
Characterization and Comparison of Steroidal Glycosides from Polygonatum Species by High-Performance Liquid Chromatography–Electrospray Ionization Mass Spectrometry


Abstract
:1. Introduction
2. Results
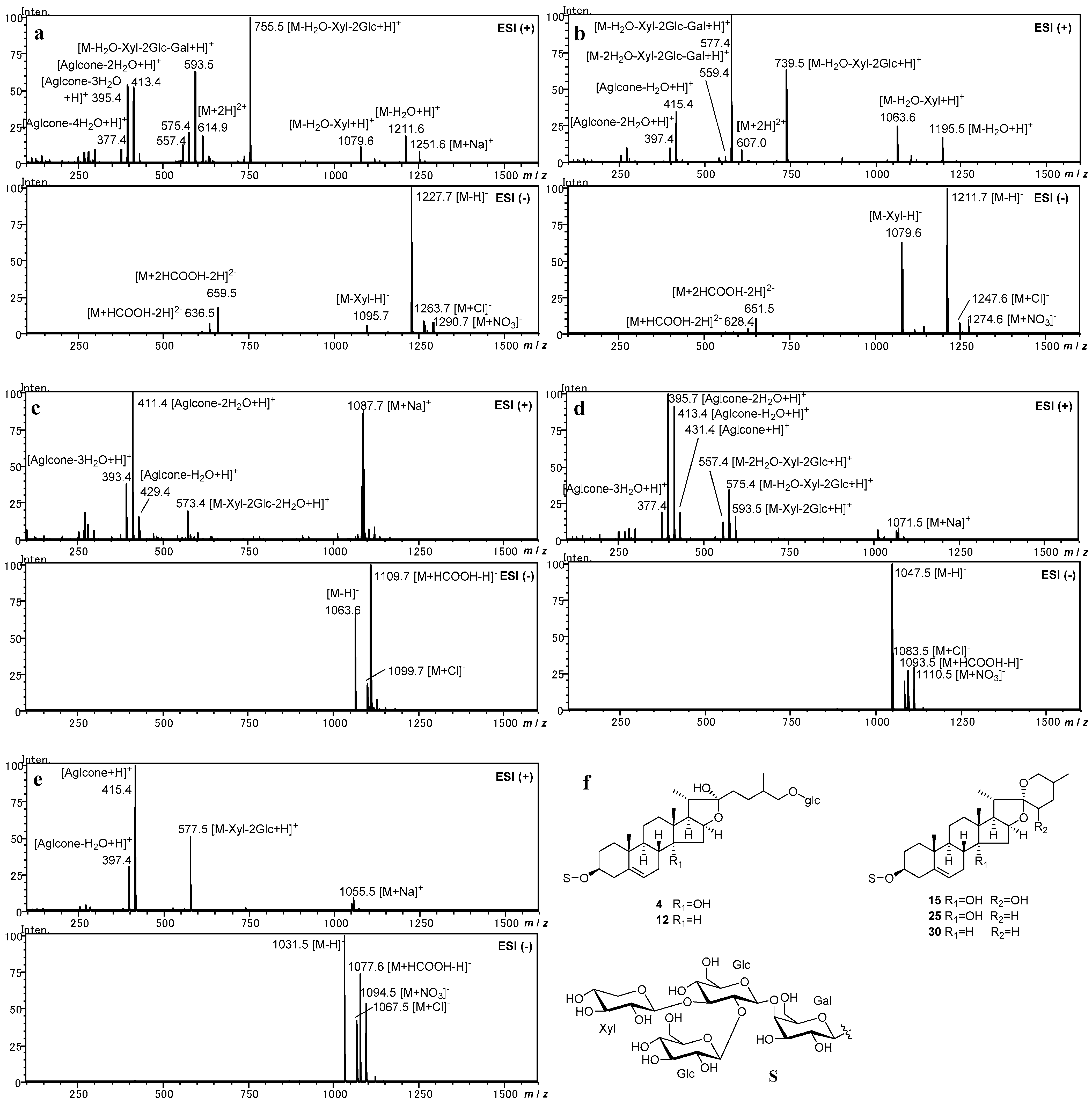
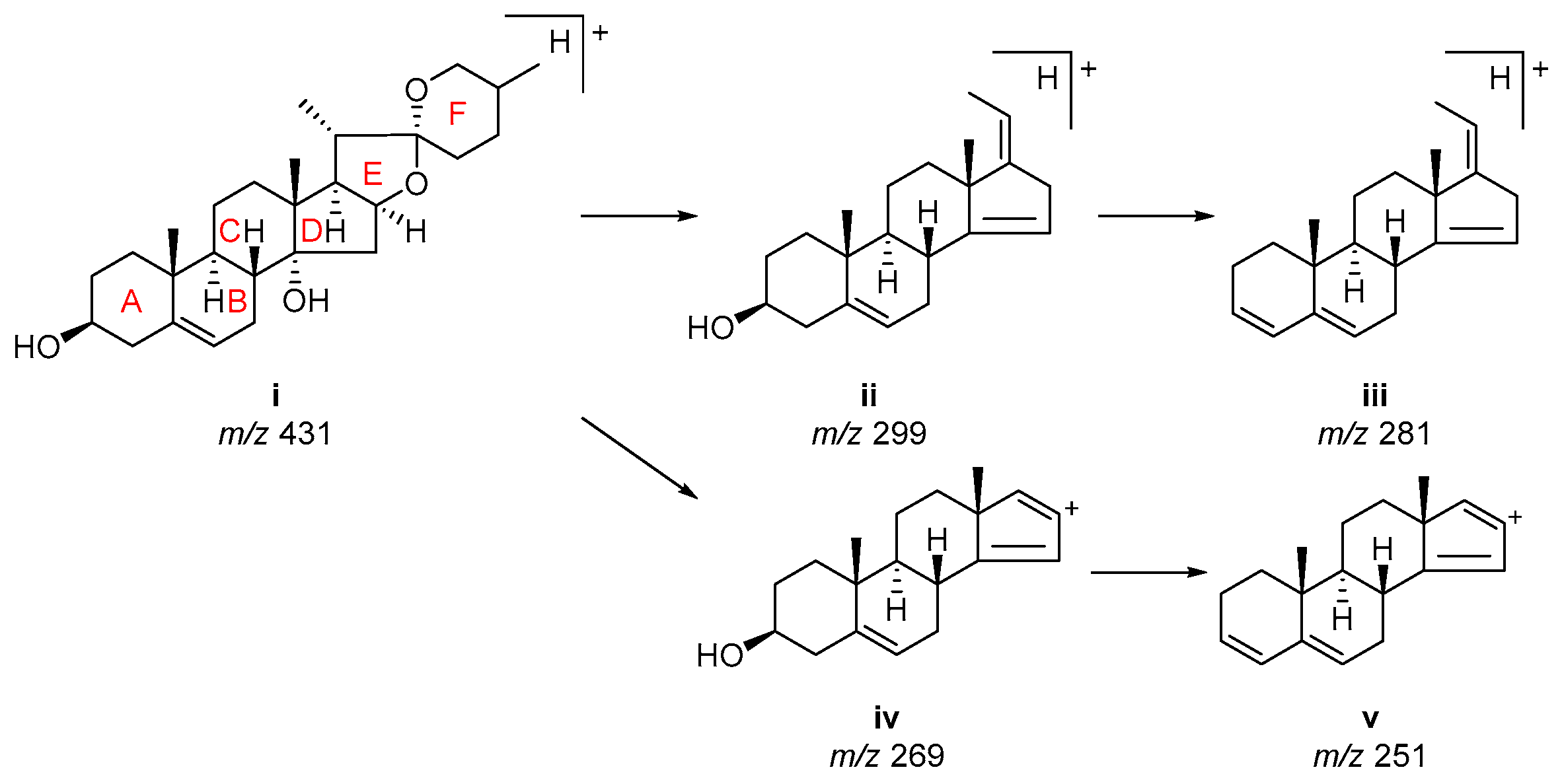
2.1. Investigation of ESI-MS Fragmentation of Authentic Steroidal Glycosides
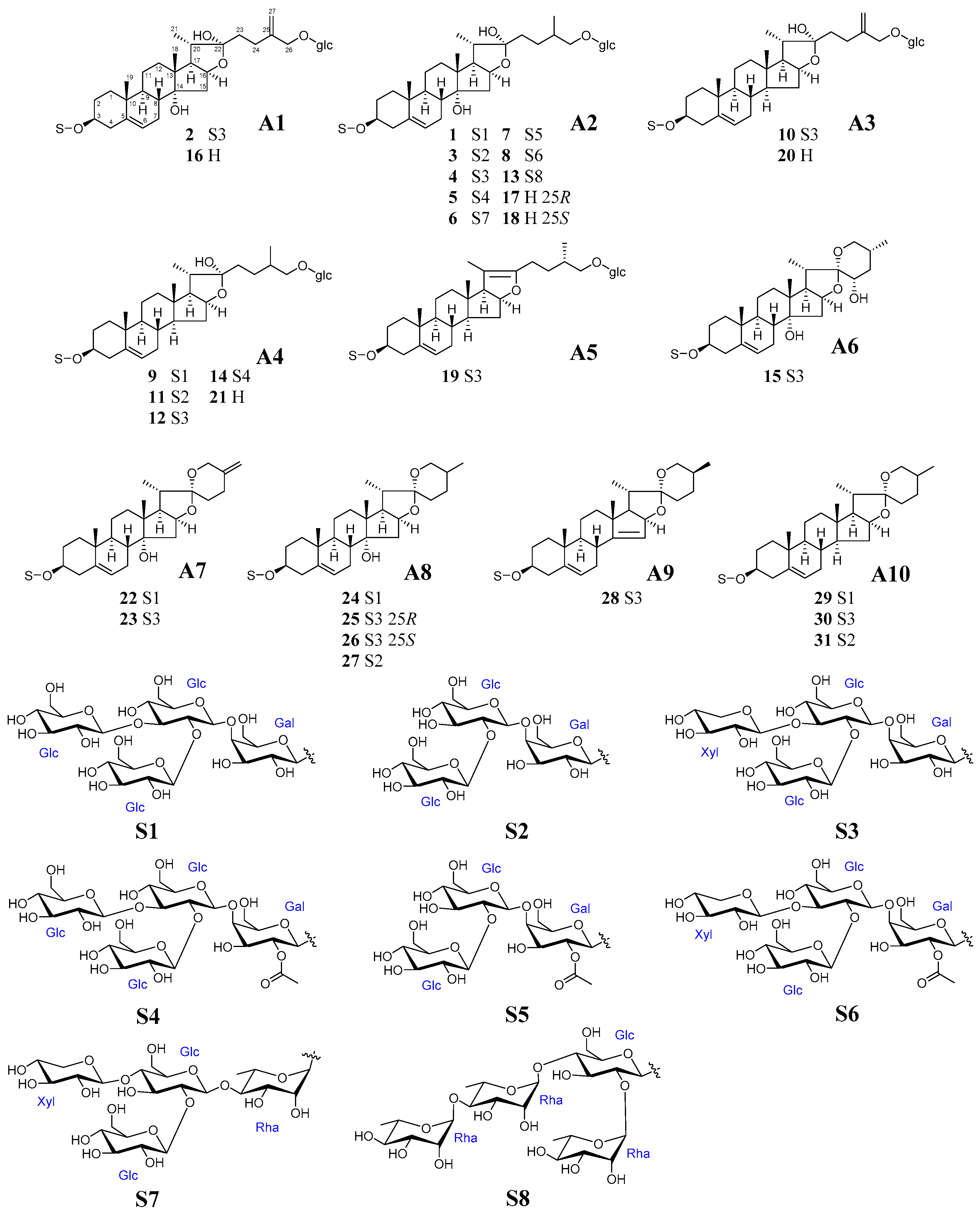
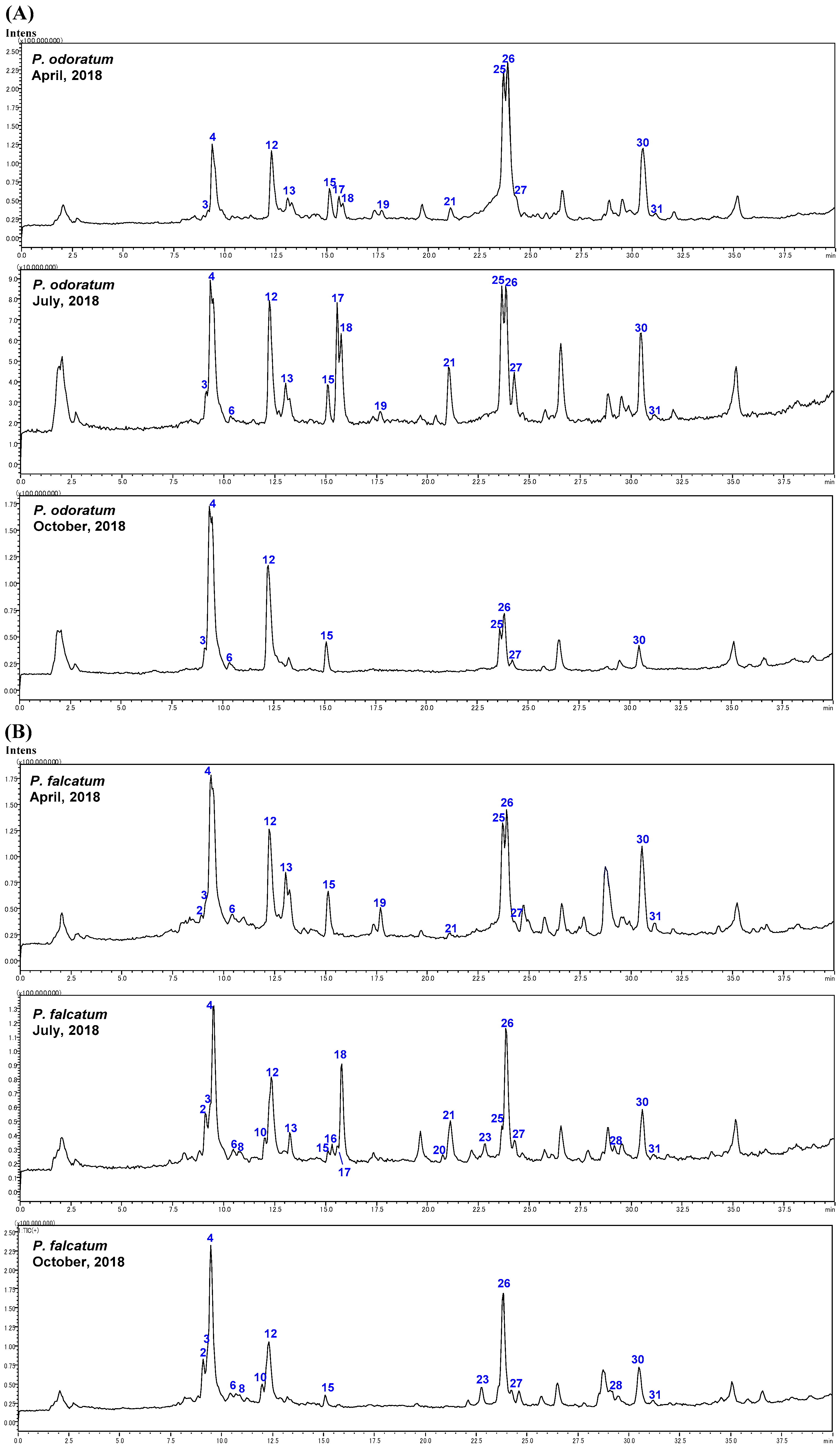
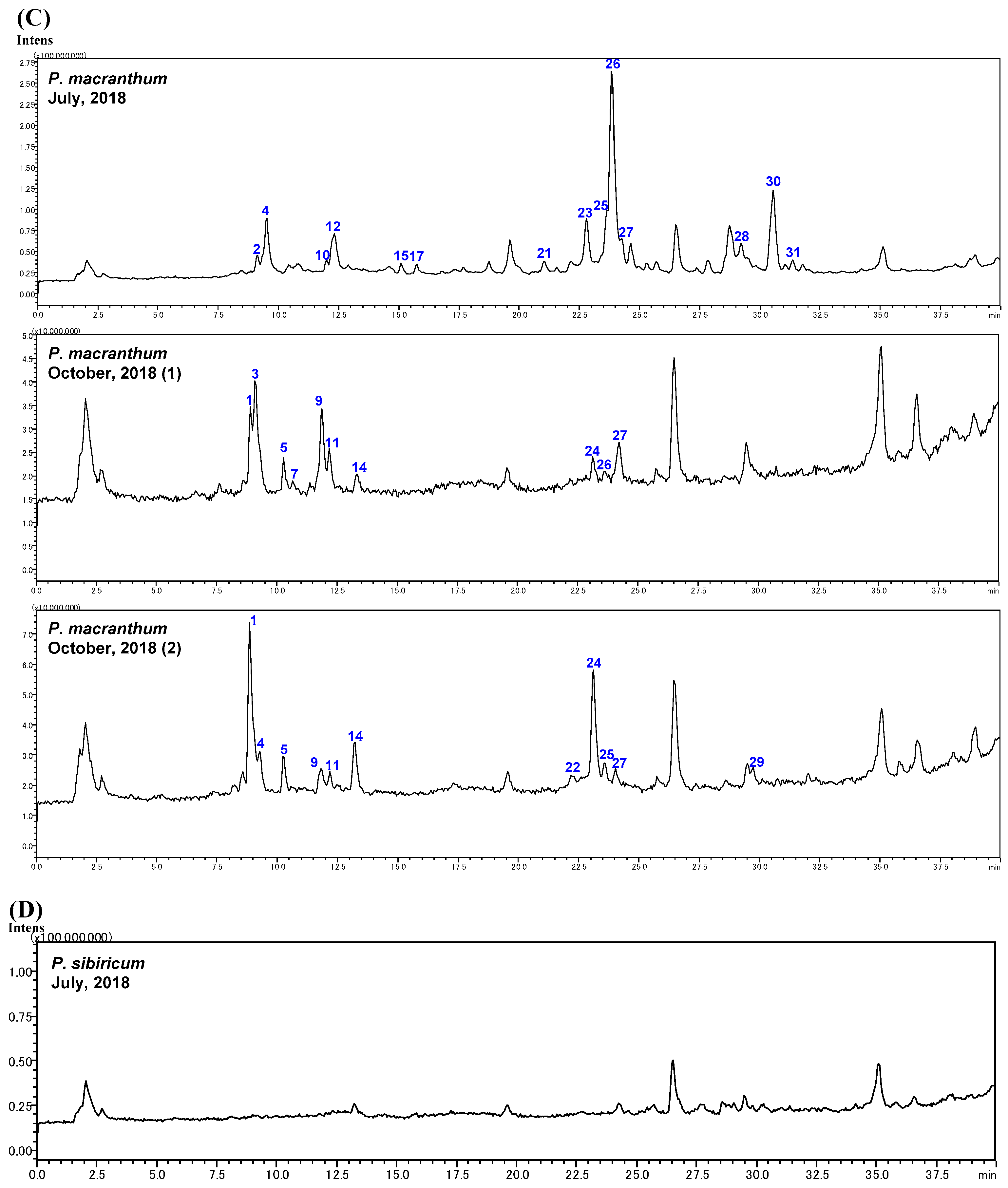
2.2. Identification of Steroidal Glycosides in the Sample Solutions
2.3. The Biogenetic Pathway of Steroidal Glycosides in Polygonatum Species
3. Materials and Methods
3.1. General Methods
3.2. Plant Materials
3.3. Preparation of the Sample Solutions
3.4. LC-MS Analysis
3.5. Isolation of Sibiricoside B (15)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ohashi, H.; Kadota, Y.; Murata, J.; Yonekura, K.; Kihara, H. Wild Flowers of Japan, 2nd ed.; Heibonsha: Tokyo, Japan, 2015; Volume 1, p. 256. [Google Scholar]
- Hatfield, G. Encyclopedia of Folk Medicine: Old World and New World Traditions; ABC-CLIO: Santa Barbara, CA, USA, 2004; p. 150. [Google Scholar]
- Dhyani, A.; Nautiyal, B.P.; Nautiyal, M.C. Importance of Astavarga plants in traditional systems of medicine in Garhwal, Indian Himalaya. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2010, 6, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Shanghai Scientific and Technical Publishers. Dictionary of Chinese Materia Medica Japanese Edition; Shogakukan: Tokyo, Japan, 1985; Volume 1, pp. 144–147. [Google Scholar]
- Pharmacopoeia Committee of P. R. China. Pharmacopoeia of People’s Republic of China; China Medical Science and Technology Press: Beijing, China, 2020; pp. 86, 319. [Google Scholar]
- The Committee on the Japanese Pharmacopoeia. The Japanese Pharmacopoeia, 18th ed.; Ministry of Health, Labour and Welfare: Tokyo, Japan, 2021; pp. 2103–2104.
- Satake, Y. Wild Flowers of Japan: Herbaceous Plants 1; Heibonsha: Tokyo, Japan, 1982; p. 45. [Google Scholar]
- Zhao, P.; Zhao, C.; Li, X.; Gao, Q.; Huang, L.; Xiao, P.; Gao, W. The genus Polygonatum: A review of ethnopharmacology, phytochemistry and pharmacology. J. Ethnopharmacol. 2018, 214, 274–291. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Li, W.; Nagata, K.; Fu, H.; Okada, S.; Tanabe, I.; Kobori, Y.; Higai, K.; Norie, W.; Sasaki, T. Isolation, structural elucidation, and liquid chromatography-mass spectrometry analysis of steroidal glycosides from Polygonatum odoratum. J. Agric. Food Chem. 2018, 66, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Ahn, M.J.; Kim, J. Identification and quantification of steroidal saponins in Polygonatum species by HPLC/ESI/MS. Arch. Pharm. Res. 2005, 28, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Takagi, H. Breaking of two types of dormancy in seeds of Polygonatum odoratum used as vegetables. J. Jpn. Soc. Hortic. Sci. 2001, 70, 416–423. [Google Scholar] [CrossRef]
- Son, K.H.; Do, J.C. Steroidal saponins from the rhizomes of Polygonatum sibiricum. J. Nat. Prod. 1990, 53, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Shin, S.Y.; Moon, Y.J.; Seo, J.Y.; Kim, H.K.; Whang, W.K. Content analysis and classification for Polygonati Odorati Rhizoma and Polygonati Rhizoma by steroidal saponin. Yakhak Hoechi 2010, 54, 441–448. [Google Scholar]
- Ahn, M.J.; Kim, C.Y.; Yoon, K.D.; Ryu, M.Y.; Cheong, J.H.; Chin, Y.W.; Kim, J. Steroidal Saponins from the Rhizomes of Polygonatum sibiricum. J. Nat. Prod. 2006, 69, 360–364. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, L.; Kou, J.P.; Zhu, D.N.; Qi, J.; Yu, B.Y. Steroidal sapogenins and glycosides from the fibrous roots of Polygonatum odoratum with inhibitory effect on tissue factor (TF) procoagulant activity. Steroids 2014, 89, 1–10. [Google Scholar] [CrossRef]
- Yu, H.S.; Ma, B.P.; Song, X.B.; Kang, L.P.; Zhang, T.; Fu, J.; Zhao, Y.; Xiong, C.Q.; Tan, D.W.; Zhang, L.J.; et al. Two New Steroidal Saponins from the Processed Polygonatum kingianum. Helv. Chim. Acta 2010, 93, 1086–1092. [Google Scholar] [CrossRef]
- Jin, J.M.; Zhang, Y.J.; Li, H.Z.; Yang, C.R. Cytotoxic Steroidal Saponins from Polygonatum zanlanscianense. J. Nat. Prod. 2004, 67, 1992–1995. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.X.; Zhang, Y.J.; Li, H.Z.; Yang, C.R. Polyhydroxylated steroidal constituents from the fresh rhizomes of Tupistra yunnanensis. Steroids 2005, 70, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Mari, A.; Napolitano, A.; Perrone, A.; Pizza, C.; Piacente, S. An analytical approach to profile steroidal saponins in food supplements: The case of Ruscus aculeatus. Food Chem. 2012, 134, 461–468. [Google Scholar] [CrossRef]
- Kintya, P.K.; Mashchenko, N.E.; Kononova, N.I.; Lazurevskii, G.V. Steroid saponins. XII. Preliminary characterization of steroid glycosides from Funkia ovata. Chem. Nat. Compd. 1976, 12, 241–242. [Google Scholar] [CrossRef]
- Janeczko, Z.; Jansson, P.E.; Sendra, J. A new steroidal saponin from Polygonatum officinale. Plant. Med. 1987, 53, 52–54. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xiang, L.; Yi, X.; He, X. Potential Anti-inflammatory Steroidal Saponins from the Berries of Solanum nigrum L. (European Black Nightshade). J. Agric. Food Chem. 2017, 65, 4262–4272. [Google Scholar] [CrossRef]
- Qin, Z.F.; Dai, Y.; Yao, Z.H.; He, L.L.; Wang, Q.Y.; Geng, J.L.; Chen, H.F.; Yao, X.S. Study on chemical profiles and metabolites of Allii Macrostemonis Bulbus as well as its representative steroidal saponins in rats by ultra-performance liquid chromatography coupled with quadrupole time-of-flight tandem mass spectrometry. Food Chem. 2016, 192, 499–515. [Google Scholar] [CrossRef]
- Bai, H.; Li, W.; Zhao, H.; Anzai, Y.; Li, H.; Guo, H.; Kato, F.; Koike, K. Isolation and structural elucidation of novel cholestane glycosides and spirostane saponins from Polygonatum odoratum. Steroids 2014, 80, 7–14. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, E.; Li, P. Chemotaxonomic studies of nine Paris species from China based on ultra-high performance liquid chromatography tandem mass spectrometry and Fourier transform infrared spectroscopy. J. Pharm. Biomed. Anal. 2017, 140, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Li, D.; Zhu, W.; Zhang, J.; Peng, P. Steroidal saponins from the rhizomes of Polygonatum odoratum. Nat. Prod. Res. 2009, 23, 940–947. [Google Scholar] [CrossRef]
- Quan, L.T.; Wang, S.C.; Zhang, J. Chemical constituents from Polygonatum odoratum. Biochem. Syst. Ecol. 2015, 58, 281–284. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peaks | Structure | Rt (min) | Formula | MW | Positive Ion (m/z) | Negative Ion (m/z) | Occurrence | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Aglycone | Sugar | [M+Na]+ | [M-H2O+H]+ | [Aglycone-H2O+H]+ | [Aglycone- 2H2O+H]+ | [Aglycone- 3H2O+H]+ | Others | [M-H]− | [M+Cl]− | [M+HCOOH-H]− | ||||||||
| 1 | A2 | S1 | 8.90 | C57H94O30 | 1259.4 | 1281.6 | 1241.6 | 413.4 | 395.4 | 755.5 | 575.4 | 1257.7 | 1293.7 | PM | ||||
| 2 | A1 | S3 | 9.11 | C56H90O29 | 1227.3 | 1249.6 | 1209.6 | 411.4 | 393.4 | 753.5 | 591.4 | 1225.7 | 1261.7 | PF, PM | ||||
| 3 b | A2 | S2 | 9.18 | C51H84O25 | 1097.2 | 1119.6 | 1079.6 | 413.5 | 395.3 | 917.6 | 755.5 | 593.5 | 1095.5 | PO, PF, PM | ||||
| 4 a,b | A2 | S3 | 9.38 | C56H92O29 | 1229.3 | 1251.6 | 1211.6 | 413.3 | 395.4 | 755.5 | 593.5 | 575.4 | 557.4 | 1227.7 | 1263.7 | PO, PF, PM | ||
| 5 | A2 | S4 | 10.28 | C59H96O31 | 1301.4 | 1323.6 | 1283.6 | 413.4 | 395.4 | 797.5 | 593.5 | 1299.7 | 1335.7 | PM | ||||
| 6 | A2 | S7 | 10.31 | C56H92O28 | 1213.3 | 1235.6 | 1195.6 | 413.4 | 395.5 | 739.5 | 593.5 | 1211.7 | 1247.5 | PO, PF | ||||
| 7 | A2 | S5 | 10.66 | C53H86O26 | 1139.2 | 1161.6 | 1121.6 | 413.7 | 395.6 | 959.5 | 797.7 | 593.5 | 1137.7 | 1173.8 | PM | |||
| 8 | A2 | S6 | 10.78 | C58H94O30 | 1271.4 | 1293.6 | 1253.5 | 413.4 | 395.4 | 797.6 | 755.4 | 593.6 | 1269.8 | 1305.7 | PF | |||
| 9 b | A4 | S1 | 11.87 | C57H94O29 | 1243.4 | 1265.6 | 1225.6 | 415.4 | 397.4 | 901.6 | 739.5 | 577.4 | 1241.7 | 1277.7 | PM | |||
| 10 | A3 | S3 | 12.01 | C56H90O28 | 1211.3 | 1233.6 | 1193.6 | 413.3 | 395.3 | 1031.5 | 899.4 | 737.6 | 575.4 | 1209.7 | 1245.5 | PF, PM | ||
| 11 | A4 | S2 | 12.18 | C51H84O24 | 1081.2 | 1103.6 | 1063.6 | 415.5 | 397.5 | 901.6 | 739.6 | 577.4 | 1079.6 | 1115.6 | PM | |||
| 12 a,b | A4 | S3 | 12.28 | C56H92O28 | 1213.3 | 1235.6 | 1195.7 | 415.4 | 397.4 | 1033.6 | 901.6 | 739.6 | 577.5 | 1211.7 | 1247.7 | PO, PF, PM | ||
| 13 | A2 | S8 | 13.08 | C57H94O27 | 1211.4 | 1233.6 | 413.4 | 395.4 | 901.6 | 755.4 | 575.5 | 1209.7 | 1245.6 | 1255.7 | PO, PF | |||
| 14 | A4 | S4 | 13.32 | C59H96O30 | 1285.4 | 1307.6 | 1267.6 | 415.5 | 397.4 | 943.7 | 781.5 | 577.4 | 1283.7 | 1319.7 | PM | |||
| 15 a,b | A6 | S3 | 15.14 | C50H80O24 | 1065.2 | 1087.6 | 411.4 | 393.4 | 573.4 | 1063.6 | 1109.6 | PO, PF, PM | ||||||
| 16 | A1 | H | 15.32 | C33H52O10 | 608.8 | 631.4 | 591.5 | 607.4 | 653.4 | PF | ||||||||
| 17 b | A2 | H | 15.59 | C33H54O10 | 610.8 | 633.5 | 593.5 | 431.4 | 609.4 | 655.5 | PO, PF, PM | |||||||
| 18 b | A2 | H | 15.78 | C33H54O10 | 610.8 | 633.4 | 593.5 | 431.2 | 609.4 | 655.5 | PO, PF, PM | |||||||
| 19 b | A5 | S3 | 17.70 | C56H90O27 | 1195.3 | 1217.7 | 397.4 | 739.5 | 577.5 | 415.4 | 1193.7 | 1229.8 | 1239.8 | PO, PF | ||||
| 20 | A3 | H | 20.74 | C33H52O9 | 592.8 | 615.4 | 575.4 | 413.5 | 637.5 | PF | ||||||||
| 21 b | A4 | H | 21.08 | C33H54O9 | 594.8 | 617.5 | 577.5 | 415.3 | 639.5 | PO, PF, PM | ||||||||
| 22 b | A7 | S1 | 22.29 | C51H80O24 | 1077.2 | 1099.6 | 411.4 | 393.4 | 573.6 | 1075.6 | 1121.6 | PM | ||||||
| 23 b | A7 | S3 | 22.81 | C50H78O23 | 1047.2 | 1069.6 | 411.4 | 393.3 | 573.4 | 1045.6 | 1091.6 | PF, PM | ||||||
| 24 b | A8 | S1 | 23.12 | C51H82O24 | 1079.2 | 1101.6 | 413.4 | 395.4 | 575.4 | 431.4 | 1077.7 | 1123.6 | PM | |||||
| 25 a,b | A8 | S3 | 23.69 | C50H80O23 | 1049.2 | 1071.6 | 413.4 | 395.7 | 575.2 | 557.2 | 431.4 | 1047.6 | 1093.6 | PO, PF, PM | ||||
| 26 b | A8 | S3 | 23.89 | C50H80O23 | 1049.2 | 1071.5 | 413.4 | 395.4 | 575.4 | 557.4 | 431.4 | 1047.6 | 1093.6 | PO, PF, PM | ||||
| 27 b | A8 | S2 | 24.26 | C45H72O19 | 917.1 | 939.5 | 413.4 | 395.4 | 575.4 | 431.4 | 915.5 | 961.6 | PO, PF, PM | |||||
| 28 b | A9 | S3 | 29.21 | C50H78O22 | 1031.2 | 1053.6 | 395.4 | 575.4 | 413.4 | 1029.6 | 1075.6 | PF | ||||||
| 29 b | A10 | S1 | 29.71 | C51H82O23 | 1063.2 | 1085.6 | 397.3 | 577.4 | 415.3 | 1061.5 | 1107.7 | PM | ||||||
| 30 a,b | A10 | S3 | 30.52 | C50H80O22 | 1033.2 | 1055.6 | 397.4 | 577.5 | 415.4 | 1031.6 | 1077.6 | PO, PF, PM | ||||||
| 31 b | A10 | S2 | 31.16 | C45H72O18 | 901.1 | 923.6 | 397.4 | 577.7 | 415.4 | 899.6 | 945.6 | PO, PF, PM | ||||||
| No. | Species | Sample | Medicinal Parts |
|---|---|---|---|
| 1 | P. odoratum | PO4 | Rhizomes |
| 2 | P. odoratum | PO7 | Rhizomes |
| 3 | P. odoratum | PO10 | Rhizomes |
| 4 | P. falcatum | PF4 | Rhizomes |
| 5 | P. falcatum | PF7 | Rhizomes |
| 6 | P. falcatum | PF10 | Rhizomes |
| 7 | P. macranthum | PM7 | Rhizomes |
| 8 | P. macranthum | PM10 | Rhizomes |
| 9 | P. macranthum | PM10-2 | Rhizomes |
| 10 | P. sibiricum | OS7 | Rhizomes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, D.; Kikuchi, T.; Li, W. Characterization and Comparison of Steroidal Glycosides from Polygonatum Species by High-Performance Liquid Chromatography–Electrospray Ionization Mass Spectrometry. Molecules 2023, 28, 705. https://doi.org/10.3390/molecules28020705
Liu D, Kikuchi T, Li W. Characterization and Comparison of Steroidal Glycosides from Polygonatum Species by High-Performance Liquid Chromatography–Electrospray Ionization Mass Spectrometry. Molecules. 2023; 28(2):705. https://doi.org/10.3390/molecules28020705
Chicago/Turabian StyleLiu, Danyang, Takashi Kikuchi, and Wei Li. 2023. "Characterization and Comparison of Steroidal Glycosides from Polygonatum Species by High-Performance Liquid Chromatography–Electrospray Ionization Mass Spectrometry" Molecules 28, no. 2: 705. https://doi.org/10.3390/molecules28020705