
A Novel Cold-Adapted and High-Alkaline Alginate Lyase with Potential for Alginate Oligosaccharides Preparation

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
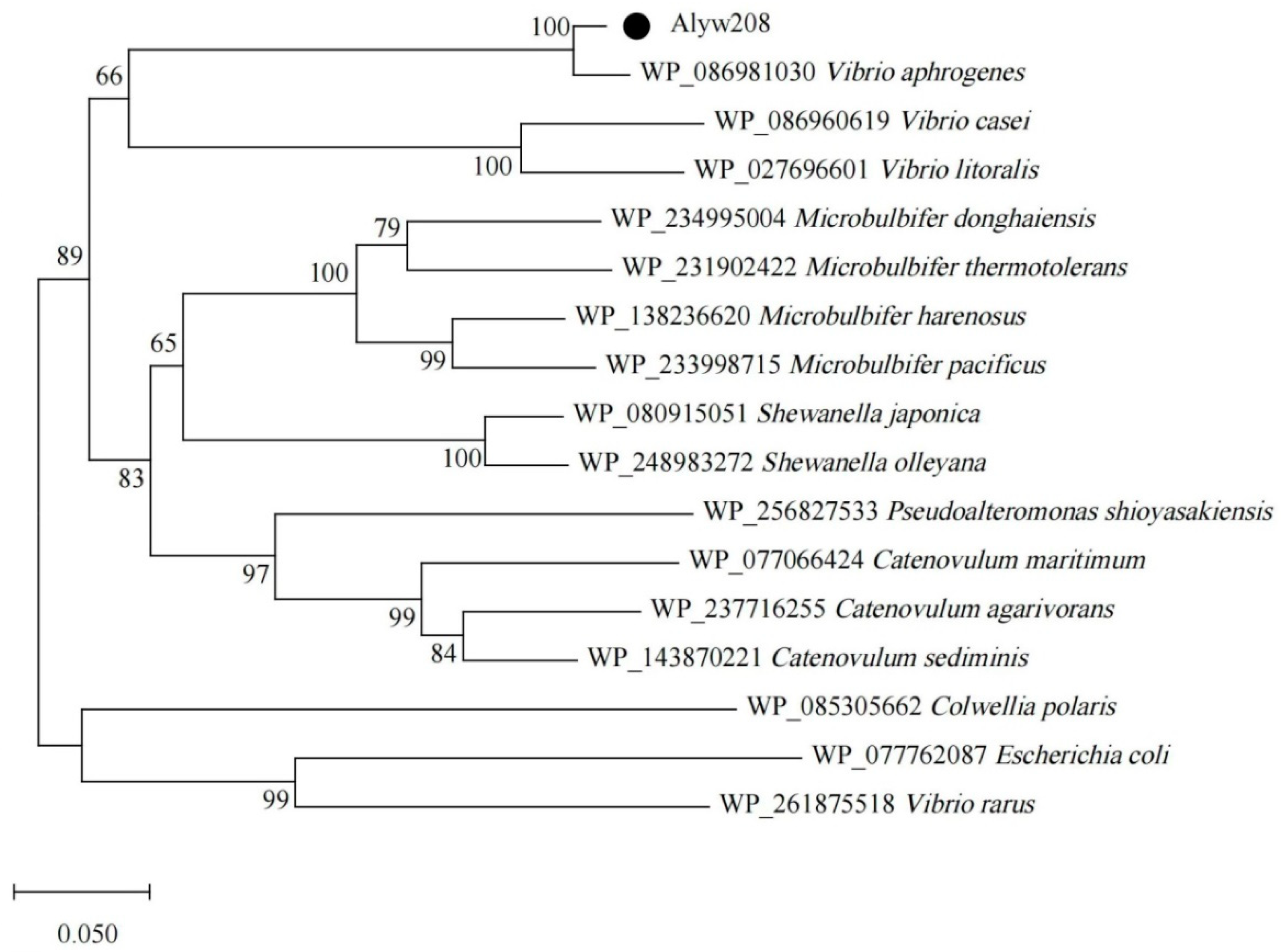
2.1. Bioinformatics Analysis of the Alginate Lyase Alyw208
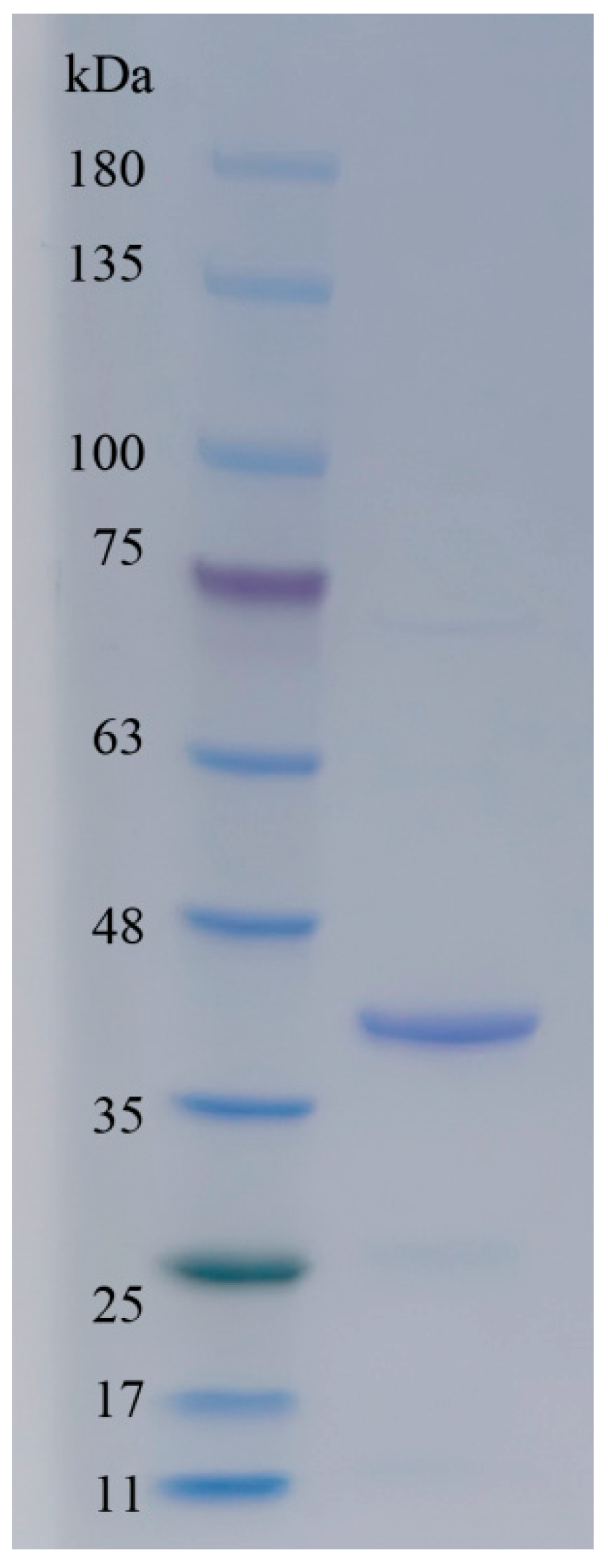
2.2. Secretory Expression and Purification of Alyw208
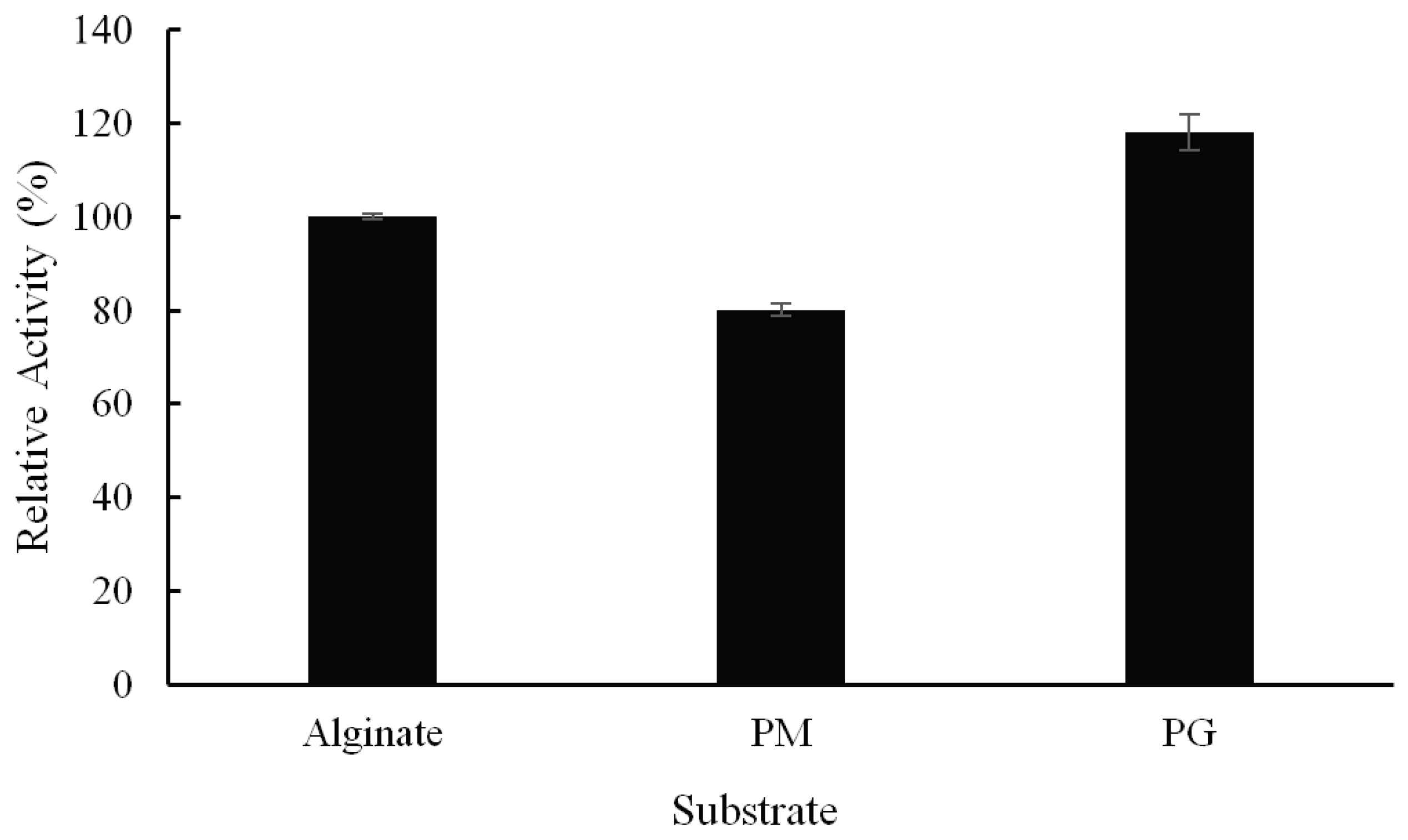
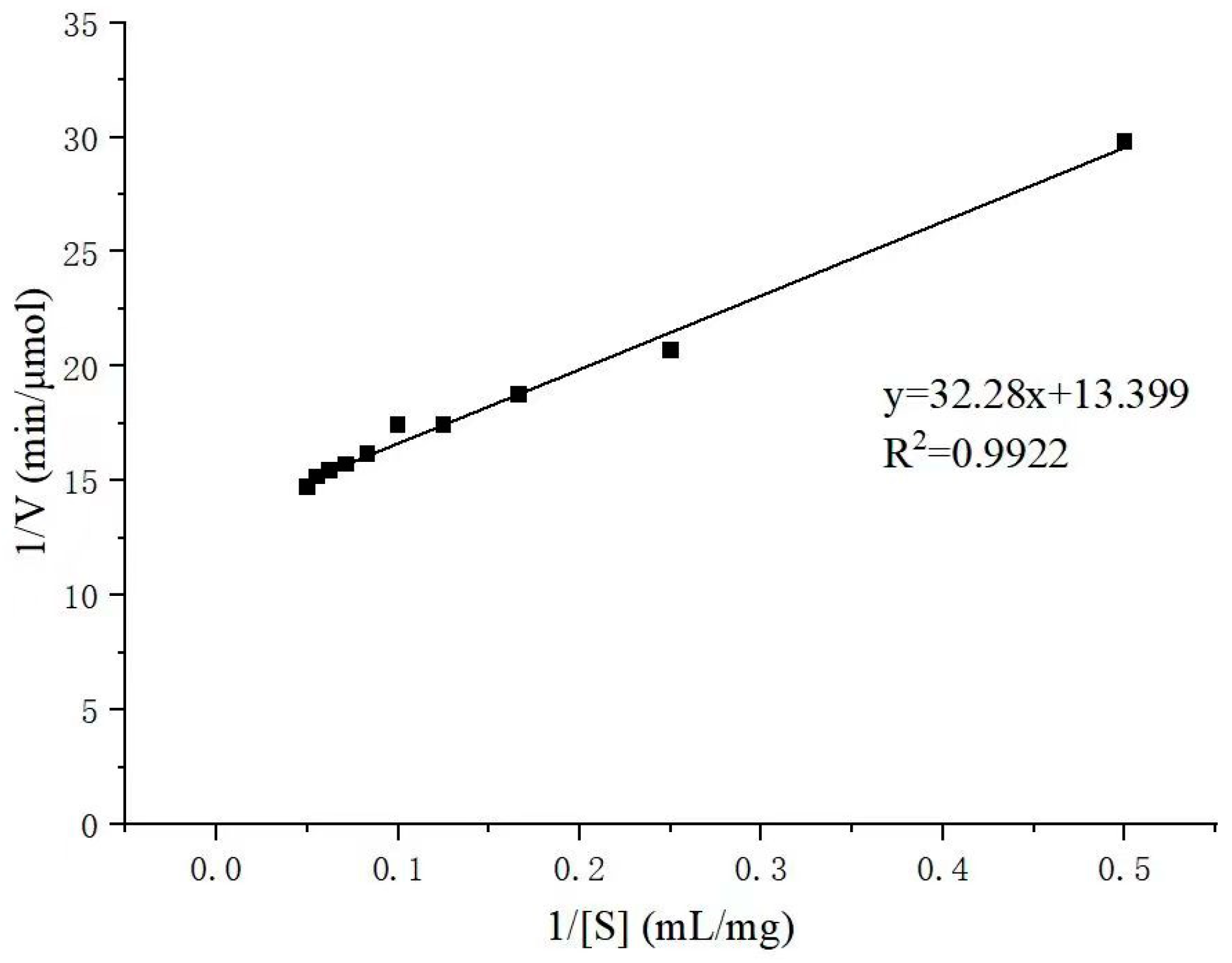
2.3. Substrate Specificity and Kinetic Parameters of Alyw208
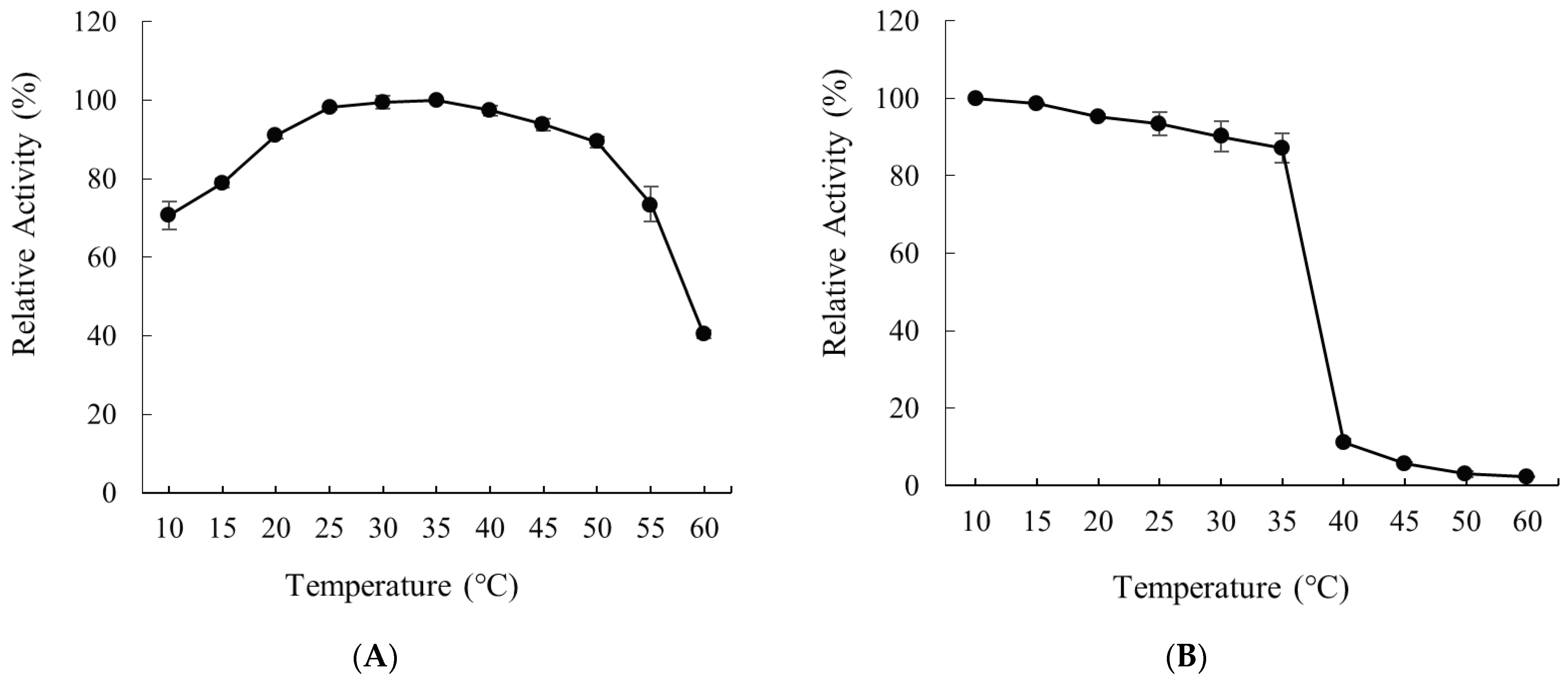
2.4. Effects of Temperature on the Activity and Stability of Recombinant Alyw208
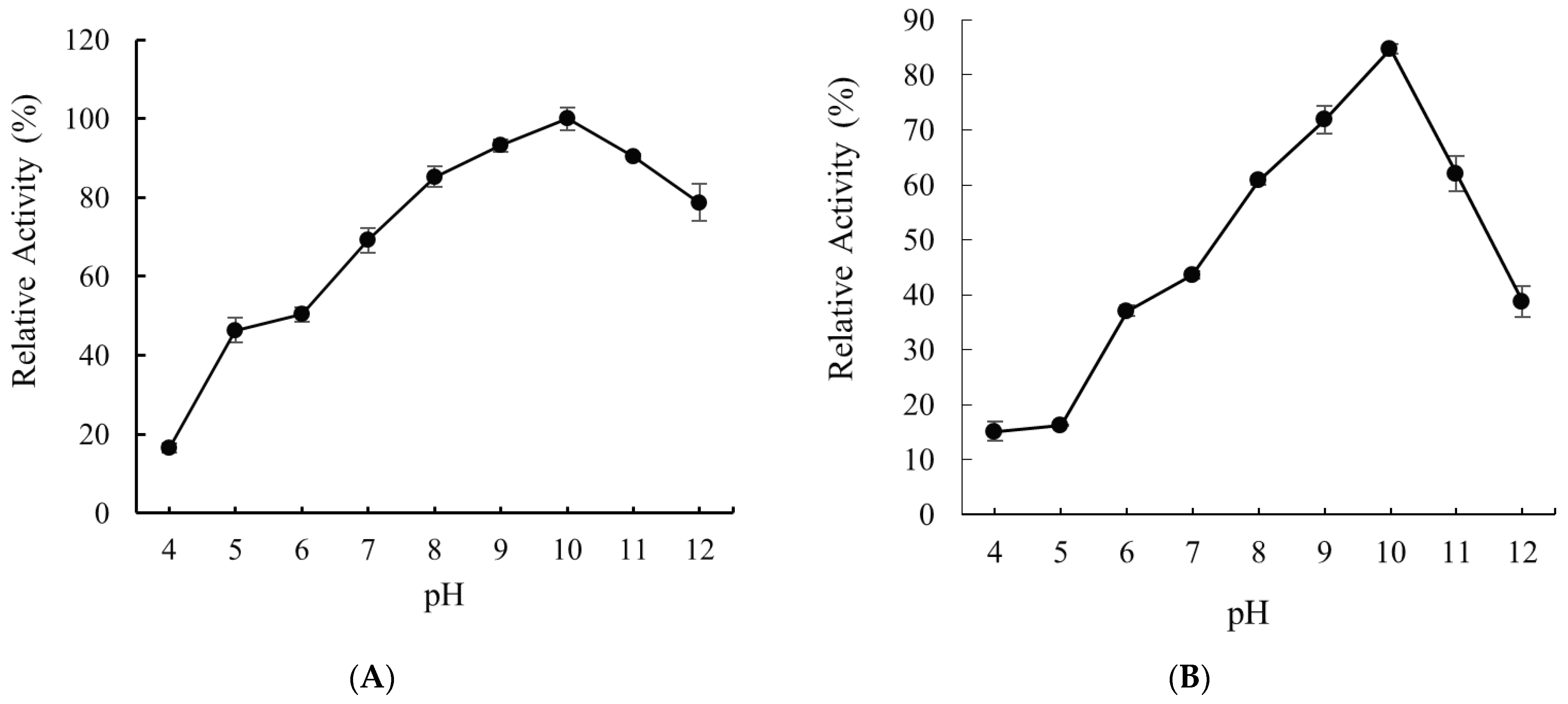
2.5. Effects of pH on the Activity and Stability of Recombinant Alyw208
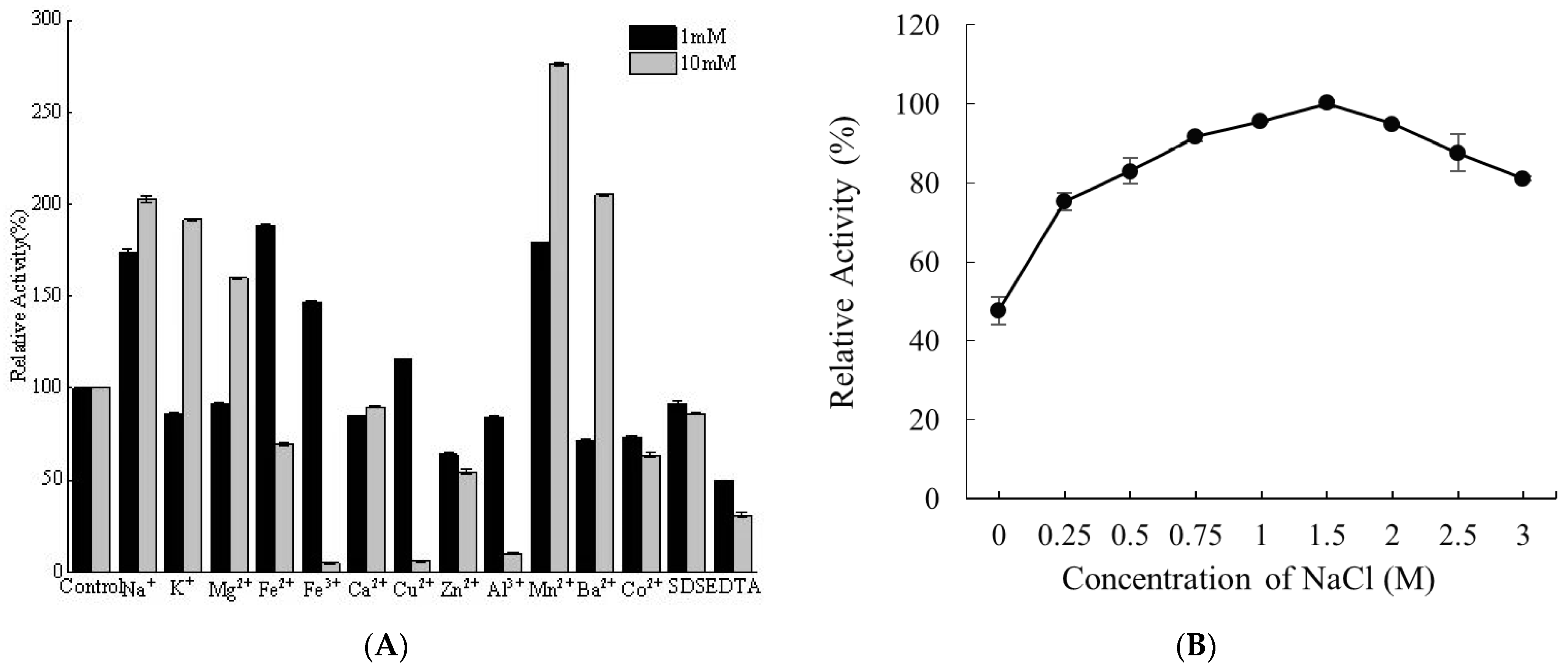
2.6. Effects of Metal Ions and NaCl on Alyw208 Activity
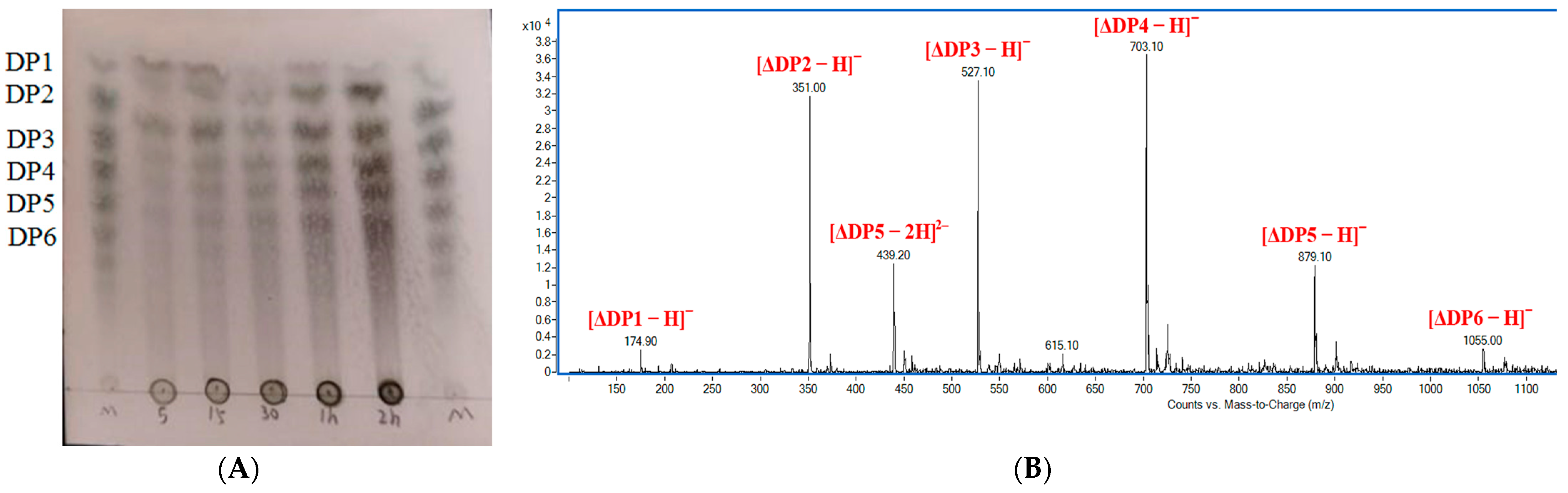
2.7. Action Pattern and Final Products of Alyw208
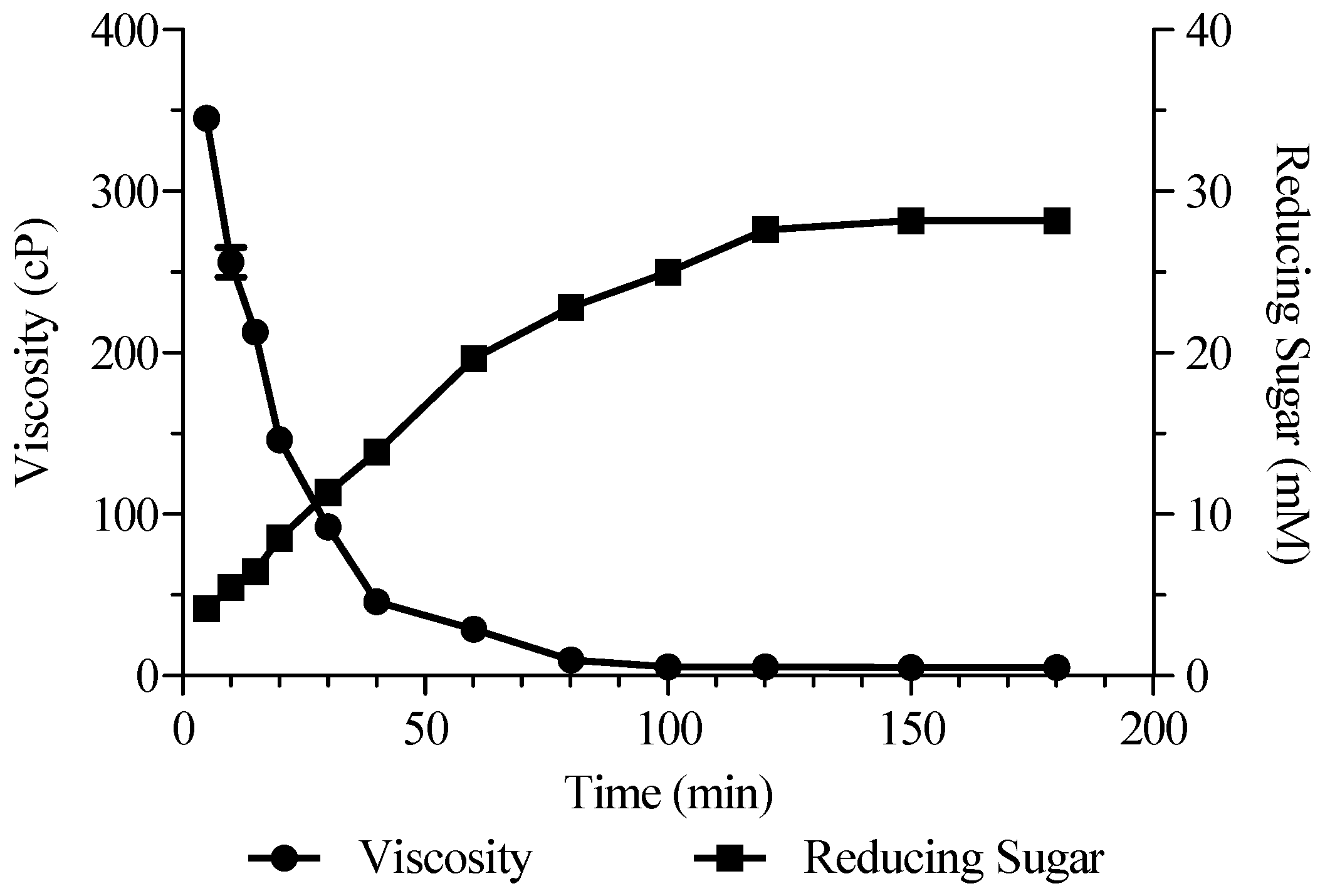
2.8. Preparation of AOs by Alyw208
3. Materials and Methods
3.1. Strains, Media, and Materials
3.2. Bioinformatics Analysis of Alyw208
3.3. Expression and Purification of Recombinant Alyw208
3.4. Substrate Specificity and Kinetic Constants
3.5. Effects of Temperature and pH on the Activity and Stability of Alyw208
3.6. Effects of Ions and NaCl on Alyw208 Activity
3.7. Final Product Analysis of Alyw208
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Florez-Fernandez, N.; Torres, M.D.; Gonzalez-Munoz, M.J.; Dominguez, H. Recovery of bioactive and gelling extracts from edible brown seaweed Laminaria ochroleuca by non-isothermal autohydrolysis. Food Chem. 2019, 277, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Popper, Z.A.; Michel, G.; Hervé, C.; Domozych, D.S.; Willats, W.G.; Tuohy, M.G.; Kloareg, B.; Stengel, D.B. Evolution and diversity of plant cell walls: From algae to flowering plants. Annu. Rev. Plant Biol. 2011, 62, 567–590. [Google Scholar] [CrossRef] [PubMed]
- Szekalska, M.; Puciłowska, A.; Szymańska, E.; Ciosek, P.; Winnicka, A.K. Alginate-current use and future perspectives in pharmaceutical and biomedical applications. Int. J. Polym. Sci. 2016, 2016, 7697031. [Google Scholar] [CrossRef]
- Gacesa, P. Alginates. Carbohydr. Polym. 1988, 8, 161–182. [Google Scholar] [CrossRef]
- Li, S.Y.; Wang, Z.P.; Wang, L.N.; Peng, J.X.; Wang, Y.N.; Han, Y.T.; Zhao, S.F. Combined enzymatic hydrolysis and selective fermentation for green production of alginate oligosaccharides from Laminaria japonica. Bioresour. Technol. 2019, 281, 84–89. [Google Scholar] [CrossRef]
- Liu, J.; Yang, S.Q.; Li, X.T.; Yan, Q.J.; Reaney, M.J.T.; Jiang, Z.Q. Alginate oligosaccharides: Production, biological activities, and potential applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1859–1881. [Google Scholar] [CrossRef]
- Zhao, X.; Li, B.; Xue, C.; Sun, L. Effect of molecular weight on the antioxidant property of low molecular weight alginate from Laminaria japonica. J. Appl. Phycol. 2012, 24, 295–300. [Google Scholar] [CrossRef]
- Li, S.Y.; He, N.N.; Wang, L.N. Efficiently anti-obesity effects of unsaturated alginate oligosaccharides (UAOS) in high-fat diet (HFD)-fed mice. Mar. Drugs 2019, 17, 540. [Google Scholar] [CrossRef]
- Terakado, S.; Ueno, M.; Tamura, Y.; Toda, N.; Yoshinaga, M.; Otsuka, K.; Numabe, A.; Kawabata, Y.; Murota, I.; Sato, N.; et al. Sodium alginate oligosaccharides attenuate hypertension and associated kidney damage in Dahl salt-sensitive rats fed a high-salt diet. Clin. Exp. Hypertens. 2012, 34, 99–106. [Google Scholar] [CrossRef]
- Han, Y.; Zhang, L.; Yu, X.; Wang, S.D.; Xu, C.Y.; Yin, H.; Wang, S.J. Alginate oligosaccharide attenuates alpha 2,6-sialylation modification to inhibit prostate cancer cell growth via the Hippo/YAP pathway. Cell Death Dis. 2019, 10, 374. [Google Scholar] [CrossRef]
- Eftekharzadeh, B.; Khodagholi, F.; Abdi, A.; Maghsoudi, N. Alginate protects NT2 neurons against H2O2-induced neurotoxicity. Carbohyd. Polym. 2010, 79, 1063–1072. [Google Scholar] [CrossRef]
- Fang, W.S.; Bi, D.C.; Zheng, R.J.; Cai, N.; Xu, H.; Zhou, R.; Lu, J.; Wan, M.; Xu, X. Identification and activation of TLR4-mediated signaling pathways by alginate-derived guluronate oligosaccharide in RAW264.7 macrophages. Sci. Rep. 2017, 7, 1663. [Google Scholar] [CrossRef]
- Cherry, P.; Yadav, S.; Strain, C.R.; Allsopp, P.J.; Stanton, C. Prebiotics from seaweeds: An ocean of opportunity? Mar. Drugs 2019, 17, 327. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, M.; Rauf, A.; Khalil, A.A.; Shan, Z.; Chen, C.; Rengasamy, K.R.R.; Wan, C. Process and applications of alginate oligosaccharides with emphasis on health beneficial perspectives. Crit. Rev. Food Sci. Nutr. 2021, 13, 303–329. [Google Scholar] [CrossRef]
- Xu, X.; Yoshiko, I.; Yoshie, K.; Tatsuya, O.; Tsuyoshi, M. Root growth-promoting activity of unsaturated oligomeric uronates from alginate on carrot and rice plants. Biosci. Biotechnol. Biochem. 2003, 67, 2022–2025. [Google Scholar] [CrossRef]
- Ma, L.K.; Zhang, B.; Deng, S.G.; Xie, C. Comparison of the cryoprotective effects of trehalose, alginate, and its oligosaccharides on peeled shrimp (Litopenaeus vannamei) during frozen storage. J. Food Sci. 2015, 80, C540–C546. [Google Scholar] [CrossRef]
- Liu, S.; Su, C.; Guo, J.; Meng, F.; Fan, D.; Wang, F.; Yang, J.; Wang, J.; Zhou, H. Anti-tobacco mosaic virus effects of silver nanoparticles coated with alginate oligosaccharides prepared through enzymatic degradation. ACS Sustain. Chem. Eng. 2022, 10, 11726–11738. [Google Scholar] [CrossRef]
- Falkeborg, M.; Cheong, L.Z.; Gianfico, C.; Sztukiel, K.M.; Kristensen, K.; Glasius, M.; Xu, X.; Guo, Z. Alginate oligosaccharides: Enzymatic preparation and antioxidant property evaluation. Food Chem. 2014, 164, 185–194. [Google Scholar] [CrossRef]
- Wong, T.Y.; Preston, L.A.; Schiller, N.L. Alginate lyase: Review of major sources and enzyme characteristics, structure-function analysis, biological roles, and applications. Annu. Rev. Microbiol. 2000, 54, 289. [Google Scholar] [CrossRef]
- Kim, H.S.; Lee, C.G.; Lee, E.Y. Alginate lyase: Structure, property, and application. Biotechnol. Bioprocess Eng. 2011, 16, 843–851. [Google Scholar] [CrossRef]
- Jagtap, S.S.; Hehemann, J.H.; Polz, M.F.; Lee, J.K.; Zhao, H. Comparative biochemical characterization of three exolytic oligoalginate lyases from Vibrio splendidus reveals complementary substrate scope, temperature, and pH adaptations. Appl. Environ. Microbiol. 2014, 80, 4207–4214. [Google Scholar] [CrossRef]
- Wang, L.; Li, S.; Yu, W.; Gong, Q. Cloning, overexpression and characterization of a new oligoalginate lyase from a marine bacterium, Shewanella sp. Biotechnol. Lett. 2015, 37, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Wargacki, A.J.; Leonard, E.; Win, M.N.; Regitsky, D.D.; Santos, C.N.S.; Kim, P.B.; Cooper, S.R.; Raisner, R.M.; Herman, A.; Sivitz, A.B.; et al. An engineered microbial platform for direct biofuel production from Brown macroalgae. Science 2012, 335, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Suzuki, K.I.; Inoue, A.; Ojima, T. A novel oligoalginate lyase from abalone, Haliotis discus hannai, that releases disaccharide from alginate polymer in an exolytic manner. Carbohydr. Res. 2006, 341, 1809–1819. [Google Scholar] [CrossRef]
- Li, S.; Wang, L.; Hao, J.; Xing, M.; Sun, J.; Sun, M. Purification and characterization of a new alginate lyase from marine bacterium Vibrio sp. SY08. Mar. Drugs 2017, 15, 1. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Sun, X.M.; Chen, C.; Zhang, X.Y.; Chen, X.L.; Zhang, Y.Z.; Fan, S.J.; Xu, F. A novel alginate lyase: Identification, characterization, and potential application in alginate trisaccharide preparation. Mar. Drugs 2022, 20, 159. [Google Scholar] [CrossRef]
- Zhang, Z.; Tang, L.; Bao, M.; Liu, Z.; Yu, W.; Han, F. Functional characterization of carbohydrate-binding modules in a new alginate lyase, TsAly7B, from Thalassomonas sp. LD5. Mar. Drugs 2019, 18, 25. [Google Scholar] [CrossRef]
- Zhu, B.W.; Chen, M.J.; Yin, H.; Du, Y.G.; Ning, L.M. Enzymatic hydrolysis of alginate to produce oligosaccharides by a new purified endo-type alginate lyase. Mar. Drugs 2016, 14, 108. [Google Scholar] [CrossRef]
- Li, Q.; Hu, F.; Wang, M.; Zhu, B.; Ni, F.; Yao, Z. Elucidation of degradation pattern and immobilization of a novel alginate lyase for preparation of alginate oligosaccharides. Int. J. Biol. Macromol. 2020, 146, 579–587. [Google Scholar] [CrossRef]
- Zhu, B.W.; Tan, H.D.; Qin, Y.Q.; Du, Y.G.; Yin, H. Characterization of a new endo-type polyM-specific alginate lyase from Pseudomonas sp. Biotechnol. Lett. 2015, 37, 409–415. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Bi, X.; Ren, Y.; Han, Q.; Zhou, Y.; Han, Y.; Li, S. Characterization of an alkaline alginate lyase with pH-stable and thermo-tolerance property. Mar. Drugs 2019, 17, 308. [Google Scholar] [CrossRef]
- Yamasaki, M.; Moriwaki, S.; Miyake, O.; Hashimoto, W.; Murata, K.; Mikami, B. Structure and function of a hypothetical Pseudomonas aeruginosa protein PA1167 classified into family PL-7: A novel alginate lyase with a β-sandwich fold. Int. J. Biol. Chem. 2004, 279, 31863–31872. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.X.; Xu, S.S.; Yin, X.J.; Wang, F.L.; Li, Y. Characterization of a new bifunctional and cold-adapted polysaccharide lyase (PL) family 7 alginate lyase from Flavobacterium sp. Mar. Drugs 2020, 18, 388. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Wang, Q.; Lu, M.; Xu, C.; Li, F.; Zhang, R.; Liao, W.; Huang, S. AlgM4: A new salt-activated alginate lyase of the PL7 family with endolytic activity. Mar. Drugs 2018, 16, 120. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Jiang, C.; Xu, J.; Liu, Z.; Mao, X. Characteristics and applications of alginate lyases: A review. Int. J. Biol. Macromol. 2020, 164, 1304–1320. [Google Scholar] [CrossRef] [PubMed]
- Aarstad, O.A.; Tondervik, A.; Sletta, H.; Skjakbraek, G. Alginate sequencing: An analysis of block distribution in alginates using specific alginate degrading enzymes. Biomacromolecules 2012, 13, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Inoue, A.; Mashino, C.; Kodama, T.; Ojima, T. Protoplast preparation from Laminaria japonica with recombinant alginate lyase and cellulase. Mar. Biotechnol. 2011, 13, 256–263. [Google Scholar] [CrossRef]
- Islan, G.A.; Martinez, Y.N.; Illanes, A.; Castro, G.R. Development of novel alginate lyase cross-linked aggregates for the oral treatment of cystic fibrosis. RSC Adv. 2014, 4, 11758. [Google Scholar] [CrossRef]
- Lamppa, J.W.; Griswold, K.E. Alginate lyase exhibits catalysis-independent biofilm dispersion and antibiotic synergy. Antimicrob. Agents Chemother. 2013, 57, 137–145. [Google Scholar] [CrossRef]
- Shiraiwa, Y.; Abe, K.; Sasaki, S.F.; Ikawa, T.; Nisizawa, K. Alginate lyase activities in the extracts from several brown algae. Bot. Mar. 2009, 18, 97–104. [Google Scholar] [CrossRef]
- Ogura, K.; Yamasaki, M.; Yamada, T.; Mikami, B.; Hashimoto, W.; Murata, K. Crystal structure of Family 14 polysaccharide lyase with pH-dependent modes of action. Int. J. Biol. Chem. 2009, 284, 35572–35579. [Google Scholar] [CrossRef]
- Singh, R.P.; Gupta, V.; Kumari, P.; Kumar, M.; Reddy, C.R.K.; Prasad, K.; Jha, B. Purification and partial characterization of an extracellular alginate lyase from Aspergillus oryzae isolated from brown seaweed. J. Appl. Phycol. 2011, 23, 755–762. [Google Scholar] [CrossRef]
- Belik, A.A.; Silchenko, A.S.; Kusaykin, M.I.; Zvyagintseva, T.N.; Ermakova, S.P. Alginate lyases: Substrates, structure, properties, and prospects of application. Russ. J. Bioorg. Chem. 2018, 44, 386–396. [Google Scholar] [CrossRef]
- Pei, X.; Chang, Y.; Shen, J. Cloning, expression and characterization of an endo-acting bifunctional alginate lyase of marine bacterium Wenyingzhuangia fucanilytica. Protein Expr. Purif. 2019, 154, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Yin, H. Alginate lyase: Review of major sources and classification, properties, structure-function analysis and applications. Bioengineered 2015, 6, 125–131. [Google Scholar] [CrossRef]
- Zhu, B.; Li, K.; Wang, W.; Ning, L.; Tan, H.; Zhao, X.; Yin, H. Preparation of trisaccharides from alginate by a novel alginate lyase Alg7A from marine bacterium Vibrio sp. W13. Int. J. Biol. Macromol. 2019, 139, 879–885. [Google Scholar] [CrossRef]
- Inoue, A.; Anraku, M.; Nakagawa, S.; Ojima, T. Discovery of a novel alginate lyase from Nitratiruptor sp. SB155-2 thriving at deep-sea hydrothermal vents and identification of the residues responsible for its heat stability. Int. J. Biol. Chem. 2016, 291, 15551–15563. [Google Scholar] [CrossRef]
- Wang, Z.P.; Cao, M.; Li, B.; Ji, X.F.; Zhang, X.Y.; Zhang, Y.Q.; Wang, H.Y. Cloning, secretory expression and characterization of a unique pH-stable and cold-adapted alginate lyase. Mar. Drugs 2020, 18, 189. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Tan, H.; Qin, Y.; Xu, Q.; Du, Y.; Yin, H. Characterization of a new endo-type alginate lyase from Vibrio sp. W13. Int. J. Biol. Macromol. 2015, 75, 330–337. [Google Scholar] [CrossRef]
- Liu, L.; Wang, Z.; Zheng, Z.; Li, Z.; Ji, X.; Cong, H.; Wang, H. Secretory expression of an alkaline alginate lyase with heat recovery property in Yarrowia lipolytica. Front. Microbiol. 2021, 12, 710533. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Uchimura, K.; Miyazaki, M.; Nogi, Y.; Horikoshi, K. A new high-alkaline alginate lyase from a deep-sea bacterium Agarivorans sp. Extremophiles 2009, 13, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Li, J.; Zhang, X.Y.; Ni, H.D.; Wang, F.B.; Wang, H.Y.; Wang, Z.P. Characterization of a new intracellular alginate lyase with metal ions-tolerant and pH-stable properties. Mar. Drugs 2020, 18, 416. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T.; Okano, S.; Kasai, N. Inactivation of Escherichia coli endotoxin by soft hydrothermal processing. Appl. Environ. Microbiol. 2009, 75, 5058–5063. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, S.; Zhang, Y.; Chen, L. High-level expression of a thermally stable alginate lyase using Pichia pastoris, characterization and application in producing brown alginate oligosaccharide. Mar. Drugs 2018, 16, 158. [Google Scholar] [CrossRef] [PubMed]
- Madzak, C. Yarrowia lipolytica: Recent achievements in heterologous protein expression and pathway engineering. Appl. Microbiol. Biotechnol. 2015, 99, 4559–4577. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Ni, F.; Ning, L.; Sun, Y.; Yao, Z. Cloning and characterization of a new pH-stable alginate lyase with high salt tolerance from marine Vibrio sp. NJ-04. Int. J. Biol. Macromol. 2018, 115, 1063–1070. [Google Scholar] [CrossRef]
- Liu, L.; Guo, J.; Zhou, X.F.; Li, Z.; Zhou, H.X.; Song, W.Q. Characterization and secretory expression of thermostable tannase from Aureobasidium melanogenum T9: Potential candidate for food and agricultural industries. Front. Bioeng. Biotechnol. 2022, 9, 769816. [Google Scholar] [CrossRef]
- Kang, L.X.; Chen, X.M.; Fu, L.; Ma, L.X. Recombinant expression of chitosanase from Bacillus subtilis HD145 in Pichia pastoris. Carbohyd. Res. 2012, 352, 37–43. [Google Scholar] [CrossRef]
- Liu, S.; Wan, D.; Wang, M.; Madzak, C.; Du, G.; Chen, J. Overproduction of pro-transglutaminase from Streptomyces hygroscopicus in Yarrowia lipolytica and its biochemical characterization. BMC Biotechnol. 2015, 15, 75. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhou, J.; Li, X.; Peng, Q.; Lu, H.; Du, Y. Characterization of a new alginate lyase from newly isolated Flavobacterium sp. S20. J. Ind. Microbiol. Biotechnol. 2013, 40, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Uchimura, K.; Miyazaki, M.; Nogi, Y.; Kobayashi, T.; Horikoshi, K. Cloning and sequencing of alginate lyase genes from deep-sea strains of Vibrio and Agarivorans and characterization of a new Vibrio enzyme. Mar. Biotechnol. 2010, 12, 526–533. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Shao, Y.; Jiao, C.; Yang, Q.M.; Weng, H.F.; Xiao, A.F. Characterization and application of an alginate lyase, Aly1281 from marine bacterium Pseudoalteromonas carrageenovora ASY5. Mar. Drugs 2020, 18, 95. [Google Scholar] [CrossRef]
- Hu, X.; Jiang, X.; Hwang, H.M. Purification and characterization of an alginate lyase from marine bacterium Vibrio sp. mutant strain 510-64. Curr. Microbiol. 2006, 53, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Liu, Z.; Jiang, C.; Li, L.; Xue, C.; Mao, X. Biochemical characterization and degradation pattern analysis of a novel PL-6 alginate lyase from Streptomyces coelicolor A3(2). Food Chem. 2020, 323, 126852. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Zhang, Z.; Li, S.; Su, H.; Tang, L.; Tan, Y.; Yu, W.; Han, F. Characterization of a new endo-type polysaccharide lyase (PL) family 6 alginate lyase with cold-adapted and metal ions-resisted property. Int. J. Biol. Macromol. 2018, 120, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, W.; Miyake, O.; Ochiai, A.; Murata, K. Molecular identification of Sphingomonas sp. A1 alginate lyase (A1-IV’) as a member of novel polysaccharide lyase family 15 and implications in alginate lyase evolution. J. Biosci. Bioeng. 2005, 99, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, H.; Horibe, A.; Miki, Y.; Kimura, T.; Tanaka, K.; Nakagawa, T.; Kawamukai, M.; Matsuda, H. Cloning and sequencing analysis of alginate lyase genes from the marine bacterium Vibrio sp. O2. Mar. Biotechnol. 2006, 8, 481–490. [Google Scholar] [CrossRef]
- Inoue, A.; Takadono, K.; Nishiyama, R.; Tajima, K.; Kobayashi, T.; Ojima, T. Characterization of an alginate lyase, FlAlyA, from Flavobacterium sp. strain UMI-01 and its expression in Escherichia coli. Mar. Drugs 2014, 12, 4693–4712. [Google Scholar] [CrossRef]
- Yagi, H.; Isobe, N.; Itabashi, N.; Fujise, A.; Ohshiro, T. Characterization of a long-lived alginate lyase derived from Shewanella species YH1. Mar. Drugs 2018, 16, 4. [Google Scholar] [CrossRef]
- Iwamoto, Y.; Araki, R.; Iriyama, K.I.; Oda, T.; Fukuda, H.; Hayashida, S.; Muramastu, T. Purification and characterization of bifunctional alginate lyase from Alteromonas sp. strain no. 272 and its action on saturated oligomeric substrates. Biosci. Biotechnol. Biochem. 2001, 65, 133–142. [Google Scholar] [CrossRef]
- Zhang, L.L.; Jiang, X.H.; Xiao, X.F.; Zhang, W.X.; Shi, Y.Q.; Wang, Z.P.; Zhou, H.X. Expression and characterization of a novel cold-adapted chitosanase from marine Renibacterium sp. suitable for chitooligosaccharides preparation. Mar. Drugs 2021, 19, 596. [Google Scholar] [CrossRef]
- Kunjukunju, S.; Roy, A.; Shekhar, S.; Kumta, P.N. Cross-linked enzyme aggregates of alginate lyase: A systematic engineered approach to controlled degradation of alginate hydrogel. Int. J. Biol. Macromol. 2018, 115, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Fertah, M.; Belfkira, A.; Dahmane, E.; Taourirte, M.; Brouillette, F. Extraction and characterization of sodium alginate from Moroccan Laminaria digitata brown seaweed. Arab. J. Chem. 2017, 10, S3707–S3714. [Google Scholar] [CrossRef]
- Boyen, C.; Kloareg, B.; Polne-Fuller, M.; Gibor, A. Preparation of alginate lyases from marine molluscs for protoplast isolation in brown algae. Phycologia 1990, 29, 173–181. [Google Scholar] [CrossRef]
- Chen, X.L.; Dong, S.; Xu, F.; Dong, F.; Li, P.Y.; Zhang, X.Y.; Zhou, B.C.; Zhang, Y.Z.; Xie, B.B. Characterization of a new cold-adapted and salt-activated polysaccharide lyase family 7 alginate lyase from Pseudoalteromonas sp. SM0524. Front. Microbiol. 2016, 7, 1120. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.J.; Mikami, B.; Hashimoto, W.; Murata, K. Crystal structure of alginate lyase A1-III from Sphingomonas species A1 at 1.78 angstrom resolution. J. Mol. Biol. 1999, 290, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Li, J.W.; Dong, S.; Song, J.; Li, C.B.; Chen, X.L.; Xie, B.B.; Zhang, Y.Z. Purification and characterization of a bifunctional alginate lyase from Pseudoalteromonas sp. SM0524. Mar. Drugs 2011, 9, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Wang, Q.; Lu, D.; Han, W.; Li, F. A novel bifunctional endolytic alginate lyase with variable alginate-degrading modes and versatile monosaccharide-producing properties. Front. Microbiol. 2018, 9, 167. [Google Scholar] [CrossRef]
- Chen, J.; Hu, Y.; Zhang, L.; Wang, Y.; Wang, S.; Zhang, Y.; Guo, H.; Ji, D.; Wang, Y. Alginate oligosaccharide DP5 exhibits antitumor effects in osteosarcoma patients following surgery. Front. Pharmacol. 2017, 8, 623. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Zhang, M.; Gao, L.; Eromosele, O.; Qiao, Y.; Shi, B. Effects of alginate oligosaccharides with different molecular weights and guluronic to mannuronic acid ratios on glyceollin induction and accumulation in soybeans. J. Food Sci. Technol. 2018, 55, 1850–1858. [Google Scholar] [CrossRef]
- An, Q.D.; Zhang, G.L.; Wu, H.T.; Zhang, Z.C.; Zheng, G.S.; Luan, L.; Murata, Y.; Li, X. Alginate-deriving oligosaccharide production by alginase from newly isolated Flavobacterium sp. LXA and its potential application in protection against pathogens. J. Appl. Microbiol. 2009, 106, 161–170. [Google Scholar] [CrossRef]
- Gietz, R.D.; Schiestl, R.H. High-effciency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Lineweaver, H.; Burk, D. The determination of enzyme dissociation constants. J. Am. Chem. Soc. 1934, 56, 658–666. [Google Scholar] [CrossRef]
- Yagi, H.; Fujise, A.; Itabashi, N.; Ohshiro, T. Characterization of a novel endo-type alginate lyase derived from Shewanella sp. YH1. J. Biochem. 2018, 163, 341–350. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Step | Total Protein (mg) | Total Activity (U) | Specific Activity (U/mg) | Purification Fold | Yield (%) |
|---|---|---|---|---|---|
| Crude enzyme | 32.23 ± 2.59 | 3776 ± 305 | 117.2 | 1 | 100 |
| Ni-IDA agarose | 3.42 ± 0.25 | 2495 ± 234 | 729.5 | 6.22 | 66.1 |
| Enzyme | Source | PL Family | Optimal Temperature (°C) | Optimal pH | Substrate Specificity | Products (DP) | Reference |
|---|---|---|---|---|---|---|---|
| Alyw208 | Vibrio sp. W2 | 7 | 35 | 10.0 | polyG, polyM | 1–6 | This study |
| Alyw201 | Vibrio sp. W2 | 7 | 30 | 8.0 | polyG, polyM | 2–6 | [48] |
| Alyw202 | Vibrio sp. W2 | 7 | 45 | 9.0 | polyG, polyM | 2–4 | [52] |
| Alyw203 | Vibrio sp. W2 | 7 | 45 | 10.0 | − | 1–2 | [50] |
| Algb | Vibrio sp. W13 | 7 | 30 | 8.0 | polyMG, polyG, polyM | 2–5 | [49] |
| Alg7A | Vibrio sp. W13 | 7 | 30 | 7.0 | polyG, polyM, polyMG | 3 | [46] |
| AlySY08 | Vibrio sp. SY08 | − | 40 | 7.6 | polyG, polyM | 2 | [25] |
| Aly510-64 | Vibrio sp. 510-64 | − | 35 | 7.5 | polyG, polyMG | 3 | [63] |
| AlgM4 | Vibrio weizhoudaoensis M0101 | 7 | 30 | 8.5 | polyG, polyM | 2–9 | [34] |
| AlyS02 | Flavobacterium sp. S02 | 7 | 30 | 7.6 | polyG, polyM | 2, 3 | [33] |
| Alg2A | Flavobacterium sp. S20 | 7 | 45 | 8.5 | polyG | 5–7 | [60] |
| PA1167 | Pseudomonas aeruginosa PAO1 | 7 | 40 | 8.5 | polyMG | 2–4 | [32] |
| AlgA | Pseudomonas sp. E03 | 5 | 30 | 8.0 | polyM | 2–5 | [30] |
| OUC-ScCD6 | Streptomyces coelicolor A3(2) | 6 | 50 | 9.0 | polyM, polyG | 2–6 | [64] |
| TsAly6A | Thalassomonas sp. LD5 | 6 | 35 | 8.0 | polyG, polyM | 2, 3 | [65] |
| AlyPL6 | Pedobacter hainanensis NJ-02 | 6 | 45 | 10.0 | polyMG, polyG, polyM | 1–4 | [29] |
| A1-IV’ | Sphingomonas sp. A1 | 15 | 50 | 8.5 | polyM, polyMG | 2, 3 | [66] |
| Alginate Lyase/Source | Relative Activity at 25/20/15/10 °C (%) | Optimal Temperature (°C) | Reference |
|---|---|---|---|
| Algb/Vibrio sp. W13 | −/75/−/50 | 30 | [49] |
| Alg7A/Vibrio sp. W13 | −/90/−/70 | 30 | [46] |
| AlgM4/V. weizhoudaoensis M0101 | −/90/−/50 | 30 | [34] |
| AlyS02/Flavobacterium sp. S02 | 90/70/50/40 | 30 | [33] |
| Alyw201/Vibrio sp. W2 | 90/80/50/40 | 30 | [48] |
| Alyw208/Vibrio sp. W2 | 99/92/79/71 | 35 | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.-Y.; Chen, Z.-F.; Zheng, Z.-H.; Lei, H.-W.; Cong, H.-H.; Zhou, H.-X. A Novel Cold-Adapted and High-Alkaline Alginate Lyase with Potential for Alginate Oligosaccharides Preparation. Molecules 2023, 28, 6190. https://doi.org/10.3390/molecules28176190
Wang H-Y, Chen Z-F, Zheng Z-H, Lei H-W, Cong H-H, Zhou H-X. A Novel Cold-Adapted and High-Alkaline Alginate Lyase with Potential for Alginate Oligosaccharides Preparation. Molecules. 2023; 28(17):6190. https://doi.org/10.3390/molecules28176190
Chicago/Turabian StyleWang, Hai-Ying, Zhi-Fang Chen, Zhi-Hong Zheng, Hui-Wen Lei, Hai-Hua Cong, and Hai-Xiang Zhou. 2023. "A Novel Cold-Adapted and High-Alkaline Alginate Lyase with Potential for Alginate Oligosaccharides Preparation" Molecules 28, no. 17: 6190. https://doi.org/10.3390/molecules28176190