
Genome Investigation and Functional Annotation of Lactiplantibacillus plantarum YW11 Revealing Streptin and Ruminococcin-A as Potent Nutritive Bacteriocins against Gut Symbiotic Pathogens

 , ,
, ,  , ,
, ,  , , ,
, , ,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Genomic Characteristics and Functional Annotation of Lp YW 11
2.2. Phage Site Prediction
2.3. Interpretation of Transporter Proteins
2.4. Carbohydrate Active Enzyme
2.5. EPS-Producing Genes in Lp YW11
2.6. Interaction of EPS-Producing Genes
2.7. Comparative Genome Analysis
2.8. Molecular Modelling
2.8.1. Interactome Prediction
2.8.2. Molecular Docking and Dynamic Simulations
2.8.3. Immune Simulations
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain and Culture Condition
4.2. DNA Extraction and Whole Genome Sequencing
4.3. Genomic Investigation
4.4. Genome Comparison
4.5. Functional Annotation
4.6. Bacteriocin Production
4.7. Molecular Modelling
4.8. Interactome Prediction of Pathogenic genes of Gut Bacteria
4.9. Molecular Docking and Dynamic Simulations
4.10. Immune Simulations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sarwar, A.; Aziz, T.; Din, J.; Khalid, A.; Rahman, T.; Daudzai, Z. Pros of Lactic Acid Bacteria in Microbiology: A Review. Biomed. Lett. 2018, 4, 59–66. [Google Scholar]
- Aziz, T.; Sarwar, A.; Naveed, M.; Shahzad, M.; Aqib Shabbir, M.; Anas, S.D.; ud Din, J.; Ali Khan, A.; Sumaira, N.; Haiyang, C.; et al. Bio-Molecular Analysis of Selected food derived Lactiplantibacillus strains for CLA Production Reveals possibly a complex mechanism. Food Res. Int. 2022, 154, 111031. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.J.; Park, H.J.; Cha, M.G.; Park, E.; Won, S.M.; Ganesan, R.; Gupta, H.; Gebru, Y.A.; Sharma, S.P.; Lee, S.B.; et al. The Lactobacillus as a Probiotic: Focusing on Liver Diseases. Microorganisms 2022, 10, 288. [Google Scholar] [CrossRef]
- Stefanovic, E.; Fitzgerald, G.; McAuliffe, O. Advances in the genomics and metabolomics of dairy lactobacilli: A review. Food Microbiol. 2017, 61, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Yonekura, S.; Okamoto, Y.; Okawa, T.; Hisamitsu, M.; Chazono, H.; Kobayashi, K.; Sakurai, D.; Horiguchi, S.; Hanazawa, T. Effects of daily intake of Lactobacillus paracasei strain KW3110 on Japanese cedar pollinosis. Allergy Asthma Proc. 2009, 30, 397–405. [Google Scholar] [CrossRef]
- Zhang, Z.; Lv, J.; Pan, L.; Zhang, Y. Roles, and applications of probiotic Lactobacillus strains. Appl. Microbiol. Biotechnol. 2018, 102, 8135–8143. [Google Scholar] [CrossRef]
- Kwon, H.K.; Lee, C.G.; So, J.S.; Chae, C.S.; Hwang, J.S.; Sahoo, A.; Nam, J.H.; Rhee, J.H.; Hwang, K.C.; Im, S.H. Generation of regulatory dendritic cells and CD4+Foxp3+ T cells by probiotics administration suppresses immune disorders. Proc. Natl. Acad. Sci. USA 2010, 107, 2159–2164. [Google Scholar] [CrossRef] [Green Version]
- Hojsak, I.; Snovak, N.; Abdovic, S.; Szajewska, H.; Misak, Z.; Kolacek, S. Lactobacillus GG in the prevention of gastrointestinal and respiratory tract infections in children who attend day care centers: A randomized, double-blind, placebo-controlled trial. Clin. Nutr. 2010, 29, 312–316. [Google Scholar] [CrossRef]
- Joo, H.M.; Hyun, Y.J.; Myoung, K.S.; Ahn, Y.T.; Lee, J.H.; Huh, C.S.; Han, M.J.; Kim, D.H. Lactobacillus johnsonii HY7042 ameliorates Gardnerella vaginalis-induced vaginosis by killing Gardnerella vaginalis and inhibiting NF-κB activation. Int. Immunopharmacol. 2011, 11, 1758–1765. [Google Scholar] [CrossRef]
- Denkova, Z.; Krastanov, A. Development of New Products: Probiotics and Probiotic Foods; IntechOpen: London, UK, 2012; Available online: www.intechopen.com (accessed on 29 October 2022).
- Wang, Y.; Liang, Q.; Lu, B.; Shen, H.; Liu, S.; Shi, Y.; Leptihn, S.; Li, H.; Wei, J.; Liu, C.; et al. Whole-genome analysis of probiotic product isolates reveals the presence of genes related to antimicrobial resistance, virulence factors, and toxic metabolites, posing potential health risks. BMC Genom. 2021, 22, 210. [Google Scholar] [CrossRef]
- Peng, X.; Ed-Dra, A.; Yue, M. Whole genome sequencing for the risk assessment of probiotic lactic acid bacteria. Crit. Rev. Food Sci. Nutr. 2022, 3, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Surve, S.; Shinde, D.B.; Kulkarni, R. Isolation, characterization, and comparative genomics of potentially probiotic Lactiplantibacillus plantarum strains from Indian foods. Sci. Rep. 2022, 12, 1940. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.; Hart, A.; Kamm, M.; Stagg, A.; Knight, S. CMechanisms of action of probiotics: Recent advances. Inflamm. Bowel Dis. 2009, 15, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, R.; Altermann, E.; Anderson, R.C.; McNabb, W.C.; Moughan, P.J.; Roy, N.C. The role of cell surface architecture of lactobacilli in host-microbe interactions in the gastrointestinal tract. Mediators Inflamm. 2013, 2013, 237921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velez, M.P.; Keersmaecker, D.S.C.; Vanderleyden, J. Adherence factors of Lactobacillus in the human gastrointestinal tract. FEMS Microbiol. Lett. 2007, 276, 140–148. [Google Scholar] [PubMed] [Green Version]
- Hörmannsperger, G.; Haller, D. Molecular crosstalk of probiotic bacteria with the intestinal immune system: Clinical relevance in the context of inflammatory bowel disease. Int. J. Med. Microbiol. 2010, 300, 63–73. [Google Scholar] [CrossRef]
- Ramchandran, L.; Shah, N.P. Effect of exopolysaccharides on the proteolytic and Angiotensin-I converting enzyme-inhibitory activities and textural and rheological properties of low-fat yogurt during refrigerated storage. J. Dairy Sci. 2009, 92, 895–906. [Google Scholar] [CrossRef] [Green Version]
- Rajoka, M.S.R.; Wu, Y.; Mehwish, H.M.; Bansal, H.; Zhao, L. Lactobacillus exopolysaccharides: New perspectives on engineering strategies, physiochemical functions, and immunomodulatory effects on host health. Trends Food Sci. Technol. 2020, 103, 36–48. [Google Scholar] [CrossRef]
- Oleksy, M.; Klewicka, E. Exopolysaccharides produced by Lactobacillus sp.: Biosynthesis and applications. Crit. Rev. Food Sci. Nutr. 2018, 11, 450–462. [Google Scholar]
- Angelin, J.; Kavitha, M. Exopolysaccharides from probiotic bacteria and their health potential. Int. J. Biol. Macromol. 2020, 162, 853–865. [Google Scholar] [CrossRef]
- Fei, Y.T.; Liu, D.M.; Luo, T.H.; Chen, G.; Wu, H.; Li, L.; Yu, Y.G. Molecular characterization of Lactobacillus plantarum DMDL 9010, a strain with efficient nitrite degradation capacity. PLoS ONE 2014, 25, e113792. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.M.; Guo, J.; Zeng, X.A.; Sun, D.W.; Brennan, C.S.; Zhou, Q.X.; Jin, S.Z. The probiotic role of Lactobacillus plantarum in reducing risks associated with cardiovascular disease. Int. J. Food Sci. Technol. 2017, 52, 127–136. [Google Scholar] [CrossRef]
- Saenz, Y.; Rojo-Bezares, B.; Navarro, L.; Díez, L.; Somalo, S.; Zarazaga, Z.; Torres, C. Genetic diversity of the pln locus among oenological Lactobacillus plantarum strains. Int. J. Food Microbiol. 2009, 134, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Seddik, H.A.; Bendali, F.; Gancel, F.; Fliss, I.; Spano, G.; Drider, D. Lactobacillus plantarum and Its Probiotic and Food Potentialities. Probiotics Antimicrob. Proteins 2017, 9, 111–122. [Google Scholar] [CrossRef]
- Devi, S.M.; Halami, P.M. Genetic variation of pln loci among probiotic Lactobacillus plantarum group strains with antioxidant and cholesterol-lowering ability. Probiotics Antimicrob. Proteins 2019, 11, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Aziz, T.; Naveed, M.; Sarwar, A.; Makhdoom, S.I.; Mughal, M.S.; Ali, U.; Yang, Z.; Shahzad, M.; Sameeh, M.Y.; Alruways, M.W.; et al. Functional Annotation of Lactiplantibacillus plantarum 13-3 as a Potential Starter Probiotic Involved in the Food Safety of Fermented Products. Molecules 2022, 27, 5399. [Google Scholar] [CrossRef]
- Vries, M.; Vaughan, E.E.; Kleerebezem, M.; Vos, W. Lactobacillus plantarum-survival, functional and potential probiotic properties in the human intestinal tract. Int. Dairy J. 2006, 16, 1018–1028. [Google Scholar] [CrossRef]
- Aziz, T.; Sarwar, A.; ud Din, J.; Al Dalali, S.; Ali Khan, A.; Din, Z.U.; Yang, Z. Biotransformation of linoleic acid into different metabolites by food derived Lactobacillus plantarum 12-3 and in silico characterization of relevant reactions. Food Res. Int. 2021, 147, 110470. [Google Scholar] [CrossRef]
- Aziz, T.; Sarwar, A.; Fahim, M.; Al Dalali, S.; Din, Z.U.; ud Din, J.; Xin, Z.; Jian, Z.; Pacheco Fill, T.; Yang, Z. In silico characterization of linoleic acid biotransformation to rumenic acid in food derived Lactobacillus plantarum YW11. Acta Biochim. Pol. 2020, 7, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Aziz, T.; Sarwar, A.; Fahim, M.; Din, J.U.; Al-Dalali, S.; Din, Z.U.; Ali Khan, A.; Jian, Z.; Yang, Z. Dose-dependent Production of Linoleic Acid Analogues in food derived Lactobacillus plantarum K25 and in silico Characterization of Relevant Reactions. Acta Biochim. Pol. 2020, 67, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Aziz, T.; Sarwar, A.; Fahim, M.; Al-Dalali, S.; Din, Z.U.; ud Din, J.; Fill, P.T.; Yang, Z. Conversion of linoleic acid to different fatty acid metabolites by Lactobacillus plantaraum 13-3 and in silico characterization of the prominent reactions. J. Chil. Chem. Soc. 2020, 65, 4879–4884. [Google Scholar] [CrossRef]
- Yunyun, J.; Jian, Z.; Xiao, Z.; Wen, Z.; Zhijian, Y.; Chao, C.; Yang, Z. Complete genome sequencing of exopolysaccharide-producing Lactobacillus plantarum K25 provides genetic evidence for the probiotic functionality and cold endurance capacity of the strain. Bioscience 2018, 82, 1225–1233. [Google Scholar]
- Wang, J.; Zhao, X.; Tian, Z.; Yang, Y.; Yang, Z. Characterization of an exopolysaccharide produced by Lactobacillus plantarum YW11 isolated from Tibet Kefir. Carbohydr. Polym. 2015, 10, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Churro, C.; Pereira, P.; Vasconcelos, V.; Valério, E. Species-specific real-time PCR cell number quantification of the bloom-forming cyanobacterium Planktothrix agardhii. Arch Microbiol. 2012, 194, 749–757. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
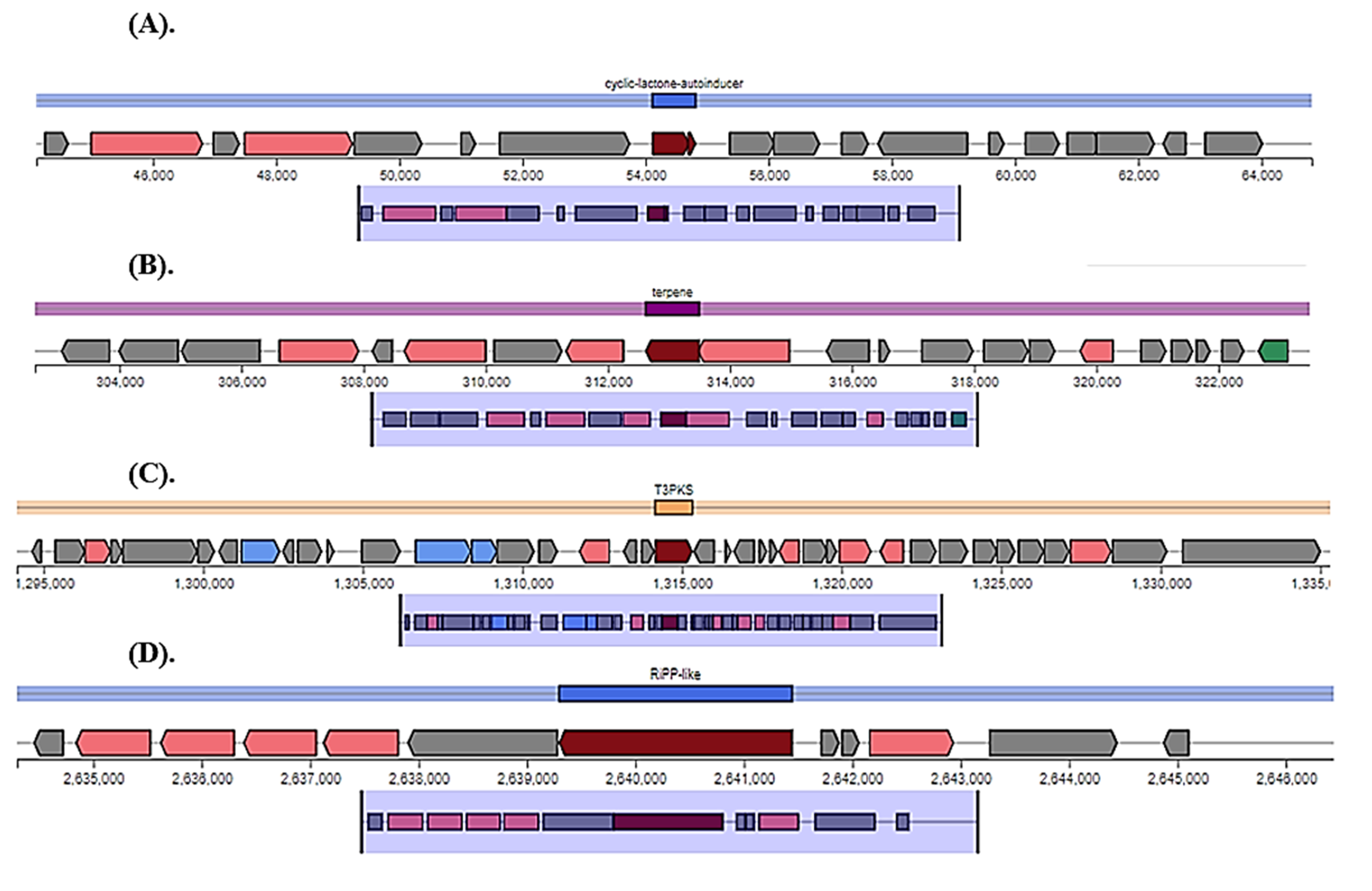{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
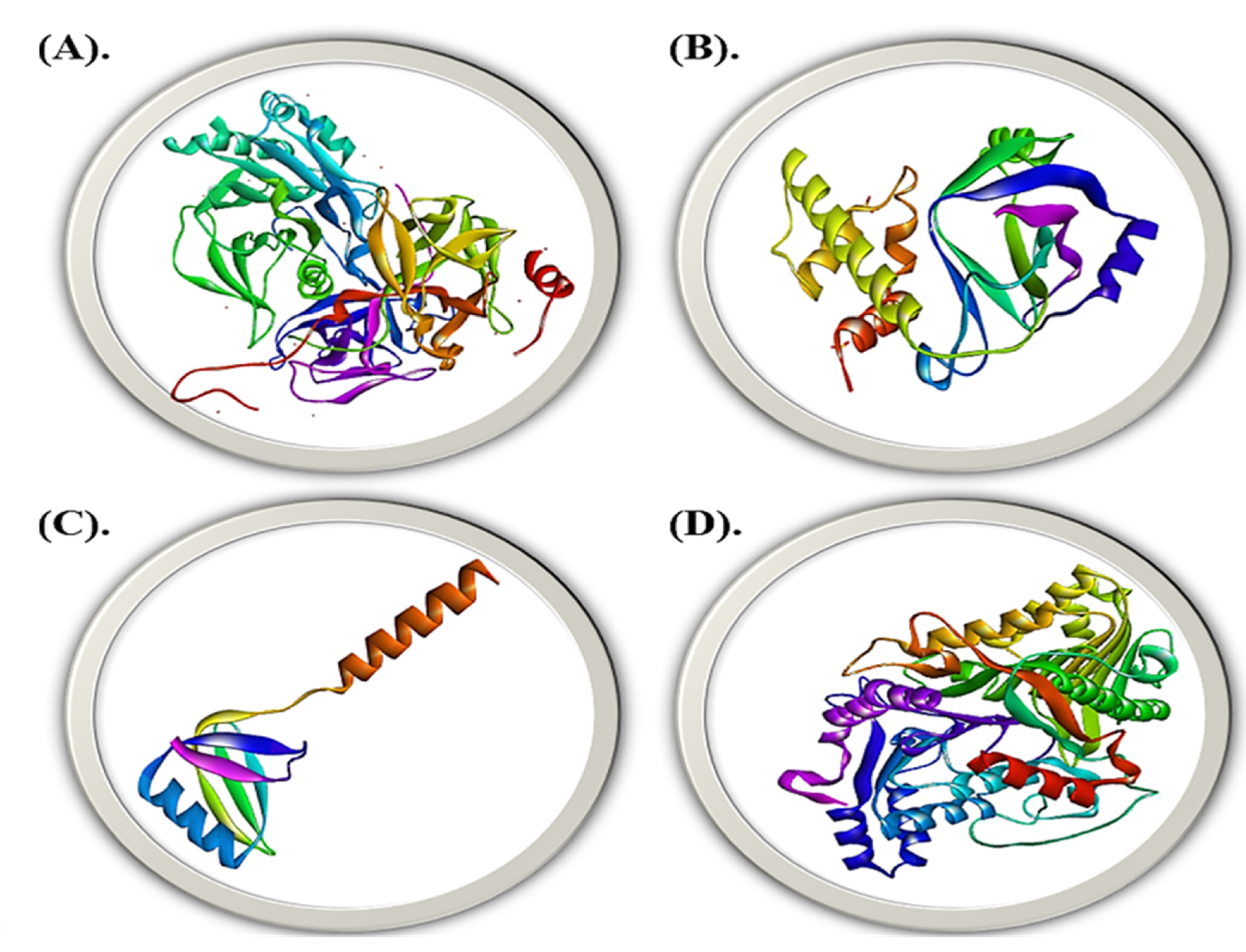{kind=link}
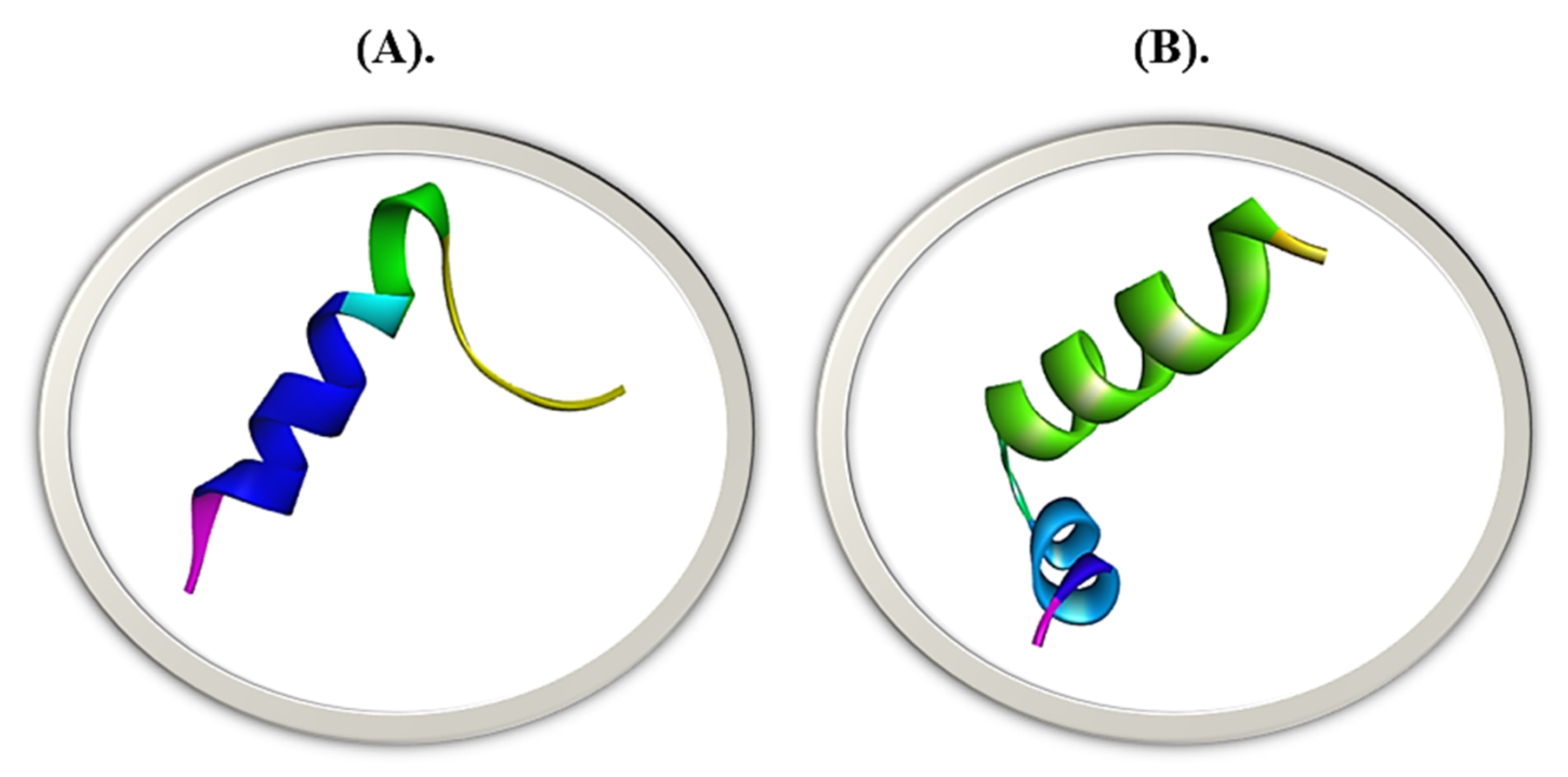{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TCDB ID | Protein Name | CELLO |
|---|---|---|
| Q13U92 | Putative membrane-anchored cell sub-family protein | Extracellular |
| WP_060331514.1 | autotransporter-associated beta structure protein | Extracellular |
| CAH35630.1 | putative outer membrane protein | Extracellular |
| Q79FV6 | PE-PGRS FAMILY PROTEIN | Extracellular |
| Q86IX4 | Nsp1_C domain-containing protein | Extracellular |
| WP_010035418.1 | FG-GAP repeat protein | Extracellular |
| B1Z6U1 | Filamentous haemagglutinin family protein | Extracellular |
| B3FNS7 | Trimeric autotransporter adhesin protein | Extracellular |
| XP_021194905.1 | papilin isoform X5 | Extracellular |
| Q86AS3 | EGF-like domain-containing protein | Extracellular |
| P46531.4 | Neurogenic locus notch homolog protein | Extracellular |
| A2VEC9.2 | SCO-spondin protein | Extracellular |
| P06620.1 | Ice nucleation protein | Extracellular |
| G0SBQ3.1 | Nucleoporin NSP1 | Extracellular |
| B0FXJ3 | Ice nucleation protein | Extracellular |
| WP_057332199.1 | PE family protein | Extracellular |
| L7VCL1 | PE-PGRS family protein | Extracellular |
| P10079 | Fibropellin-1 | Extracellular |
| Q03650 | Cysteine-rich, acidic integral membrane protein | Extracellular |
| G0SAK3 | Nucleoporin NUP145 | Extracellular |
| WP_084871275.1 | carbohydrate-binding domain protein | Extracellular |
| O00468 | Agrin | Extracellular |
| D3KYQ3 | Macronuclear nucleoporin protein | Extracellular |
| L7V457 | PE-PGRS family protein | Extracellular |
| A7SCE9 | Predicted protein (Fragment) | Extracellular |
| P37198 | Nuclear pore glycoprotein | Extracellular |
| E1ZJE6 | Putative uncharacterized protein | Extracellular |
| WP_010044366.1 | hypothetical protein | Extracellular |
| P35658 | Nuclear pore complex protein | Extracellular |
| Q749L8 | Cytochrome c | Extracellular |
| WP_010044253.1 | hypothetical protein | Extracellular |
| AVM72784.1 | magnetosome membrane-specific protein | Extracellular |
| A8CG34 | Nuclear envelope pore membrane protein | Extracellular |
| A6NF01.2 | Putative nuclear envelope pore membrane protein | Extracellular |
| P71187 | TrbL protein | Extracellular |
| Q8H384 | Cadmium selective transporter protein | Extracellular |
| A3M3H0 | Adhesin Ata autotransporter | Extracellular |
| MBC7395593.1 | flagellar hook-length control protein | Extracellular |
| XP_003062523.1 | predicted protein | Extracellular |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aziz, T.; Naveed, M.; Makhdoom, S.I.; Ali, U.; Mughal, M.S.; Sarwar, A.; Khan, A.A.; Zhennai, Y.; Sameeh, M.Y.; Dablool, A.S.; et al. Genome Investigation and Functional Annotation of Lactiplantibacillus plantarum YW11 Revealing Streptin and Ruminococcin-A as Potent Nutritive Bacteriocins against Gut Symbiotic Pathogens. Molecules 2023, 28, 491. https://doi.org/10.3390/molecules28020491
Aziz T, Naveed M, Makhdoom SI, Ali U, Mughal MS, Sarwar A, Khan AA, Zhennai Y, Sameeh MY, Dablool AS, et al. Genome Investigation and Functional Annotation of Lactiplantibacillus plantarum YW11 Revealing Streptin and Ruminococcin-A as Potent Nutritive Bacteriocins against Gut Symbiotic Pathogens. Molecules. 2023; 28(2):491. https://doi.org/10.3390/molecules28020491
Chicago/Turabian StyleAziz, Tariq, Muhammad Naveed, Syeda Izma Makhdoom, Urooj Ali, Muhammad Saad Mughal, Abid Sarwar, Ayaz Ali Khan, Yang Zhennai, Manal Y. Sameeh, Anas S. Dablool, and et al. 2023. "Genome Investigation and Functional Annotation of Lactiplantibacillus plantarum YW11 Revealing Streptin and Ruminococcin-A as Potent Nutritive Bacteriocins against Gut Symbiotic Pathogens" Molecules 28, no. 2: 491. https://doi.org/10.3390/molecules28020491