1. Introduction
Bisphenol A (BPA) is an extensively used chemical compound, primarily serving as a monomer in the manufacturing of polycarbonate plastics and epoxy resins [
1]. These materials are commonly employed in the production of various everyday items such as food containers, water bottles, milk containers, tin can liners, and numerous household products [
2]. BPA is also present in dental sealants, further expanding its range of applications. This compound, while economically significant, has gained attention as an endocrine-disrupting chemical (EDC) due to its adverse effects on metabolic functions within the human body [
3]. Numerous studies have established a link between BPA exposure and the impairment of several metabolic processes, including the development of conditions such as diabetes mellitus (DM), obesity, and metabolic syndromes [
3,
4,
5,
6,
7,
8]. The disruption of metabolic pathways with BPA can have significant implications for human health [
2]. Studies have indicated that even at low doses, BPA can interfere with carbohydrate metabolism, impair insulin secretion and sensitivity, and contribute to the development of insulin resistance [
9]. Additionally, BPA-induced oxidative stress has been linked to the dysregulation of lipid metabolism, leading to abnormal lipid profiles and increased risk of cardiovascular diseases [
1,
10]. The precise mechanisms through which this occurs have been the subject of scientific inquiry. Additionally, experimental studies have focused on the impact of BPA on the functionality of β-cells and α-cells found in the islets of Langerhans, which are responsible for the secretion of insulin and glucagon, respectively [
11]. These investigations have provided evidence that BPA can interfere with the normal functioning of these cells, potentially leading to imbalances in the regulation of glucose levels and hormone secretion [
2]. Overall, the utilization of BPA in various consumer products raises concerns regarding its potential detrimental effects on metabolic health and endocrine function [
12]. Extensive research efforts continue to delve into the mechanisms and consequences of BPA exposure in order to better understand and address the associated risks to human health. Accumulating evidence suggests that BPA exposure is associated with the development of metabolic disorders, including DM, obesity, and metabolic syndromes [
13].
The detrimental effects of BPA on metabolic functions have been extensively investigated in both in vitro and in vivo studies [
14]. Experimental evidence has revealed that BPA can disrupt the functionality of pancreatic β-cells and α-cells, which are responsible for insulin and glucagon secretion, respectively [
11]. The dysregulation of these cells can lead to impaired glucose metabolism and insulin resistance, contributing to the development of DM and related metabolic disorders [
15]. Furthermore, BPA-induced oxidative stress plays a critical role in the pathogenesis of metabolic disorders [
16]. Lipid peroxidation, resulting from the oxidation of lipoproteins and lipids, can overwhelm the antioxidant defense system, induce cellular apoptosis, and elevate the production of ROS [
17]. Elevated ROS levels can further exacerbate insulin resistance, impair pancreatic β-cell function, and promote inflammation and tissue damage [
18]. Given the alarming rise in BPA exposure and its potential health risks, there is a need to explore therapeutic interventions that can counteract the disruptions caused by BPA on metabolic pathways [
19].
Maintaining a delicate balance between the oxidation and reduction of chemicals is crucial for various aspects of cellular development, growth, and survival [
20]. Reactive oxygen species (ROS), including peroxides, O
2−, and OH radicals, are naturally produced as byproducts of cellular metabolism [
21,
22]. Cells possess tightly regulated pathways to utilize small amounts of these radicals for gene regulation while effectively clearing excessive ROS to prevent detrimental effects [
22]. Disruptions in this balance can render cells insensitive to cell death signals triggered by environmental toxins, leading to genetic mutations and alterations in gene expression [
23,
24]. Thus, the intricate management of ROS levels is essential for maintaining cellular health and preventing oxidative-stress-related damage [
25].
Resveratrol (RSV), a member of the polyphenol class, possesses antioxidant properties and plays a role in reducing cellular oxidative stress [
26]. RSV has emerged as a potential candidate due to its ability to modulate gene expressions related to glucose metabolism, enhance insulin sensitivity, and mitigate oxidative stress [
26]. This compound can be found naturally in various sources such as blueberries, mulberries, raspberries, and peanuts, and is particularly abundant in the peel and seeds of red grapes [
27]. The beneficial properties of RSV are wide-ranging and include antioxidant, antidiabetic, antiadipogenic, anti-inflammatory, antitumor, and antimicrobial activities [
28]. RSV exists in two interconvertible isomeric forms [
29], with trans-resveratrol exhibiting greater stability compared to cis-resveratrol [
30,
31,
32]. Previous research has demonstrated that RSV can improve glucose homeostasis, enhance insulin sensitivity, and ameliorate oxidative stress in various experimental models [
33]. These beneficial effects of RSV make it a promising candidate for mitigating the adverse effects induced by BPA exposure.
In this study, we aimed to investigate the disruptions of metabolic pathways induced by BPA and explore the potential therapeutic intervention provided by RSV. We assessed the effects of BPA exposure on various biomarkers related to glucose metabolism, lipid metabolism, oxidative stress, and inflammation. Additionally, we examined the gene expressions of key biomarkers involved in glucose metabolism and insulin regulation. By examining the effects of resveratrol on BPA-induced metabolic dysfunction, we aimed to uncover its ability to counteract the detrimental effects of BPA and potentially provide a therapeutic intervention for individuals exposed to BPA-related metabolic disturbances.
3. Materials and Methods
3.1. Chemicals and Assay Kits
Insulin ELISA kit (catalogue number: IS130D, Calbiotech, Inc., El Cajon, CA, USA), HbA1c ELISA kit (catalogue number: SG 10984, Elabsciences, Wuhan, China), non-esterified free fatty acid ELISA kit (catalogue number: E-BC-K013, Elabsciences), HDL ELISA kit (catalogue number: E-BC-K222-S, Elabsciences), cholesterol ELISA kit (catalogue number: BD090618, Human diagnostics, Ahrensburg, German), triglyceride ELISA kit (catalogue number: BD090618, bioactive, Bangkok, Thailand.), HMG-CoA reductase ELISA kit (catalogue number: E-EL-H2472 Elabsciences), leptin ELISA kit (catalogue number: 201905, Elabsciences), adiponectin ELISA kit (catalogue number: E-BC-K013-S, Elabsciences), DPP-4 ELISA kit (catalogue number: ab133081, Abcam, Waltham, MA, USA), catalases ELISA kit (catalogue number: E-BC-K106, Elabsciences), glutathione peroxidae ELISA kit (catalogue number: E-EL-R2491, Elabsciences), superoxide dismutase ELISA kit (catalogue number: E-BC-K020, Elabsciences), malondialdehyde ELISA kit (catalogue number: E-EL-0060, Elabsciences), α-Amylase ELISA kit (catalogue number: E-EL-R2544, Elabsciences), α-glucosidase ELISA kit (catalogue number: E-EL-R1083, Elabsciences), hexokinase ELISA kit (catalogue number: E-EL-RR0502, Elabsciences), glucose 6 phosphatase ELISA kit (catalogue number: E-EL-M1362, Elabsciences), GLUT-4 ELISA kit (catalogue number: E-EL-RR0430, Elabsciences), TNF-α ELISA kit (catalogue number: E-EL-R0019, Elabsciences), and IL-6 ELISA kit (catalogue number: E-EL-R0015, Elabsciences).
3.2. Experimental Design
For this study, a total of thirty adult Wistar male albino rats, weighing between 150 and 200 g, were carefully selected. These rats were housed in the animal facility of the university, providing a suitable environment for their well-being. The rats were accommodated in polycarbonate cages with stainless steel covers and provided with woodchip bedding, which offered comfort and a suitable surface for movement. The study was conducted for four weeks.
The animal facility maintained optimal conditions to ensure the rats’ health and minimize any external factors that could influence the experiment’s outcomes. The temperature within the facility was kept constant at 25 ± 5 °C, creating a comfortable and stable environment. The relative humidity was maintained at 50 ± 10%, providing adequate moisture levels for the rats’ well-being. Appropriate light and dark cycles were implemented to simulate natural day and night conditions, promoting regular circadian rhythms.
To meet their nutritional needs, the rats were provided with standard pellet food and water ad libitum, meaning they had unrestricted access to food and water throughout the study. The food provided met the required nutritional standards for laboratory rats, ensuring a balanced diet. The thirty rats selected for the experiment were divided into three groups as follows:
(1) Control group: This group received distilled water, serving as the baseline reference for comparison.
(2) Group 2: This group received BPA at a dosage of 70 mg/kg/day.
(3) This group received a combination of BPA at a dosage of 70 mg/kg/day and RSV at a dosage of 100 mg/kg/day.
Preparation of stock solution of BPA: BPA is available in a small white pellet form. Its stock solution was prepared by following the method as described previously [
34].
Preparation of stock solution of RSV: RSV is available in the form of a coarse powder and its solubility in water is limited but is more soluble in organic solutions. For the administration of the RSV into the experimental animals, RSV was blended in corn oil and the concentration of RSV was adjusted in the corn oil according to the body weight of the experimental animals by following the method as described previously [
34] with some modifications.
Optimum selection of RSV dose: The dose of RSV (100 mg/kg/day) was selected on the basis of the beneficial effects of the RSV as described previously [
35,
36]. Moreover, in our study, we also observed that when we administered RSV blended in corn oil to the normal rats, we did not observe any alterations in the metabolomic profiling of the normal rats [
34]. Therefore, we did not choose a separate experimental rat group to investigate the any possible toxicity and/or effects of RSV on the normal metabolic processes of healthy rats.
The administration of BPA and/or RSV to the rats continued for a period of four weeks. Throughout the study, the rats’ food and water intake were closely monitored on a daily basis. This monitoring aimed to ensure that the rats consumed the administered substances and maintained their usual dietary patterns. By tracking their food and water intake, any potential variations or effects on the rats’ overall well-being could be observed and considered during the analysis of the experiment’s results.
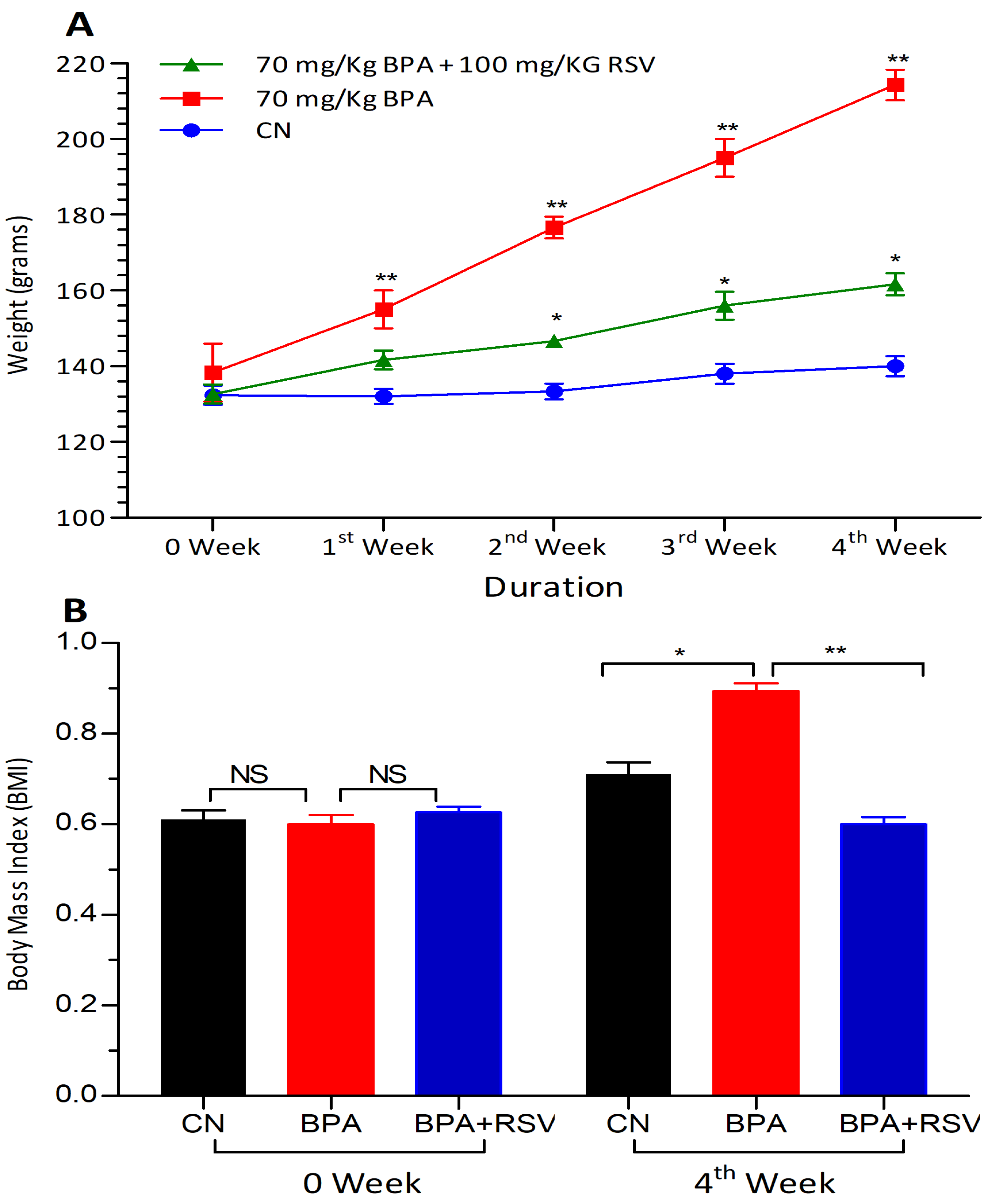
3.3. Evaluation of Anthropometric Markers
In this study, anthropometric markers were used to assess the body weight and body mass index (BMI) of the experimental rats. The body weight of each rat was measured at the beginning of the experiment (week 0) and then monitored weekly throughout the study period. This allowed us to track any changes in body weight over time and evaluate the impact of the experimental conditions on the rats’ growth and development. The formula used to calculate the BMI of rats in this study was as follows:
By plugging in the recorded body weight and body length values into this equation, we were able to obtain the BMI values for each rat at the start and end of the experiment. By monitoring both the body weight and BMI of the rats throughout the study, we gained important insights into the physical changes and growth patterns of the experimental animals. These anthropometric markers served as valuable indicators of the rats’ response to the experimental conditions and provided quantitative data for evaluating the effects of the study interventions on their body composition.
3.4. Biochemical Analysis
3.4.1. Evaluation of Glycemic Control Biomarkers
In this study, we aimed to evaluate glycemic control by assessing several important biomarkers, including serum insulin level (measured in mU/L), plasma glucose level (measured in mmol/L), HbA1c (measured in mmol/mol), and HOMA-IR.
To measure rat blood glucose levels, we employed the glucose strip method. The estimation of serum insulin, HbA1c, and DPP-4 levels in rats was conducted using ELISA kits in accordance with the guidelines provided by the manufacturers. To quantify the optical density (OD) measurements, a microplate ELISA (Bio Tek instrument, Inc., Winooski, VT, USA) was employed. Additionally, to calculate the HOMA-IR, we utilized fasting values of glucose and insulin. HOMA-IR is a widely used index for assessing insulin resistance. It is calculated using the following formula:
By plugging in the fasting values of glucose (measured in nmol/L) and insulin (measured in µU/mL) into this formula, we were able to obtain a numerical value representing the degree of insulin resistance. Overall, through the combination of these measurement methods and calculations, we gained valuable insights into the glycemic control and insulin sensitivity of the study participants, providing a comprehensive understanding of their metabolic health.
3.4.2. Evaluation of Lipid Profile Biomarkers
In this study, the serum levels of important lipid profile biomarkers, including free fatty acids (FFAs), high-density lipoprotein (HDL), and triglycerides (TGs), were meticulously measured to gain insights into lipid metabolism. To assess these biomarkers accurately, calorimetric assay kits specifically designed for quantifying FFAs, HDL, and TGs in serum samples were used.
To perform the analysis, we employed the Micro-lab 300 chemistry analyzer. By using the calorimetric assay kits in conjunction with the Micro-lab 300 chemistry analyzer, we were able to obtain precise measurements of FFAs, HDL, and TGs in the serum samples. These measurements provided valuable information about the lipid profile of the study participants, enabling a deeper understanding of lipid metabolism and its potential implications in various health conditions, such as cardiovascular diseases and metabolic disorders.
3.4.3. Evaluation of Cholesterol Biosynthesis Biomarker
In this study, the hepatic cholesterol biosynthesis biomarker, HMG-CoA reductase, was carefully assessed from the serum to understand its activity. We employed the ELISA kit assay method to evaluate the serum levels of HMG-CoA reductase.
3.4.4. Evaluation of Biomarkers of Carbohydrate Metabolism
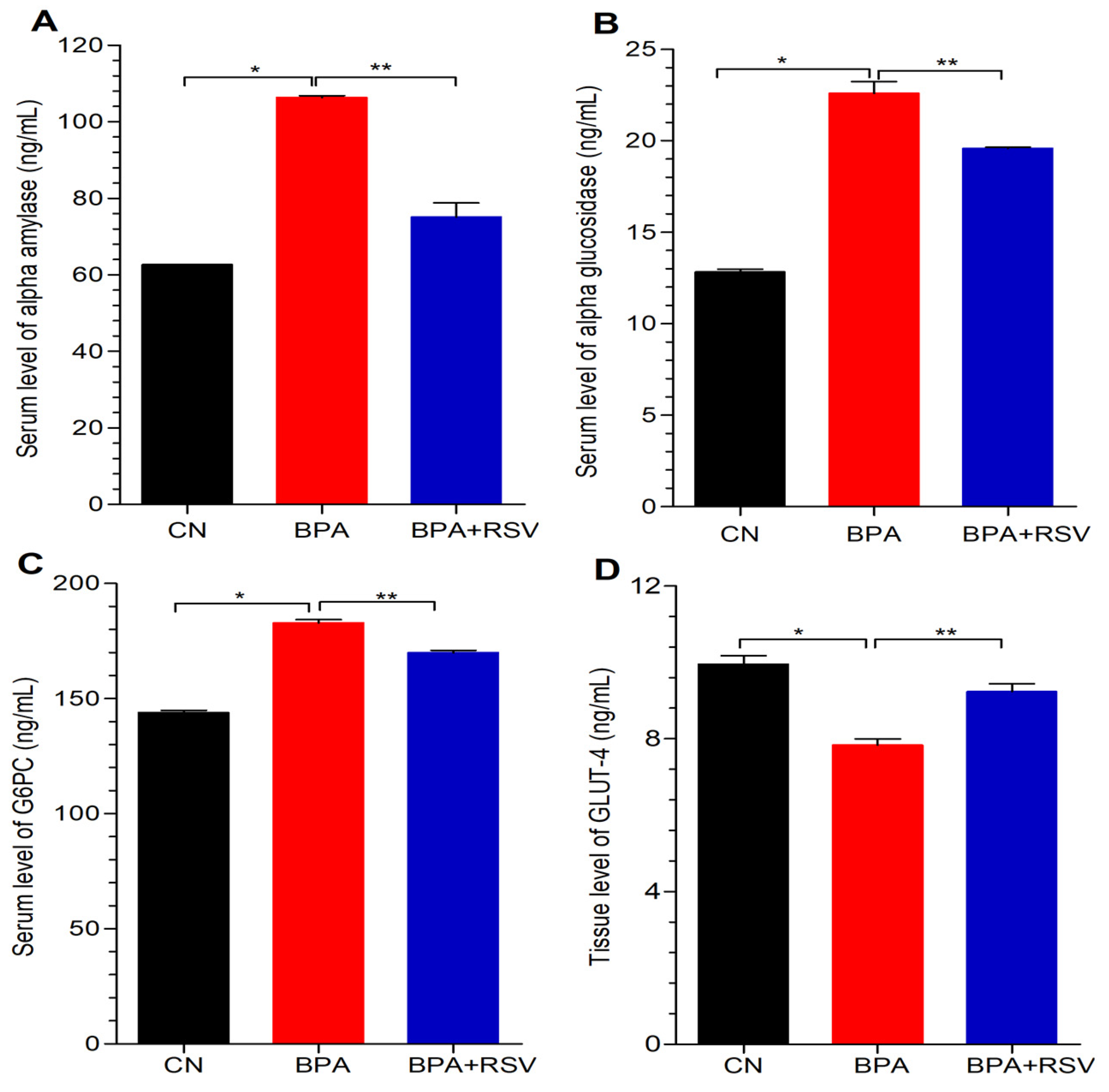
We also evaluated the serum levels of vital biomarkers directly associated with carbohydrate metabolism. By focusing on the key enzymes involved in this metabolic pathway, namely α-amylase, α-glucosidase, hexokinase, and glucose-6-phosphatase, we sought to unravel valuable insights into the intricate workings of carbohydrate utilization within the body. To accomplish this, we employed the ELISA method to accurately quantify the serum levels of these biomarkers, which allowed us to determine the OD at a specific wavelength of 450 nm. By examining the serum levels of α-amylase, α-glucosidase, hexokinase, and glucose-6-phosphatase, we were able to gain comprehensive insights into the activity and levels of these crucial enzymes involved in carbohydrate metabolism.
3.4.5. Evaluation of Inflammatory Biomarkers
In addition to the extensive assessments mentioned above, we further investigated the serum levels of crucial inflammatory biomarkers including TNF-α (Tumor Necrosis Factor-alpha), IL-6 (Interleukin-6), adiponectin, and leptin to gain a comprehensive understanding of the inflammatory status and potential metabolic implications in the study subjects. To quantify the serum levels of these inflammatory biomarkers, we employed the widely employed ELISA method. To capture the measurements, we utilized a microplate ELISA reader and recorded the OD readings at a specific wavelength of 450 nm. By measuring the levels of these inflammatory biomarkers, we aimed to gain valuable insights into the inflammatory status of the study subjects.
3.5. Preparation of Tissue Homogenates
At the end of the treatment period, the muscle tissues were collected for further analysis. To ensure the preservation of tissue integrity, the muscle samples were promptly isolated and placed in an icebox. This step was crucial in preventing degradation and maintaining the quality of the samples.
To prepare tissue homogenates for subsequent analysis, the muscle samples were homogenized using a tissue homogenizer. The homogenization process involved combining the muscle samples with 0.01 M phosphate buffer at a ratio of 1 g of tissue per 9 mL of buffer. The purpose of this step was to break down the muscle tissues and create a uniform suspension of cellular components in the buffer solution.
Following the homogenization step, the resulting homogenates were incubated on ice for 10 min. This incubation period was necessary to ensure proper sample stabilization and allow any enzymatic reactions to subside. Cooling the homogenates on ice helped to maintain the integrity of the cellular components and prevent further enzymatic activity that could affect the subsequent analysis. After the incubation period, the homogenates underwent centrifugation at 14,000× g for 10 min. This centrifugation step aimed to separate the cellular debris from the clear supernatants containing the desired components for analysis. By subjecting the homogenates to high centrifugal forces, the cellular debris settled at the bottom of the tubes, allowing for the extraction of the clear supernatants. Carefully extracting the clear supernatants, these samples were then utilized for performing ELISA tests. To ensure accurate and reliable results, the ELISA tests were performed following the instructions provided by the manufacturers of the ELISA kits.
3.5.1. Evaluation of Insulin-Regulated Glucose Transporter
In this significant study, we aimed to assess the levels of GLUT-4 (Glucose Transporter 4) in skeletal muscle tissue homogenates using the ELISA method. By quantifying the levels of GLUT-4 in skeletal muscle tissue, we aimed to gain valuable insights into its role in carbohydrate metabolism.
3.5.2. Evaluation of Oxidative Stress and Lipid Peroxidation
The levels of oxidative stress biomarkers were meticulously assessed in liver tissue homogenates to gain insights into the extent of oxidative stress (catalase, glutathione, and superoxide dismutase) and lipid peroxidation (MDA) using the ELISA kit assay method. By evaluating these biomarkers collectively, we aimed to obtain a comprehensive assessment of oxidative stress and lipid peroxidation levels in the liver tissue homogenates, shedding light on the potential impact of oxidative stress on cellular health and function.
3.6. Evaluation of the mRNA Expression of the Transcriptional Regulation of Glucose Metabolism and Insulin Secretion
To obtain the RNA from the pancreas samples, we utilized TRIzol reagent (Biobasic BS410A-MA18DR0J) following the TRIzol method. This widely used method enabled the efficient extraction of total RNA from the tissue samples, ensuring its integrity and preserving the RNA’s quality for downstream analysis. To assess the quality and integrity of the extracted RNA, we performed gel electrophoresis using a 2% agarose gel. For the reverse-transcription process, 2 μg of the extracted total RNA was converted into single-stranded complementary DNA (cDNA) using the Revert Aid cDNA synthesis kit (Thermo Scientific, USA). To quantify gene expression, real-time polymerase chain reaction (qPCR) was performed. The qPCR thermal cycling was conducted under the following conditions: an initial denaturation step at 95 °C for 5 min, followed by 40 cycles of denaturation at 95 °C for 15 s, annealing at 60 °C for 20 s, and extension at 72 °C for 20 s. These cycling conditions ensured the amplification of specific DNA sequences and enabled accurate quantification of gene expression levels. We employed a Bio-Rad real-time PCR machine to perform the real-time PCR analysis.
In
Table 1, we present the sequence and size information of the expected PCR products for the reference and target genes (GCK, UCP2, IGF-1, and GLUT2). These genes were carefully selected based on their relevance to the study and their potential involvement in pancreatic function and metabolism. Assessing the expression levels of these genes provided valuable insights into their potential role in the pancreatic function and metabolic processes under investigation.
These genes are known to play significant roles in pancreatic function, glucose metabolism, and insulin regulation. To ensure the accurate normalization of mRNA and cDNA quantities, the GADPH (Glyceraldehyde-3-Phosphate Dehydrogenase) gene was chosen as the internal reference gene or housekeeping gene. The GADPH gene is commonly used as a reference gene due to its stable expression across different tissues and experimental conditions.
To calculate ΔCT, the average CT value obtained from triplicate measurements of both the target gene and the reference gene (GADPH) was utilized. The difference between the ΔCT values of the control (baseline) and the sample (experimental condition) was then calculated to determine ΔΔCT. This parameter provides an estimate of the variation in gene expression levels between the control and sample. The fold changes in mRNA expression were calculated using the 2^(−ΔΔCT) method, which allows for the quantification of relative changes in gene expression. The equation used was:
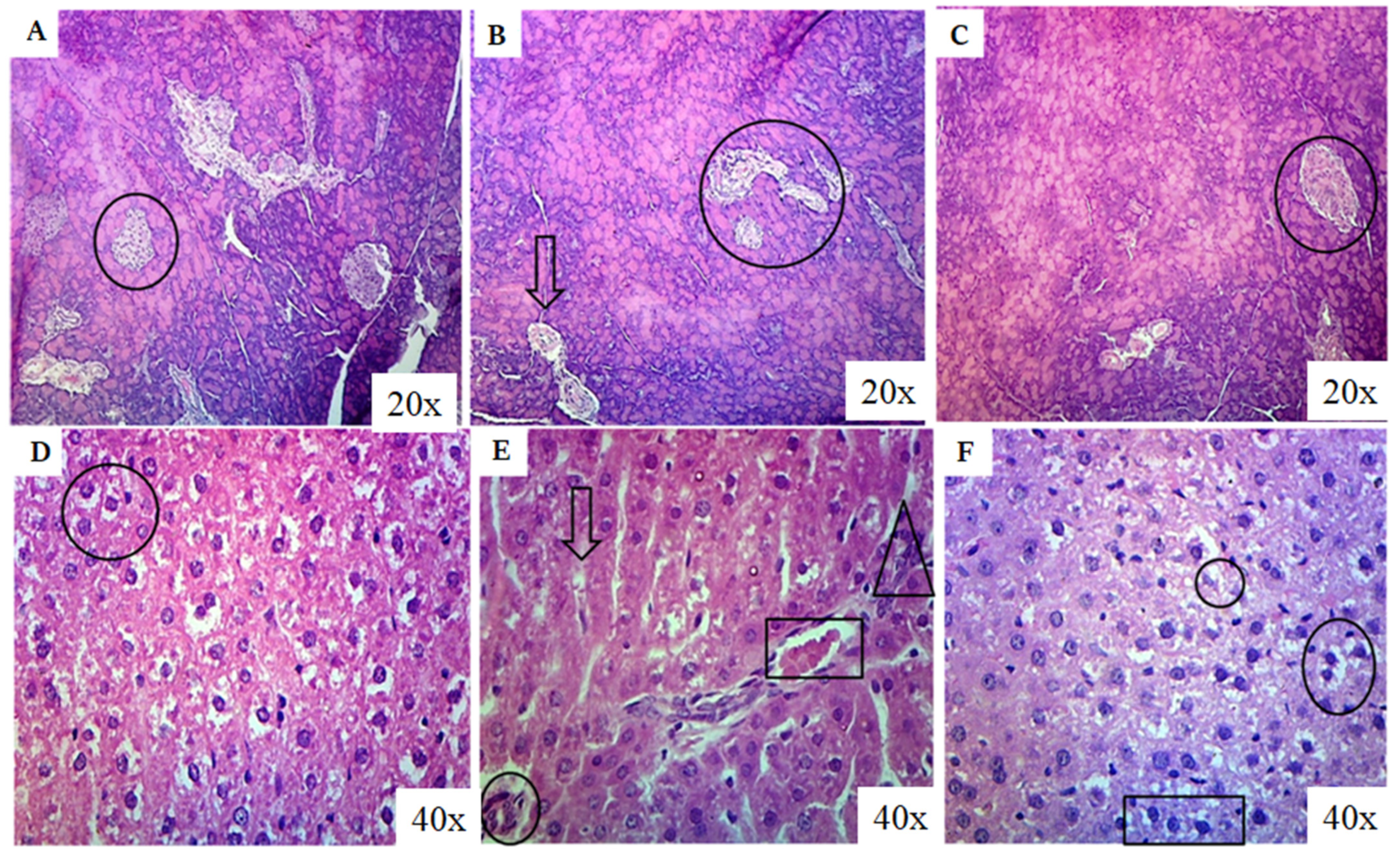
3.7. Histopathological Evaluation
After the completion of the 4th week of the experimental period, the animals were sacrificed under anesthesia. Pancreas were isolated and preserved for biochemical and histopathological analysis. For histopathological analysis, a 10% formalin solution was prepared, and tissues were preserved in this solution. The tissues were washed with distilled water for 6 h, dehydrated using 70% and 90% alcohol for 120 min, and absolute alcohol overnight. After being treated with xylene twice for 80 min, the tissues were dipped into paraffin wax overnight. Tissue blocks were prepared by placing these tissues in paraffin wax into metal plates of cube shape. Using a microtome, tissue ribbons with a thickness of 5 µm were sectioned, and the 5th ribbon section was collected and placed in a water bath at 40 °C. These sections were placed on clean slides for hematoxylin staining. They were then treated with xylene I (5 min) and xylene II (2 min) to remove the paraffin wax. The tissues were hydrated by treating them with 95% ethanol for 2 min, 70% ethanol for 3 min, and 50% ethanol for 5 min. After washing with tap water, they were treated with a sodium bicarbonate bluing solution. These sections were counterstained with eosin dye and dehydrated by increasing concentrations of ethanol (50%, 70%, and 95%). Finally, the slides were cleaned with xylene and observed under a microscope.
3.8. Statistical Analysis
The biochemical parameters were assessed, and their values were determined and reported as the mean ± standard deviation (SD) and GraphPad Prism Software, version: 5 (GraphPad Software Inc., San Diego, CA, USA) was used to analyze the statistical significance of the results. A significance level of p < 0.05 was established to determine the statistical significance of the findings. To further examine the differences between the groups, the post hoc Bonferroni test was employed for both one-way and two-way comparison analysis.
4. Discussion
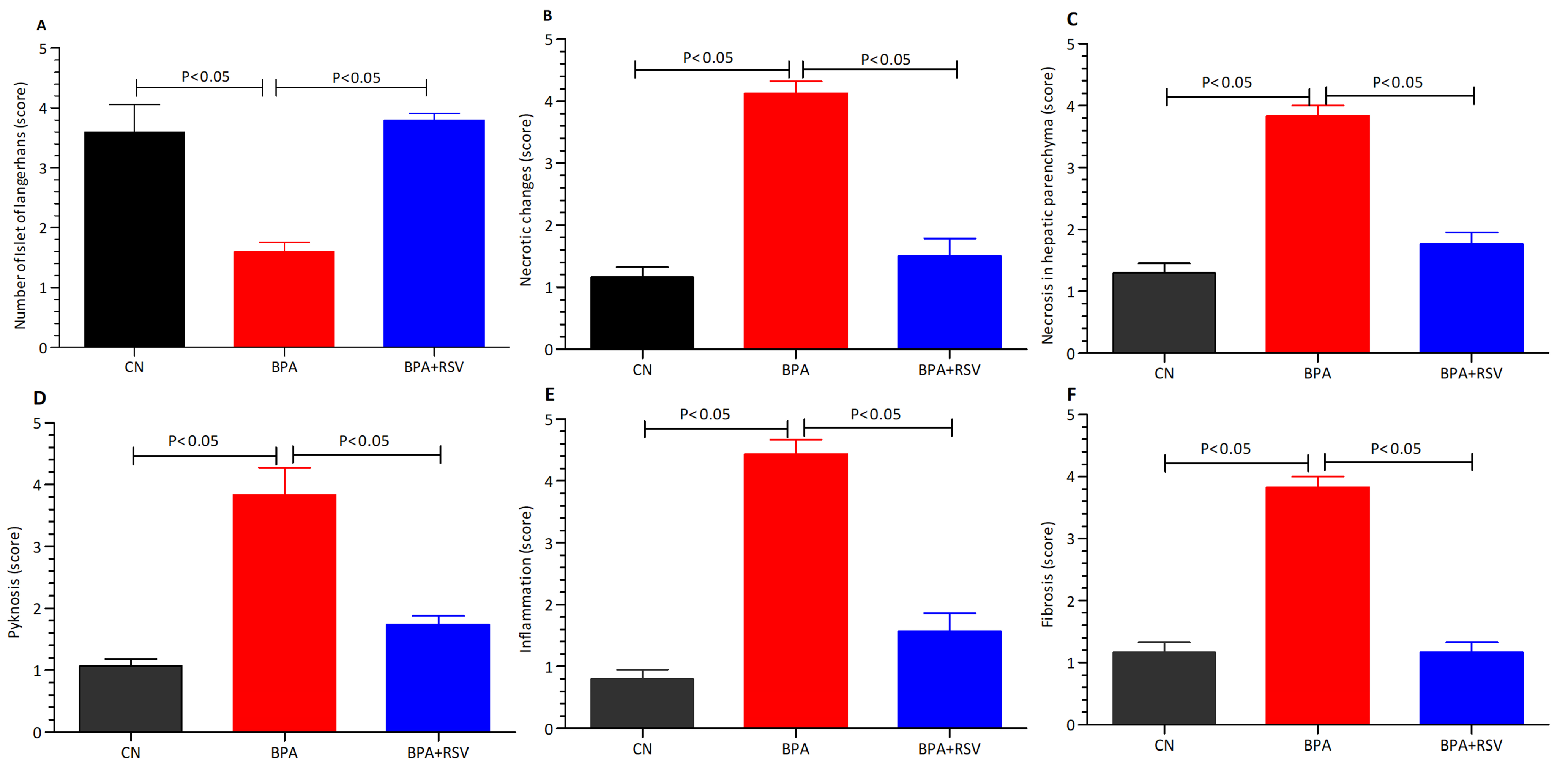
BPA is a chemical compound commonly used in the production of plastics and epoxy resins. It has been a topic of concern due to its potential adverse effects on human health. Numerous studies have investigated the role of BPA in metabolic impairment, particularly in relation to obesity and insulin resistance.
Figure 11 provides a concise summary of the intricate interactions between BPA and various metabolic and signaling pathways, ultimately culminating in the development of diverse metabolic disorders. The diagram visually represents the complex relationship between BPA exposure and its impact on metabolic parameters.
Mitochondrial UCP2 is a crucial component within the β-cells of pancreatic islets. Numerous studies have established a notable correlation between the overexpression of UCP2 and hyperinsulinemia, highlighting its involvement in the development of metabolic disorders [
37,
38]. As a result, the regulation of UCP2 levels becomes of utmost importance, as disturbances in its expression can disrupt essential metabolic processes. Another key element in maintaining glucose homeostasis is the insulin-responsive glucose transporter known as GLUT4. Primarily expressed in skeletal muscles, GLUT4 plays a vital role in facilitating the uptake of glucose into cells. Dysregulation and impairment of GLUT4 function contribute significantly to the development of insulin resistance [
39,
40]. Interestingly, while GLUT2 is predominantly identified in hepatocytes, it is also present in the pancreas, where it plays a crucial role in glucose-stimulated insulin secretion. GLUT2 enables the diffusion of glucose across cell membranes, initiating intracellular glucose metabolism through the action of the enzyme GCK. Previous findings have demonstrated a strong association between the reduced expression of GLUT2 and impaired glucose-stimulated insulin secretion in diabetic rodents [
41]. IGF-1, a polypeptide hormone composed of 70 amino acids, plays a significant role in glucose metabolism and homeostasis. The downregulation of IGF-1 levels has been shown to have hyperglycemic effects, further emphasizing its importance in maintaining proper glucose regulation [
42].
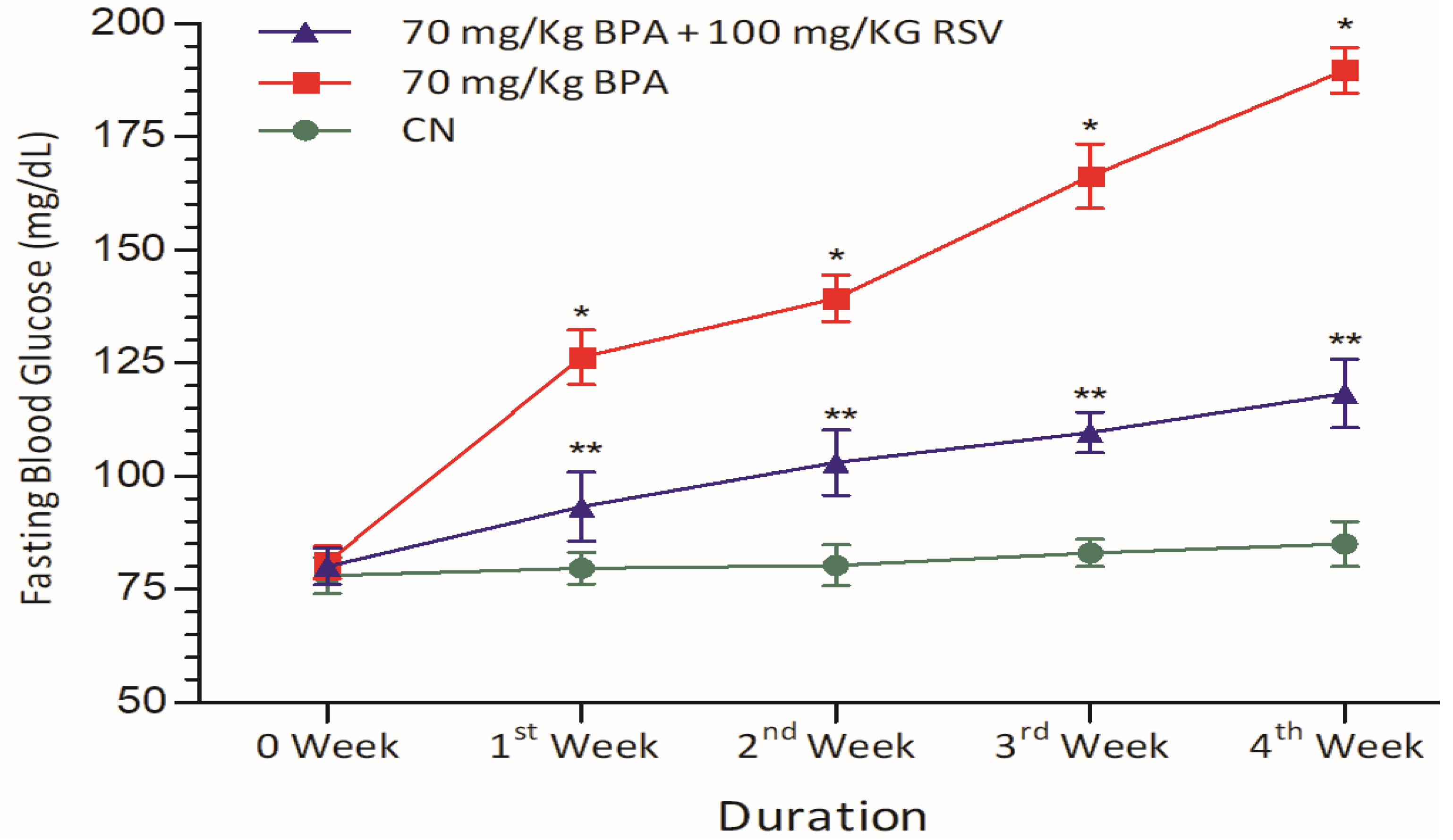
In our study, we observed that BPA exposure significantly elevated blood glucose levels (
Figure 2). This finding is further supported by the increased serum levels of HbA1c, a marker of long-term blood glucose control (
Figure 3B), and HOMA-IR, an index of insulin resistance (
Figure 3C). These results highlight the disruptive effects of BPA on glucose homeostasis and insulin sensitivity, contributing to the development of metabolic disorders. To further support the potential of RSV as a therapeutic intervention, we refer to a study conducted by Liu et al. [
43] that investigated the impact of RSV on blood sugar levels and insulin sensitivity. Their findings demonstrated that RSV exhibited notable improvements in glucose homeostasis and insulin sensitivity. These findings align with our observations and provide additional evidence for the beneficial effects of RSV in managing metabolic dysfunctions.
Collectively, the results from our study and the referenced study emphasize the detrimental effects of BPA on metabolic parameters and highlight the potential of RSV as a therapeutic agent for mitigating the disturbances caused by BPA. By improving glucose regulation and enhancing insulin sensitivity, RSV holds promise as a beneficial intervention in managing metabolic dysfunctions induced by BPA exposure. These findings provide valuable insights into the potential application of RSV as a protective and therapeutic strategy in the context of BPA-related metabolic disorders. Lipid peroxidation is a process resulting from the oxidation of lipoproteins and lipids, which can lead to cellular apoptosis and an increase in free radicals. Serum levels of MDA are often used as an indicator of lipid peroxidation and oxidative stress [
44]. In our study, we found that BPA exposure led to elevated MDA levels (
Figure 6D), indicating an increase in free radicals.
To further understand the impact of BPA on the liver, we examined the levels of antioxidants. Our findings revealed a significant decrease in CAT levels (
Figure 6A), GPx levels (
Figure 6B), and SOD levels (
Figure 6C) in the BPA-exposed group. These results indicate that BPA stimulates the production of ROS and disrupts the liver’s antioxidant defense system. Previous research has demonstrated the beneficial effects of RSV treatment in reducing serum MDA levels, increasing SOD levels, enhancing GSH and GPx levels, and reducing ROS levels in liver tissues. Consistent with these findings, our study observed similar outcomes in liver tissue homogenates, further validating the potential of RSV in counteracting BPA-induced oxidative stress. In addition to oxidative stress, we investigated the expression of key genes involved in glucose metabolism and insulin regulation. GCK, a crucial enzyme involved in glucose metabolism and insulin release, plays a significant role in maintaining glucose homeostasis. We observed that BPA suppressed the expression of GCK, while RSV treatment improved its expression (
Figure 8).
IGF-1, another factor that stimulates insulin release, was also impacted by BPA exposure. We found a significant suppression of IGF-1 expression in response to BPA exposure, while RSV treatment increased its mRNA expression (
Figure 8), consistent with previous studies. These findings suggest that BPA disrupts insulin regulation and glucose homeostasis, while RSV treatment shows potential in restoring their balance. UCP2, a critical player in lipid and energy metabolism, is associated with obesity and hyperinsulinemia. In our study, we observed an increase in UCP2 expression following BPA exposure (
Figure 9), aligning with previous research. This upregulation of UCP2 may contribute to the disruption of mitochondrial acetyl-CoA oxidation and energy metabolism.
Furthermore, our study also observed the downregulation of GCK and GLUT2 expression in the pancreas of BPA-treated groups, which can disrupt normal glucose metabolism. However, RSV treatment showed potential in restoring their activity, as seen by the augmentation of GCK and GLUT2 expression. These findings provide insights into the impact of BPA on key genes involved in carbohydrate metabolism and insulin regulation and highlight the potential of RSV in restoring their function.
Overall, our study demonstrates the detrimental effects of BPA-induced oxidative stress on liver tissue and the potential therapeutic role of RSV in counteracting these effects by effectively neutralizing free radicals. Moreover, the dysregulation of key genes involved in carbohydrate metabolism and insulin regulation further emphasizes the impact of BPA on metabolic pathways, while highlighting the potential of RSV in restoring their normal activity. RSV, through its various mechanisms of action, exerts effects on glucose homeostasis by upregulating the expression of GLUT2 and GCK genes. This regulation is mediated through the activation of SIRT1 genes, which play a crucial role in maintaining glucose balance and positively regulating glucose-stimulated insulin secretion in pancreatic β-cells. The activation of SIRT1 by RSV can increase insulin secretion and ATP production by suppressing the transcriptional activity of the UCP2 gene. Previous studies have also suggested that the RSV-mediated downregulation of UCP2 occurs through SIRT1-dependent activation. Therefore, RSV activates SIRT1, which in turn represses UCP2 expression.
In our study, we observed that RSV treatment resulted in an increase in the expression of GLUT2 and GCK genes, which are important for glucose metabolism and insulin regulation. These findings indicate that RSV has the potential to enhance glucose uptake and utilization, leading to improved glucose homeostasis. Furthermore, RSV treatment led to a decrease in the expression of UCP2, which is associated with improved mitochondrial function and energy metabolism. These findings support the notion that RSV can modulate gene expression related to glucose metabolism and contribute to the downregulation of UCP2 expression. The regulation of these key genes by RSV provides insights into the molecular mechanisms underlying its beneficial effects on metabolic disorders induced by BPA exposure.
However, it is important to consider the limitations of our study. While our findings provide valuable insights, they should be interpreted within the context of the study’s limitations. Further research is needed to validate these results and expand our understanding of the underlying mechanisms. Future studies should involve human subjects to confirm the translatability of the findings. Additionally, elucidating the specific signaling pathways and molecular mechanisms through which RSV exerts its effects would provide a more comprehensive understanding of its therapeutic potential. Long-term studies are also necessary to assess the sustained effects of RSV treatment. Exploring additional pathways and optimizing the dosage and duration of RSV treatment are important for maximizing its therapeutic benefits.
5. Conclusions
The data presented in this study highlight the detrimental effects of BPA exposure on metabolic homeostasis and provide evidence for the potential of RSV as a therapeutic intervention to mitigate these disruptions. The findings demonstrate that BPA induces inflammation, impairs carbohydrate metabolism, causes histopathological changes in pancreatic and liver tissues, induces oxidative stress, and disrupts gene expression related to glucose and lipid metabolism. However, RSV treatment shows promise in counteracting these adverse effects. RSV effectively reduces inflammation by restoring the balance of inflammatory adipocytokines. It improves carbohydrate metabolism by upregulating the expression of GLUT2 and GCK genes, crucial for glucose homeostasis. RSV also ameliorates the histopathological changes in the pancreas and liver, indicating its protective effects on these tissues. Furthermore, RSV exhibits potent antioxidant properties, scavenging free radicals and restoring the activity of key antioxidant enzymes. The modulation of gene expression by RSV is another significant finding. It upregulates GLUT2 and GCK genes while downregulating UCP2, suggesting its beneficial effects on glucose and lipid metabolism. These results collectively highlight the potential of RSV as a therapeutic agent in managing BPA-induced metabolic disorders. However, it is important to acknowledge the limitations of the current study, including the need for further research involving human subjects, the elucidation of underlying mechanisms, long-term studies, the exploration of additional pathways, and the optimization of dosage and duration. Overall, the findings underscore the importance of addressing the adverse effects of BPA on metabolic pathways and provide valuable insights into the potential of RSV as a promising intervention to restore metabolic homeostasis. Further research in this field will contribute to a better understanding of the therapeutic mechanisms and optimal utilization of RSV to counteract the detrimental effects of BPA on metabolism.


 ,
, 












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}