
Identification of Secondary Metabolites of Interest in Pleurotus djamor Using Agave tequilana Bagasse
 ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
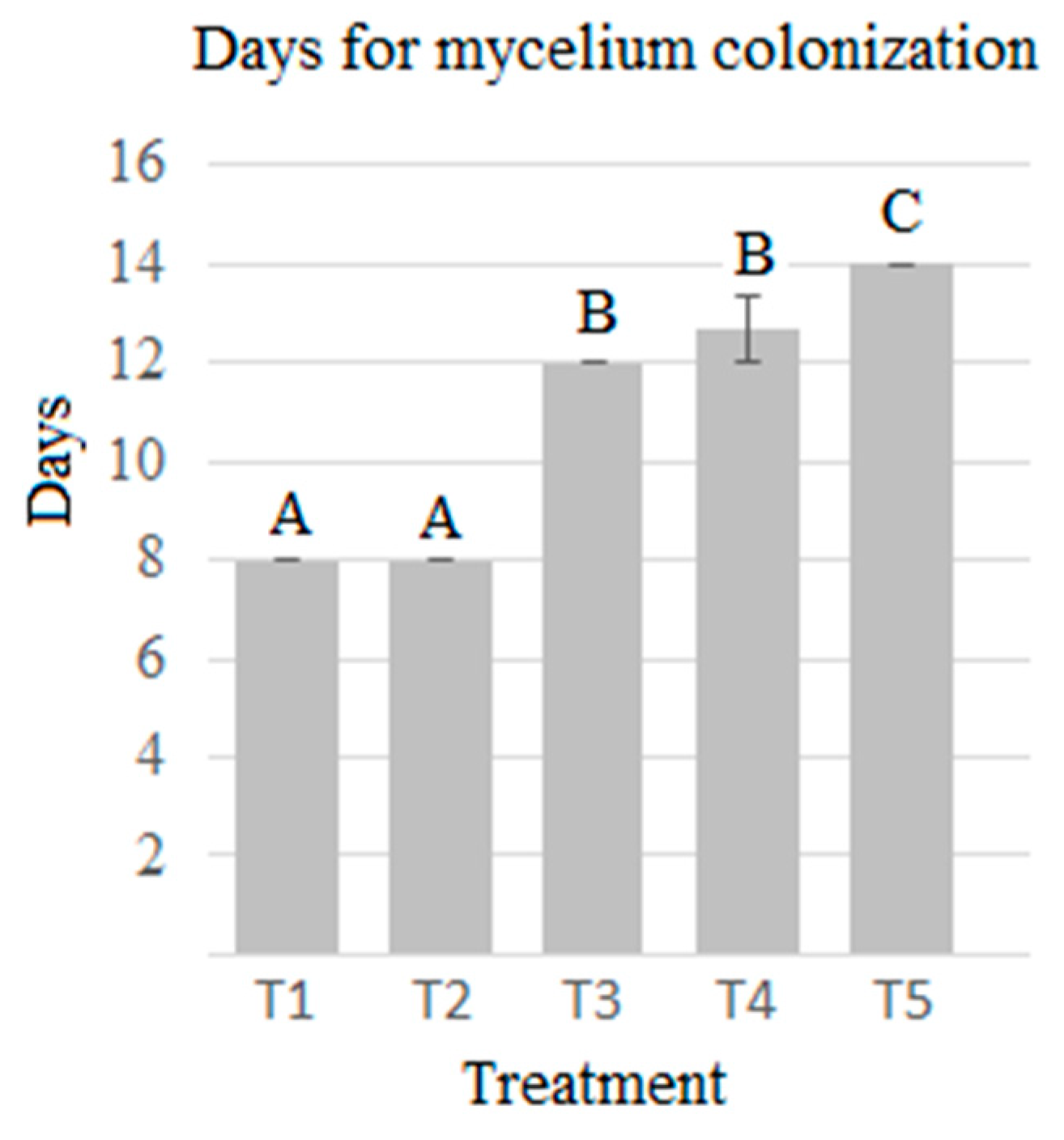
2.1. Performance and Biological Efficiency
2.2. Bromatological Analysis of Fruiting Bodies (Basidiomycetes)
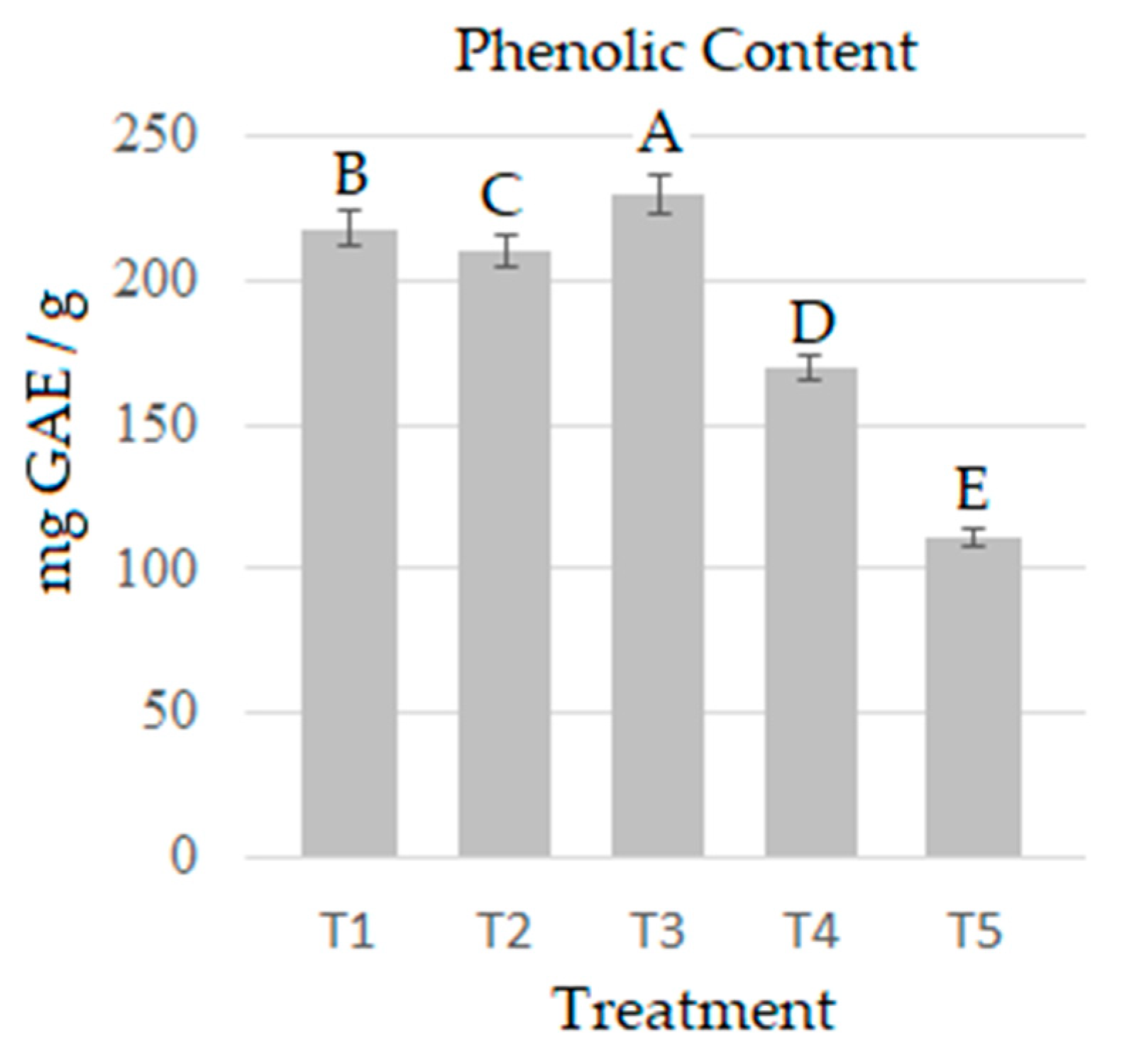
2.3. Quantification of Phenols
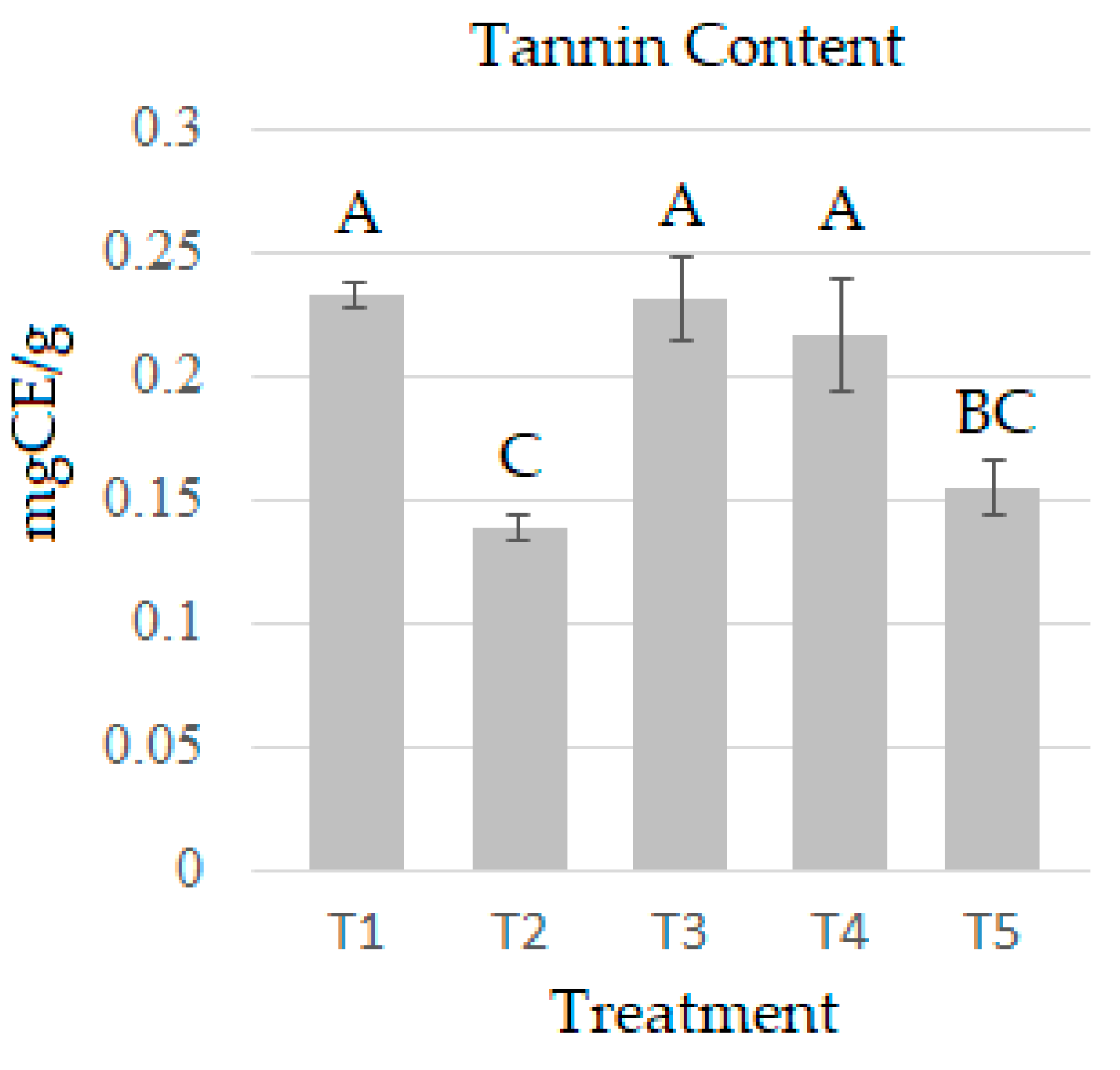
2.4. Condensed Tannins
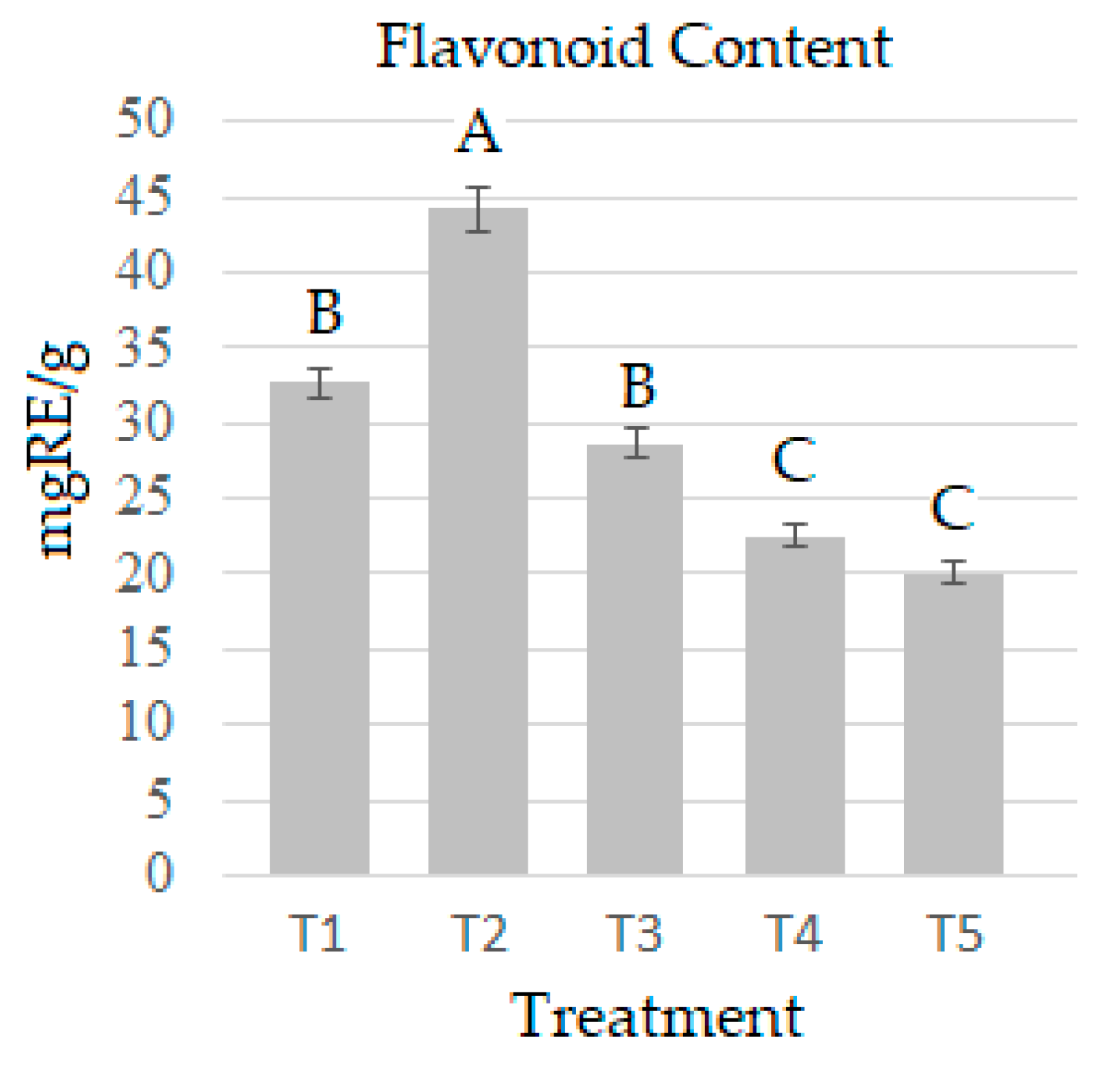
2.5. Quantification of Flavonoids
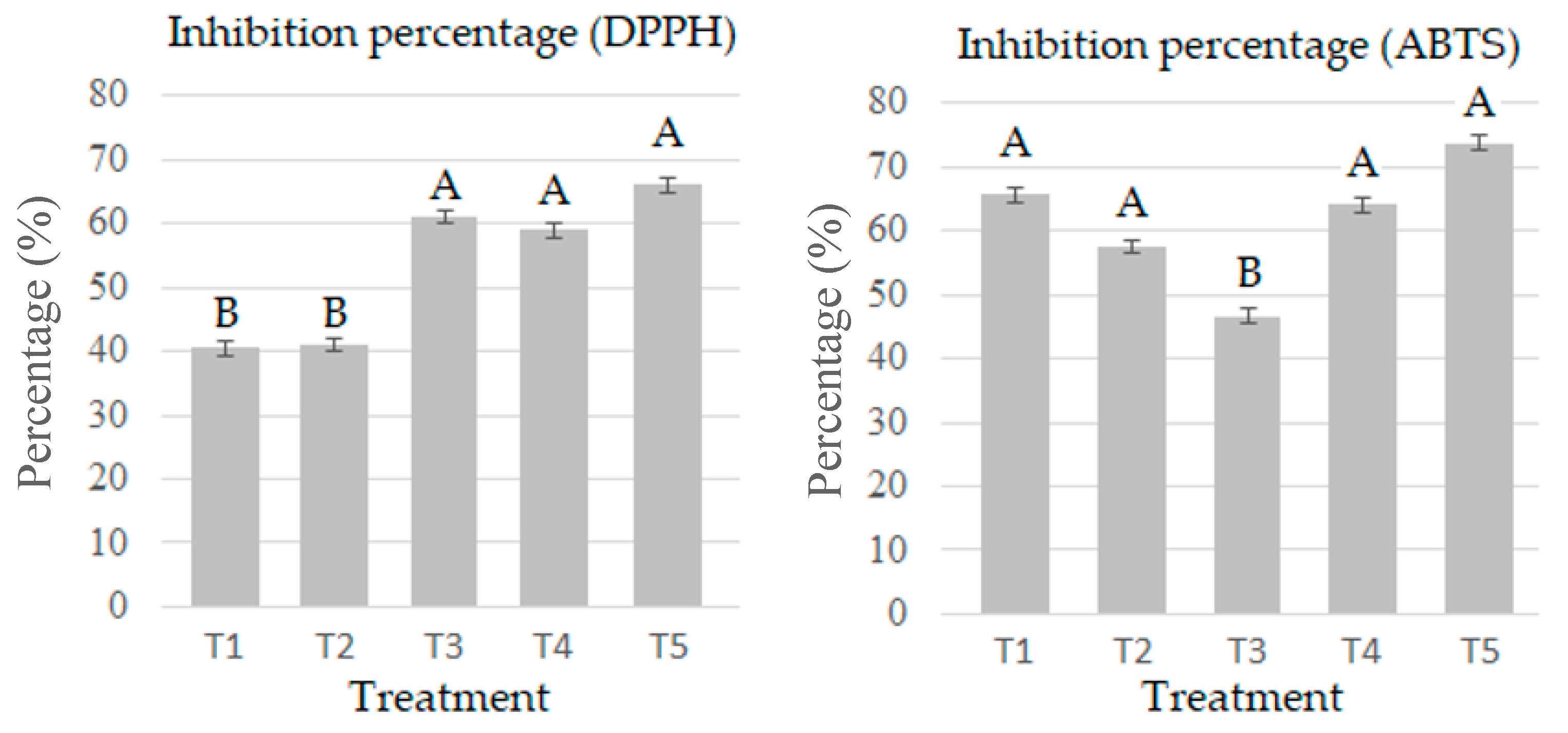
2.6. Determination of the Antioxidant Capacity by the DPPH Method
2.7. Determination of the Antioxidant Capacity by the ABTS+ Method
2.8. Gas-Chromatography-Mass Spectrometry (GC-MS) Analysis
3. Materials and Methods
3.1. Location and Cultivation System
3.2. Sampling and Analysis Methods
3.3. Bromatological Test of Fruiting Bodies
3.4. Methanolic Extraction for Polyphenolic Compounds
3.5. Quantification of Total Phenols
3.6. Condensed Tannins
3.7. Total Flavonoids
3.8. Antioxidant Activity
3.8.1. DPPH
3.8.2. ABTS+
3.8.3. Gas-Chromatography-Mass Spectrometry (GC-MS) Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beltrán-Delgado, Y.; Morris-Quevedo, H.; Llauradó-Maury, G.; Bermúdez-Savón, R.C.; García-Oduardo, N. Procedimientos producción de pleurotus con potencial. Rev. Cubana Quím. 2020, 32, 245–261. [Google Scholar]
- Chang, S.T.; Miles, P.G. Mushrooms: Cultivation, Nutritional Value, Medicinal Effect, and Environmental Impact, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar] [CrossRef]
- Martínez-Carrera, D.; Sobal, M.; Morales, P.; Martínez, W.; Martínez, M.; Mayett, Y. Los Hongos Comestibles: Propiedades Nutricionales, Medicinales, y su Contribución a la Alimentación Mexicana El Shiitake; Colegio de Postgraduados: Texcoco, Mexico, 2004; Available online: http://micaplint.fws1.com (accessed on 14 April 2022).
- Rao, A.V.; Rao, L.G. Carotenoids and human health. Pharmacol. Res. 2007, 55, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Robaszkiewicz, A.; Bartosz, G.; Ławrynowicz, M.; Soszyński, M. The Role of Polyphenols, β-Carotene, and Lycopene in the Antioxidative Action of the Extracts of Dried, Edible Mushrooms. J. Nutr. Metab. 2010, 2010, 173274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonatti, M.; Karnopp, P.; Soares, H.M.; Furlan, S.A. Evaluation of Pleurotus ostreatus and Pleurotus sajor-caju nutritional characteristics when cultivated in different lignocellulosic wastes. Food Chem. 2004, 3, 425–428. [Google Scholar] [CrossRef]
- Royse, D.J.; Baars, J.; Tan, Q. Current Overview of Mushroom Production in the World. In Edible and Medicinal Mushrooms; Wiley: Hoboken, NJ, USA, 2017; pp. 5–13. [Google Scholar] [CrossRef]
- Atila, F. Evaluation of Suitability of Various Agro-Wastes for Productivity of Pleurotus djamor, Pleurotus citrinopileatus and Pleurotus eryngii Mushrooms. J. Exp. Agric. Int. 2017, 17, 1–11. [Google Scholar] [CrossRef]
- Funaki, M.; Ducca, F.; Ferdinandi, D.M.; Zonetti, P.D.C.; Rosado, F.R. Análisis de substratos alternativos para o cultivo de Pleurotus ostreatoroseus e Pleurotus florida. Rev. Agronegocio Ambiente 2010, 3, 249–261. [Google Scholar]
- Hasan, M.T.; Khatun, M.H.A.; Sajib, M.A.M.; Rahman, M.M.; Rahman, M.S.; Roy, M.; Miah, M.N.; Ahmed, K.U. Effect of Wheat Bran Supplement with Sugarcane Bagasse on Growth, Yield and Proximate Composition of Pink Oyster Mushroom (Pleurotus djamor). Am. J. Food Sci. Technol. 2015, 3, 150–157. Available online: https://www.researchgate.net/publication/293817288_Effect_of_Wheat_Bran_Supplement_with_Sugarcane_Bagasse_on_Growth_Yield_and_Proximate_Composition_of_Pink_Oyster_Mushroom_Pleurotus_djamor (accessed on 3 October 2022).
- Salmones, D. Pleurotus djamor, un hongo con potencial aplicación biotecnológica para el neotrópico Pleurotus djamor, a mushroom with potential biotechological application for the Neotropic. Rev. Mex. Mic. 2017, 46, 73–85. Available online: https://www.scielo.org.mx/pdf/rmm/v46/0187-3180-rmm-46-73.pdf (accessed on 23 October 2021).
- Vega, A.; Franco, H. Productividad y calidad de los cuerpos fructíferos de los hongos comestibles Pleurotus pulmonarius RN2 y P. djamor RN81 y RN82 cultivados sobre sustratos lignocelulósicos. Inf. Tecnol. 2013, 24, 69–78. [Google Scholar] [CrossRef]
- Andrew, S.M.; Maliondo, S.M.S.; Munishi, P.K.T.; Msita, H. Yield of Edible Pleurotus Mushrooms Grown on Rice Straw with and without Chicken Manure Supplementation in Morogoro, Tanzania. Tanzan. J. For. Nat. Conserv. 2008, 77, 46–53. [Google Scholar]
- Vega, A.; de León, J.A.; Miranda, S.; Reyes, S.M. Agro-industrial waste improves the nutritional and antioxidant profile of Pleurotus djamor. Clean. Waste Syst. 2022, 2, 100018. [Google Scholar] [CrossRef]
- Mata, G.; Salmones, D. World Society for Mushroom Biology and Mushroom Products Symposium: Prospects of Mushroom Cultivation in Latin America for the XXI Century Edible mushroom cultivation at the institute of ecology in Mexico. Micol. Apl. Int. 2003, 15, 23–29. Available online: https://www.redalyc.org/pdf/685/68515104.pdf (accessed on 7 April 2022).
- Paswal, S.; Mughal, N.; Bharti, V.; Mahajan, S.; Majeed, M. Evaluation of Locally Available Substrates for Sporocorps Production of Pink Pleurotus [Pleurotus djamor (Rumph.ex.Fr.) boedijn] Mushroom. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1677–1684. [Google Scholar] [CrossRef]
- Ancona, L.; Zapata, G.C.; Casso, R.B.; Castro, C.S. Cultivo de Pleurotus Djamor y P. Ostreatus en Yucatán.x. In El cultivo de Pleurotus spp en México; Vázquez, S., Martínez, D., Eds.; Colegio de la Frontera Sur: Campeche, Mexico, 2007. [Google Scholar]
- Cobá, E.H.L.; Méndez, A.; Peralta, M. Cultivo de Pleurotus djamor en condiciones de laboratorio y en una casa rural tropical. Sci. Fungorum 2005, 21, 93–97. Available online: http://www.redalyc.org/articulo.oa?id=88302115 (accessed on 20 October 2021).
- Mshandete, A.M.; Cuff, J. Cultivation of three types of indigenous wild edible mushrooms: Coprinus cinereus, Pleurotus flabellatus and Volvariella volvocea on composted sisal decortications residue in Tanzania. Afr. J. Biotechnol. 2008, 7, 4551–4562. [Google Scholar] [CrossRef]
- Salmones, D.; Valdéz, L.M.; Gaitán-Hernández, R. Entrecruzamiento y evaluación de la producción de las variedades de Pleurotus djamor (Fr.) Boedijn. Sci. Fungorum 2004, 63, 21–26. [Google Scholar]
- González, M.R.C.; Eguiarte, D.R.G.; Macías, R.R.; Corral, J.A.R.; Durán Puga, N. Caracterización química y física del bagazo de agave tequilero compostado con biosólidos de vinaza como componente de sustratos para cultivos en contenedor. Rev. Int. Contam. Ambient. 2018, 34, 373–382. [Google Scholar] [CrossRef]
- Hoa, H.T.; Wang, C.L.; Wang, C.H. The effects of different substrates on the growth, yield, and nutritional composition of two oyster mushrooms (Pleurotus ostreatus and Pleurotus cystidiosus). Mycobiology 2015, 43, 423–434. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, M.; Abdullah, N.; Ahmed, K.U.; Bhuyan, M.H.M.B. Yield and nutritional composition of oyster mushroom strains newly introduced in Bangladesh. Pesqui. Agropecu. Bras. 2013, 48, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Kortei, N.K.; Dzogbefia, V.P.; Obodai, M. Assessing the Effect of Composting Cassava Peel Based Substrates on the Yield, Nutritional Quality, and Physical Characteristics of Pleurotus ostreatus (Jacq. ex Fr.) Kummer. Biotechnol. Res. Int. 2014, 2014, 571520. [Google Scholar] [CrossRef] [Green Version]
- Kurtzman, R.H. Mushrooms: Sources for Modern Western Medicine. Micol. Apl. Int. 2005, 17, 21–33. Available online: https://www.redalyc.org/pdf/685/68517202.pdf (accessed on 23 October 2021).
- Mintesnot, B.; Ayalew, A.; Kebede, M. Evaluation of biomass of some invasive weed species as substrate for oyster mushroom (Pleurotus spp.) cultivation. Pak. J. Biol. Sci. 2014, 17, 213–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaworska, G.; Bernaś, E.; Mickowska, B. Effect of production process on the amino acid content of frozen and canned Pleurotus ostreatus mushrooms. Food Chem. 2011, 125, 936–943. [Google Scholar] [CrossRef]
- Nieto, I.; Chegwin, C. Influencia del sustrato utilizado para el crecimiento de hongos comestibles sobre sus características nutraceúticas. Rev. Colomb. Biotecnol. 2010, 12, 169–178. [Google Scholar]
- Akyüz, M.; Kirbaǧ, S. Nutritive value of wild edible and cultured mushrooms. Turk. J. Biol. 2010, 34, 97–102. [Google Scholar] [CrossRef]
- Sopanrao, P.S.; Abrar, A.S.; Manoharrao, T.S.; Vaseem, B.M.M. Nutritional value of Pleurotus ostreatus (jacq.:fr.) Kumm cultivated on different lignocellulosic agro-wastes. Innov. Rom. Food Biotechnol. 2010, 7, 66–76. Available online: http://www.bioaliment.ugal.ro/ejournal.htm (accessed on 7 January 2021).
- La Guardia, M.; Venturella, G.; Venturella, F. On the chemical composition and nutritional value of Pleurotus taxa growing on Umbelliferous plants (Apiaceae). J. Agric. Food Chem. 2005, 53, 5997–6002. [Google Scholar] [CrossRef]
- Dundar, A.; Acay, H.; Yildiz, A. Effect of using different lignocellulosic wastes for cultivation of Pleurotus ostreatus (Jacq.) P. Kumm. on mushroom yield, chemical composition and nutritional value. Afr. J. Biotechnol. 2009, 8. Available online: https://www.ajol.info/index.php/ajb/article/view/59909 (accessed on 23 October 2021).
- Aida, F.M.N.A.; Shuhaimi, M.; Yazid, M.; Maaruf, A.G. Mushroom as a potential source of prebiotics: A review. Trends Food Sci. Technol. 2009, 20, 567–575. [Google Scholar] [CrossRef]
- Aishah, M.S.; Rosli, W.I.W. Effect of different drying techniques on the nutritional values of oyster mushroom (Pleurotus sajor-caju). Sains Malays. 2013, 42, 937–941. [Google Scholar]
- Akindahunsi, A.A.; Oyetayo, F.L. Nutrient and antinutrient distribution of edible mushroom, Pleurotus tuber-regium (fries) singer. LWT-Food Sci. Technol. 2006, 39, 548–553. [Google Scholar] [CrossRef]
- Owen, O.J.; Ngodigha, E.M.; Amakiri, A.O. Proximate composition of heat treated poultry litter (layers). Int. J. Poult. Sci. 2008, 7, 1033–1035. [Google Scholar] [CrossRef] [Green Version]
- Fogarasi, M.; Socaci, S.A.; Dulf, F.V.; Diaconeasa, Z.M.; Fărcas, A.C.; Tofană, M.; Semeniuc, C.A. Bioactive compounds and volatile profiles of five transylvanian wild edible mushrooms. Molecules 2018, 23, 3272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michael, H.W.; Bultosa, G.; Pant, L.M. Nutritional contents of three edible oyster mushrooms grown on two substrates at Haramaya, Ethiopia, and sensory properties of boiled mushroom and mushroom sauce. Int. J. Food Sci. Technol. 2011, 46, 732–738. [Google Scholar] [CrossRef]
- Palacios, I.; Lozano, M.; Moro, C.; D’Arrigo, M.; Rostagno, M.A.; Martínez, J.A.; García-Lafuente, A.; Guillamón, E.; Villares, A. Antioxidant properties of phenolic compounds occurring in edible mushrooms. Food Chem. 2011, 128, 674–678. [Google Scholar] [CrossRef]
- Delgado, B.Y.; Morris Quevedo, H.; de La Cruz, E.R.; Quevedo Morales, Y.; Bermúdez Savón, R.C. Contenido de fenoles totales en extractos de Pleurotus obtenidos con solventes de diferente polaridad Content of total phenols in Pleurotus sp. extracts obtained with solvents of different polarity. Rev. Cuba. Investig. Bioméd. 2013, 32, 121–129. Available online: http://scielo.sld.cu (accessed on 14 March 2021).
- Fogarasi, M.; Diaconeasa, Z.M.; Pop, C.R.; Fogarasi, S.; Semeniuc, C.A.; Fărcaş, A.C.; Țibulcă, D.; Sălăgean, C.D.; Tofană, M.; Socaci, S.A. Elemental composition, antioxidant and antibacterial properties of some wild edible mushrooms from Romania. Agronomy 2020, 10, 1972. [Google Scholar] [CrossRef]
- Hip, S.; Fook, Y.; Swee, K.; Chun, W. Phenolic profiles of selected edible wild mushrooms as affected by extraction solvent, time and temperature. Asian J. Food Agro-Ind. 2009, 2, 392–401. [Google Scholar]
- Yıldız, S.; Yılmaz, A.; Can, Z.; Kılıç, C.; Yıldız, Ü.C. Total Phenolic, Flavonoid, Tannin Contents and Antioxidant Properties of Pleurotus ostreatus and Pleurotus citrinopileatus Cultivated on Various Sawdust. Gida/J. Food 2017, 42, 315–323. [Google Scholar] [CrossRef]
- Llauradó, G.; Morris, H.J.; Lebeque, Y.; Gutiérrez, A.; Fontaine, R.; Bermúdez, R.C.; Perraud-Gaime, I. Phytochemical screening and effects on cell-mediated immune response of Pleurotus fruiting bodies powder. Food Agric. Immunol. 2013, 24, 295–304. [Google Scholar] [CrossRef]
- Lim, H.W.; Yoon, J.H.; Kim, Y.S.; Lee, M.W.; Park, S.Y.; Choi, H.K. Free radical-scavenging and inhibition of nitric oxide production by four grades of pine mushroom (Tricholoma matsutake Sing.). Food Chem. 2007, 103, 1337–1342. [Google Scholar] [CrossRef]
- Yildiz, S.; Yilmaz, A.; Can, Z.; Kılıç, C. Total Phenolics, Flavonoids, Tannin Contents and Antioxidant Properties of Pleurotus ostreatus Cultivated on Different Wastes and Sawdust. Int. J. Second. Metab. 2016, 4, 1–9. [Google Scholar] [CrossRef]
- Rajoriya, A.; Tripathy, S.S.; Gupta, N. In vitro antioxidant activity of selected Ganoderma species found in Odisha, India. Trop. Plant Res. 2015, 2, 72–77. [Google Scholar]
- Wong, F.C.; Chai, T.T.; Tan, S.L.; Yong, A.L. Evaluation of bioactivities and phenolic content of selected edible mushrooms in Malaysia. Trop. J. Pharm. Res. 2013, 12, 1011–1016. [Google Scholar] [CrossRef]
- Kalaras, M.D.; Richie, J.P.; Calcagnotto, A.; Beelman, R.B. Mushrooms: A rich source of the antioxidants ergothioneine and glutathione. Food Chem. 2017, 233, 429–433. [Google Scholar] [CrossRef]
- Taofiq, O.; Calhelha, R.C.; Heleno, S.; Barros, L.; Martins, A.; Santos-Buelga, C.; Queiroz, M.J.R.P.; Ferreira, I.C.F.R. The contribution of phenolic acids to the anti-inflammatory activity of mushrooms: Screening in phenolic extracts, individual parent molecules and synthesized glucuronated and methylated derivatives. Food Res. Int. 2015, 76, 821–827. [Google Scholar] [CrossRef] [Green Version]
- Lalitha, P.; Veena, V.; Vidhyapriya, P.; Lakshmi, P.; Krishna, R.; Sakthivel, N. Anticancer potential of pyrrole (1, 2, a) pyrazine 1, 4, dione, hexahydro 3-(2-methyl propyl) (PPDHMP) extracted from a new marine bacterium, Staphylococcus sp. strain MB30. Apoptosis 2016, 21, 566–577. [Google Scholar] [CrossRef]
- Jena, B.S.; Jayaprakasha, G.K.; Singh, R.P.; Sakariah, K.K. Chemistry and Biochemistry of (−)-Hydroxycitric Acid from Garcinia. J. Agric. Food Chem. 2002, 50, 10–22. [Google Scholar] [CrossRef]
- Saito, M.; Ueno, M.; Ogino, S.; Kubo, K.; Nagata, J.; Takeuchi, M. High dose of Garcinia cambogia is effective in suppressing fat accumulation in developing male Zucker obese rats, but highly toxic to the testis. Food Chem. Toxicol. 2005, 43, 411–419. [Google Scholar] [CrossRef]
- Bellettini, M.B.; Fiorda, F.A.; Maieves, H.A.; Teixeira, G.L.; Ávila, S.; Hornung, P.S.; Júnior, A.M.; Ribani, R.H. Factors affecting mushroom Pleurotus spp. Saudi J. Biol. Sci. 2019, 26, 633–646. [Google Scholar] [CrossRef]
- Cardador-Martínez, A.; Loarca-Piña, G.; Oomah, B.D. Antioxidant Activity in Common Beans (Phaseolus vulgaris L.). J. Agric. Food Chem. 2002, 50, 6975–6980. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1999; pp. 152–178. [Google Scholar] [CrossRef]
- Feregrino-Pérez, A.A.; Berumen, L.C.; García-Alcocer, G.; Guevara-Gonzalez, R.G.; Ramos-Gomez, M.; Reynoso-Camacho, R.; Acosta-Gallegos, J.A.; Loarca-Piña, G. Composition and Chemopreventive Effect of Polysaccharides from Common Beans (Phaseolus vulgaris L.) on Azoxymethane-Induced Colon Cancer. J. Agric. Food Chem. 2008, 56, 8737–8744. [Google Scholar] [CrossRef] [PubMed]
- Oomah, B.D.; Cardador-Martínez, A.; Loarca-Piña, G. Phenolics and antioxidative activities in common beans (Phaseolus vulgaris L.). J. Sci. Food Agric. 2005, 85, 935–942. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Fukumoto, L.R.; Mazza, G. Assessing Antioxidant and Prooxidant Activities of Phenolic Compounds. J. Agric. Food Chem. 2000, 48, 3597–3604. [Google Scholar] [CrossRef]
- Nenadis, N.; Wang, L.-F.; Tsimidou, M.; Zhang, H.-Y. Estimation of Scavenging Activity of Phenolic Compounds Using the ABTS•+ Assay. J. Agric. Food Chem. 2004, 52, 4669–4674. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Agave Bagasse: Barley Straw Proportions | r (%) | BE (%) | Colonization (Day) | Primordial Formation (Day) |
|---|---|---|---|---|---|
| T1 | 1:0 | 10.867 ± 0.395 B | 36.60 ± 1.33 B | 8 ± 0 C | 14 ± 0 B |
| T2 | 3:1 | 13.39 ± 0.323 A | 56.7 ± 1.371 A | 8 ± 0 C | 15.67 ± 2.88 A |
| T3 | 1:1 | 10.629 ± 0.04 B | 51.40 ± 0.28 C | 12 ± 0 B | 16 ± 0 A |
| T4 | 1:3 | 7.64 ± 0.21 C | 40.40 ± 1.11 D | 12.67 ± 1.15 B | 16 ± 0 A |
| T5 | 0:1 | 7.25 ± 0.197 D | 43.73 ± 1.19 D | 14 ± 0 A | 17.33 ± 2.31 A |
| Humidity | Lipids | Total Protein | Ash | Carbohydrates | |
|---|---|---|---|---|---|
| T1 | 84.83 ± 0.13 C | 0.28 ± 0.01 A | 6.50 ± 0.13 A | 1.55 ± 0.06 A | 91.67 ± 0.2 C |
| T2 | 87.13 ± 0.52 B | 0.27 ± 0.02 A | 5.88 ± 0.12 B | 1.58 ± 0.08 A | 92.27 ± 0.55 C |
| T3 | 88.04 ± 0.05 A | 0.17 ± 0.02 B | 4.09 ± 0.18 C | 1.28 ± 0.04 B | 94.46 ± 0.23 B |
| T4 | 87.91 ± 0.57 B | 0.14 ± 0.02 C | 3.67 ± 0.14 D | 1.07 ± 0.04 D | 94.91 ± 0.7 AB |
| T5 | 89.4 4± 0.13 A | 0.28 ± 0.01 A | 2.69 ± 0.42 E | 1.1 5± 0.03 C | 95.88 ± 0.3 A |
| Treatment | Chemical Compound | Use | CAS | Retention Time (min) | Molecular Weight (g/mol) |
|---|---|---|---|---|---|
| T1 | Hydroxycitric acid | Limits the transformation of carbohydrates into fats by inhibiting the enzyme ATP-citratolyase. | 3530-14-1 | 3.82 | 208.123 |
| Palmitic acid | Saturated fatty acid, common in beauty products. | 57-10-3 | 6.2 | 256.4 | |
| Paromomycin | Common oligosaccharide in the use of parasitic infections (antibiotic) | 1263-89-4 | 7.807 | 615.629 | |
| 2-deoxy-D-galactose | Tumor growth inhibitor. | 1949-89-9 | 15.5 | 164.068 | |
| Pyrrole (1, 2, a) pyrazine 1, 4, dione, hexahydro 3- (2-methylpropyl) | Anticancer metabolite. | 5654-86-4 | 17.3 | 210.273 | |
| D-mannose | Useful sugar in urinary infections. | 3458-28-4 | 18.3 | 180.156 | |
| T2 | Palmitic acid | Saturated fatty acid, common in beauty products. | 57-10-3 | 6.2 | 256.4 |
| Pyrrole (1, 2, a) pyrazine 1, 4, dione, hexahydro 3- (2-methylpropyl) | Anticancer metabolite. | 5654-86-4 | 17.3 | 210.273 | |
| T3 | Silicic acid | Chemical compound with silicon, stimulates collagen production, increases hair strength and thickness, helps safely remove plaque with brushing, desiccant. | 3555-45-1 | 2.4 | 96.11 |
| Pyrrole (1, 2, a) pyrazine 1, 4, dione, hexahydro 3- (2-methylpropyl) | Anticancer metabolite. | 5654-86-4 | 17.3 | 210.273 | |
| T4 | Quinoline | Organic compound whose derivatives have antiseptic, antibiotic, amebicide properties. Dyes in shades of blue, yellow, and red are extracted. | 1613-34-9 | 4.0 | 129.16 |
| 2-deoxy-D-galactose | Glucose analog, tumor growth inhibitor. | 1949-89-9 | 15.5 | 164.0685 | |
| T5 | DL arabinose | Monosaccharide used for the in vitro culture of various microorganisms. | 20235-19-2 | 3.27 | 150.13 |
| Mannitol | Alcohol, sweetener, blood plasma substitute, diuretic. | 69-65-8 | 5.23 | 182.172 | |
| 2-deoxy-D-galactose | Glucose analog, tumor growth inhibitor. | 1949-89-9 | 18.3 | 164.0685 | |
| Palmitic acid | Saturated fatty acid, common in beauty products. | 57-10-3 | 20.893 | 256.4 | |
| Oleic acid | Monounsaturated fatty acid, hypotensive. | 112-80-1 | 22.931 | 282.47 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz-Moreno, B.A.; Pérez, A.A.F.; García-Trejo, J.F.; Pérez-García, S.A.; Gutiérrez-Antonio, C. Identification of Secondary Metabolites of Interest in Pleurotus djamor Using Agave tequilana Bagasse. Molecules 2023, 28, 557. https://doi.org/10.3390/molecules28020557
Cruz-Moreno BA, Pérez AAF, García-Trejo JF, Pérez-García SA, Gutiérrez-Antonio C. Identification of Secondary Metabolites of Interest in Pleurotus djamor Using Agave tequilana Bagasse. Molecules. 2023; 28(2):557. https://doi.org/10.3390/molecules28020557
Chicago/Turabian StyleCruz-Moreno, Byanka A., Ana Angélica Feregrino Pérez, Juan Fernando García-Trejo, Sergio Alfonso Pérez-García, and Claudia Gutiérrez-Antonio. 2023. "Identification of Secondary Metabolites of Interest in Pleurotus djamor Using Agave tequilana Bagasse" Molecules 28, no. 2: 557. https://doi.org/10.3390/molecules28020557