1744–1773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crotti, L.; Odening, K.E.; Sanguinetti, M.C. Heritable arrhythmias associated with abnormal function of cardiac potassium channels. Cardiovasc. Res. 2020, 116, 1542–1556. [Google Scholar] [CrossRef] [PubMed]
- Nam, Y.-W.; Downey, M.; Rahman, M.A. Channelopathy of small- and intermediate-conductance Ca2+-activated K+ channels. Acta Pharmacol. Sin. 2022, 17, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gripp, K.W.; Smithson, S.F.; Scurr, I.J. Syndromic disorders caused by gain-of-function variants in KCNH1, KCNK4, and KCNN3—A subgroup of K+ channelopathies. Eur. J. Hum. Genet. 2021, 29, 1384–1395. [Google Scholar] [CrossRef]
- Orfali, R.; Nam, Y.-W.; Nguyen, H.M. Channelopathy-causing mutations in the S45A/S45B and HA/HB helices of KCa2.3 and KCa3.1 channels alter their apparent Ca2+ sensitivity. Cell Calcium. 2022, 102, 102538. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, M.; Ryba, L.; Křepelová, A. Zimmermann-Laband syndrome in monozygotic twins with a mild neurobehavioral phenotype lacking gingival overgrowth-A case report of a novel KCNN3 gene variant. Am. J. Med. Genet. A. 2022, 188, 1083–1087. [Google Scholar] [CrossRef]
- Bauer, C.K.; Schneeberger, P.E.; Kortüm, F. Gain-of-Function Mutations in KCNN3 Encoding the Small-Conductance Ca2+-Activated K+ Channel SK3 Cause Zimmermann-Laband Syndrome. Am. J. Hum. Genet. 2019, 104, 1139–1157. [Google Scholar] [CrossRef] [Green Version]
- Koot, B.G.P.; Alders, M.; Verheij, J. A de novo mutation in KCNN3 associated with autosomal dominant idiopathic non-cirrhotic portal hypertension. J. Hepatol. 2016, 64, 974–977. [Google Scholar] [CrossRef] [Green Version]
- A CAG repeat polymorphism of KCNN3 predicts SK3 channel function and cognitive performance in schizophrenia. EMBO Mol. Med. 2011, 3, 309–319. [CrossRef]
- Miller, M.J.; Rauer, H.; Tomita, H. Nuclear Localization and Dominant-negative Suppression by a Mutant SKCa3 N-terminal Channel Fragment Identified in a Patient with Schizophrenia*. J. Biol. Chem. 2001, 276, 27753–27756. [Google Scholar] [CrossRef] [Green Version]
- Glogowska, E.; Lezon-Geyda, K.; Maksimova, Y. Mutations in the Gardos channel (KCNN4) are associated with hereditary xerocytosis. Blood 2015, 126, 1281–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fermo, E.; Monedero-Alonso, D.; Petkova-Kirova, P. Gardos channelopathy: Functional analysis of a novel KCNN4 variant. Blood Adv. 2020, 4, 6336–6341. [Google Scholar] [CrossRef] [PubMed]
- Small-Conductance, Calcium-Activated Potassium Channels from Mammalian Brain | Science. Available online: https://www.science.org/doi/10.1126/science.273.5282.1709?keytype2=tf_ipsecsha&ijkey=85150f659083d52f2496eacd09e26e41880530f4 (accessed on 2 September 2022).
- Yan, J.; Aldrich, R.W. LRRC26 auxiliary protein allows BK channel activation at resting voltage without calcium. Nature 2010, 466, 513–516. [Google Scholar] [CrossRef] [PubMed]
- Sancho, M.; Kyle, B.D. The Large-Conductance, Calcium-Activated Potassium Channel: A Big Key Regulator of Cell Physiology. Frontiers in Physiology. 2021. Available online: https://www.frontiersin.org/articles/10.3389/fphys.2021.750615 (accessed on 27 August 2022).
- Li, Q.; Li, Y.; Wei, H. Molecular determinants of Ca2+ sensitivity at the intersubunit interface of the BK channel gating ring. Sci. Rep. 2018, 8, 509. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Sigworth, F.J. Structure of the BK potassium channel in a lipid membrane from electron cryomicroscopy. Nature 2009, 461, 292–295. [Google Scholar] [CrossRef] [Green Version]
- Nam, Y.-W.; Cui, M.; Orfali, R. Hydrophobic interactions between the HA helix and S4-S5 linker modulate apparent Ca2+ sensitivity of SK2 channels. Acta Physiol. 2021, 231, e13552. [Google Scholar] [CrossRef]
- Kuramoto, T.; Yokoe, M.; Kunisawa, N. Tremor dominant Kyoto (Trdk) rats carry a missense mutation in the gene encoding the SK2 subunit of small-conductance Ca2+-activated K+ channel. Brain Res. 2017, 1676, 38–45. [Google Scholar] [CrossRef]
- Mochel, F.; Rastetter, A.; Ceulemans, B. Variants in the SK2 channel gene (KCNN2) lead to dominant neurodevelopmental movement disorders. Brain 2020, 143, 3564–3573. [Google Scholar] [CrossRef]
- Castori, M.; Valiante, M.; Pascolini, G.; Leuzzi, V.; Pizzuti, A.; Grammatico, P. Clinical and genetic study of two patients with Zimmermann-Laband syndrome and literature review. Eur. J. Med. Genet. 2013, 56, 570–576. [Google Scholar] [CrossRef]
- Mutations in KCNH1 and ATP6V1B2 cause Zimmermann-Laband syndrome. Nature Genetics. 2015, 47, 661–667. [CrossRef] [PubMed] [Green Version]
- Kmeid, M.; Liu, H.; Ballentine, S.; Lee, H. Idiopathic Non-Cirrhotic Portal Hypertension and Porto-Sinusoidal Vascular Disease: Review of Current Data. Gastroenterol Res. 2021, 14, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Hoffmanz, J.F.; Joiner, W.; Nehrke, K. The hSK4 (KCNN4) isoform is the Ca2+-activated K+ channel (Gardos channel) in human red blood cells. Proc. Natl. Acad. Sci. USA 2003, 100, 7366–7371. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Expression and Physiology of the KCa2x, KCa3.1, and KCa1.1 channels [10].
Figure 1.
Expression and Physiology of the KCa2x, KCa3.1, and KCa1.1 channels [10].

Figure 2.
Diagram of the general KCa1.1 and KCa2.x channel structure. (A) Schematic channel topology of one KCa 1.1 α-subunit, including the pore-gate domain between (S5–S6) and a C-terminal cytosolic region that functions as a Ca2+ sensor constituted by two non-identical domains (RCK1 and RCK2), which contain high-affinity binding Ca2+ sites (Ca2+ bowl) and several domains for multiple ligands or cations such as Mg2+. (B) Schematic channel topology of one KCa2.x α-subunit, including the CaM that serves as the KCa2.x and KCa3.1 channel’s Ca2+ sensor. A and B were generated using Biorender.com.
Figure 2.
Diagram of the general KCa1.1 and KCa2.x channel structure. (A) Schematic channel topology of one KCa 1.1 α-subunit, including the pore-gate domain between (S5–S6) and a C-terminal cytosolic region that functions as a Ca2+ sensor constituted by two non-identical domains (RCK1 and RCK2), which contain high-affinity binding Ca2+ sites (Ca2+ bowl) and several domains for multiple ligands or cations such as Mg2+. (B) Schematic channel topology of one KCa2.x α-subunit, including the CaM that serves as the KCa2.x and KCa3.1 channel’s Ca2+ sensor. A and B were generated using Biorender.com.

Figure 3.
The regulation of KCa2.X Channels’ Ca2+ sensitivity.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Sensitive KCa channels reviewed.
| KCa1.1 | KCa2.x | KCa3.1 | |
|---|---|---|---|
| Expression | Neurons [7,18,19] vascular, respiratory, endocrine, retinal circulation [26], glomerular mesangial cells [27], and podocytes [29]. | Neurons, heart, and vascular endothelium [57]. | Microglia, lung epithelia, GI epithelia, T cells, and red blood cells [57]. |
| Mechanism | Activated by both Ca2+ and Voltage [18]. | Activated by low intracellular Ca2+ concentrations (Voltage-independent). | |
| Ca2+ binding sites | Two intracellular Ca2+-sensing RCK domains (RCK1 and RCK2) [16]. | Ca2+- binding (CaM) [10,70]. | |
| Ca2+ sensitivity Regulation | The inter-subunit assembly interface contains molecular determinants of Ca2+- sensitivity in KCa1.1 channels [73]. | Hydrophobic interactions between the HA helix and S4-S5 linker regulate the KCa2.x/KCa3.1 channel’s apparent Ca2+ sensitivity [75]. | |
| Channelopathies | Neurological disorders, diabetes [42], atherosclerosis [43], hypertension [44], cardiac hypertrophy [45], paroxysmal nonkinesigenic dyskinesia (PNKD), ataxia, hearing loss [47], urinary incontinence and overactive bladder [46]. | Cerebellar ataxia and tremor are associated with LOF KCa2.2 mutations [59]. The GOF mutations of KCa2.3 are linked with (ZLS) [60,62,63,64] and (INCPH) [60,62,65], and schizophrenia [66]. | The GOF mutations of KCa3.1 are linked with a subset of (HX) [68]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Orfali, R.; Albanyan, N. Ca2+-Sensitive Potassium Channels. Molecules 2023, 28, 885. https://doi.org/10.3390/molecules28020885
AMA Style
Orfali R, Albanyan N. Ca2+-Sensitive Potassium Channels. Molecules. 2023; 28(2):885. https://doi.org/10.3390/molecules28020885
Chicago/Turabian StyleOrfali, Razan, and Nora Albanyan. 2023. "Ca2+-Sensitive Potassium Channels" Molecules 28, no. 2: 885. https://doi.org/10.3390/molecules28020885