
Isolation, Structural Characteristics Analysis of a Vigna unguiculata Polysaccharide VUP80-3 and Its Protective Effect on GES-1 Cells In Vitro
Abstract
:1. Introduction
2. Results
2.1. Preparation of V. unguiculata Polysaccharides
2.2. Molecular Weight of VUP80-3
2.3. FTIR Spectra
2.4. Monosaccharide Analysis of VUP80-3
2.5. Methylation Analysis Results
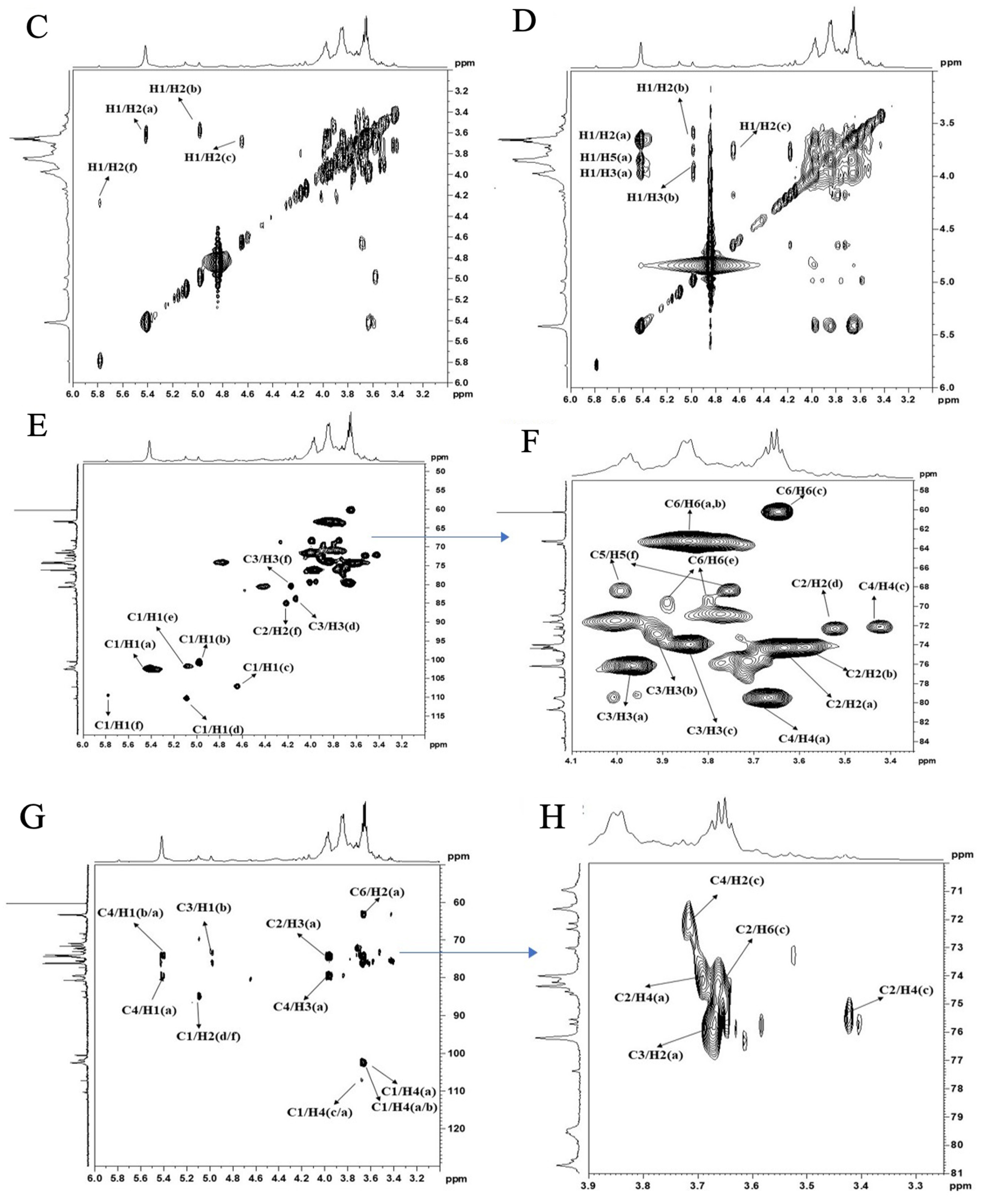
2.6. NMR Spectroscopy Analysis
2.7. SEM Analysis
2.8. Effect of VUP80-3 on the Activity of Ethanol-Induced GES-1 Damaged Cells
2.8.1. Establishment of GES-1 Cell Injury Model
2.8.2. Effect of Polysaccharide VUP80-3 on ROS in Injured GES-1 Cells
2.8.3. Effect of VUP80-3 on the Secretion of Inflammatory Factors by Injured GES-1 Cells
2.8.4. Effect of Polysaccharide VUP80-3 on Apoptosis of Injured GES-1 Cells
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Preparation of V. unguiculata Polysaccharide
4.3. Molecular Weight Analysis
4.4. Infrared Spectrum Analysis
4.5. Monosaccharide Composition Analysis
4.6. Methylation Analysis
4.7. NMR Spectroscopy Analysis
4.8. Scanning Electron Microscopy (SEM) Analysis
4.9. Protection of VUP80-3 against Ethanol-Induced GES-1 Injury
4.9.1. Cell Culture and Establishment of GES-1 Cell Injury Model
4.9.2. Analysis of Intracellular ROS and Inflammatory Factors
4.9.3. Apoptosis Analysis by Flow Cytometry
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Czekaj, R.; Majka, J.; Magierowska, K.; Sliwowski, Z.; Magierowski, M.; Pajdo, R.; Brzozowski, T. Mechanisms of curcumin-induced gastroprotection against ethanol-induced gastric mucosal lesions. J. Gastroenterol. 2017, 53, 618–630. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.; Chen, T.; Zhao, Y.; Lu, S.; Yu, H.; Chang, Y.; Chen, D. Study of the mechanism of anti-ulcer effects of virgin coconut oil on gastric ulcer-induced rat model. Arch. Med. Sci. 2019, 15, 1329–1335. [Google Scholar] [CrossRef]
- Chang, W.; Bai, J.; Tian, S.; Ma, M.; Li, W.; Yin, Y.; Deng, R.; Cui, J.; Li, J.; Wang, G.; et al. Autophagy protects gastric mucosal epithelial cells from ethanol-induced oxidative damage via mTOR signaling pathway. Exp. Biol. Med. 2017, 242, 1025–1033. [Google Scholar] [CrossRef] [Green Version]
- Gong, H.; Li, W.; Sun, J.; Jia, L.; Guan, Q.; Guo, Y.; Wang, Y. A review on plant polysaccharide based on drug delivery system for construction and application, with emphasis on traditional Chinese medicine polysaccharide. Int. J. Biol. Macromol. 2022, 211, 711–728. [Google Scholar] [CrossRef]
- Shen, C.; Wang, T.; Guo, F.; Sun, K.; Wang, B.; Wang, J.; Zhang, Z.; Zhang, X.; Zhao, Y.; Chen, Y. Structural characterization and intestinal protection activity of polysaccharides from Sea buckthorn (Hippophae rhamnoides L.) berries. Carbohydr. Polym. 2021, 274, 118648. [Google Scholar] [CrossRef]
- Liu, Y.; Sui, D.; Fu, W.; Sun, L.; Li, Y.; Yu, P.; Yu, X.; Zhou, Y.; Xu, H. Protective effects of polysaccharides from Panax ginseng on acute gastric ulcers induced by ethanol in rats. Food Funct. 2021, 12, 2741–2749. [Google Scholar] [CrossRef]
- Ranade, A.N.; Wankhede, S.S.; Ranpise, N.S.; Mundada, M.S. Development of bilayer floating tablet of amoxicillin and Aloe vera gel powder fortreatment of gastric ulcers. AAPS Pharm. Sci. Tech. 2012, 13, 1518–1523. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, S.; Sathish Kumar, D.; Arulselvan, P.; Senthilkumar, G.P.; Mahadeva Rao, U.S. Evaluation of anti-ulcerogenic potential of Aloe vera leaf gel extract studied in experimental rats. J. Pharmacol. Toxicol. 2007, 2, 85–97. [Google Scholar] [CrossRef]
- Metowogo, K.; Eklu-Gadegbeku, K.; Agbonon, A.; Aklikokou, K.A.; Gbeassor, M. Gastroprotective effect of hydroalcoholic extract of Aloe buettneri. Iran. J. Pharm. Res. 2011, 10, 69–74. [Google Scholar]
- Wang, Y.; Su, W.; Zhang, C.; Xue, C.; Chang, Y.; Wu, X.; Tang, Q.; Wang, J. Protective effect of sea cucumber (Acaudina molpadioides) fucoidan against ethanol-induced gastric damage. Food Chem. 2012, 133, 1414–1419. [Google Scholar] [CrossRef]
- Kan, J.; Hood, M.; Burns, C.; Scholten, J.; Chuang, J.; Tian, F.; Pan, X.; Du, J.; Gui, M. A Novel Combination of Wheat Peptides and Fucoidan Attenuates Ethanol-Induced Gastric Mucosal Damage through Anti-Oxidant, Anti-Inflammatory, and Pro-Survival Mechanisms. Nutrients 2017, 9, 978. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Wu, D.; Jin, Y.; Li, Q.; Liu, Y.; Qiao, X.; Zhang, J.; Dong, G.; Li, Z.; Li, T.; et al. Pre-protective effect of polysaccharides purified from Hericium erinaceus against ethanol-induced gastric mucosal injury in rats. Int. J. Biol. Macromol. 2020, 159, 948–956. [Google Scholar] [CrossRef]
- Li, A.L.; Xiong, S.L. Evaluation of amino acid content and nutritional value of cowpea seed protein. Food Res. Dev. 2008, 6, 147–150. (In Chinese) [Google Scholar] [CrossRef]
- Abd-Alla, M.H.; Bashandy, S.R.; Nafady, N.A.; Hassan, A.A. Enhancement of exopolysaccharide production by Stenotrophomonas maltophilia and Brevibacillus parabrevis isolated from root nodules of Cicer arietinum L. and Vigna unguiculata L. (Walp.) plants, Rendiconti lincei. Sci. Fis. E Nat. 2018, 29, 117–129. [Google Scholar] [CrossRef]
- Moloto, M.R.; Phan, A.D.T.; Shai, J.L.; Sultanbawa, Y.; Sivakumar, D. Comparison of Phenolic Compounds, Carotenoids, Amino Acid Composition, In Vitro Antioxidant and Anti-Diabetic Activities in the Leaves of Seven Cowpea (Vigna unguiculata) Cultivars. Foods 2020, 9, 1285. [Google Scholar] [CrossRef]
- Rengadu, D.; Gerrano, A.S.; Mellem, J.J. Physicochemical and structural characterization of resistant starch isolated from Vigna unguiculata. Int. J. Biol. Macromol. 2020, 147, 268–275. [Google Scholar] [CrossRef]
- Xin, Y.; Song, X.; Wang, Y.; Nie, S.; Yin, J. Structural Characteristics and Antioxidant Properties of Polysaccharides from Different Parts of Cowpea. Food Biosci. 2022, 43, 61–67. [Google Scholar] [CrossRef]
- Zhang, B.; Deng, Z.; Ramdath, D.D.; Tang, Y.; Chen, P.X.; Liu, R.; Liu, Q.; Tsao, R. Phenolic profiles of 20 Canadian lentil cultivars and their contribution to antioxidant activity and inhibitory effects on α-glucosidase and pancreatic lipase. Food Chem. 2015, 172, 862–872. [Google Scholar] [CrossRef]
- Wang, Z.; Zhou, X.; Shu, Z.; Zheng, Y.; Hu, X.; Zhang, P.; Huang, H.; Sheng, L.; Zhang, P.; Wang, Q.; et al. Regulation strategy, bioactivity, and physical property of plant and microbial polysaccharides based on molecular weight. Int. J. Biol. Macromol. 2023, 244, 125360. [Google Scholar] [CrossRef] [PubMed]
- Luo, A.; Ge, Z.; Fan, Y.; Luo, A.; Chun, Z.; He, X. In Vitro and In Vivo Antioxidant Activity of a Water-Soluble Polysaccharide from Dendrobium denneanum. Molecules 2011, 16, 1579–1592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Ma, J.; Wang, G.; Li, X.; Liu, Y.; Xu, E.; Luo, A. Ultrasonic extraction, structural modification and gastric mucosal cells protective activity of a polysaccharide from Dendrobium denneanum. Arab. J. Chem. 2023, 16, 105033. [Google Scholar] [CrossRef]
- Fan, Y.; Lin, M.; Luo, A.; Chun, Z.; Luo, A. Characterization and Antitumor Activity of a Polysaccharide from Sarcodia ceylonensis. Molecules 2014, 19, 10863–10876. [Google Scholar] [CrossRef]
- Fan, Y.; He, Q.; Luo, A.; Wang, M.; Luo, A. Characterization and Antihyperglycemic Activity of a Polysaccharide from Dioscorea opposita Thunb Roots. Int. J. Mol. Sci. 2015, 16, 6391–6401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Li, X.; Chen, K.; Yang, H.; Jialengbieke, B.; Hu, X. Extraction optimization, characterization and the antioxidant activities in vitro and in vivo of polysaccharide from Pleurotus ferulae. Int. J. Biol. Macromol. 2020, 160, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Miao, M.; Bai, A.; Jiang, B.; Song, Y.; Cui, S.W.; Zhang, T. Characterisation of a novel water-soluble polysaccharide from leuconostoc citreum SK24.002. Food Hydrocoll. 2014, 36, 265–272. [Google Scholar] [CrossRef]
- Fan, Y.; Lin, M.; Luo, A. Extraction, characterization and antioxidant activities of an acidic polysaccharide from Dendrobium devonianum. J. Food Meas. Charact. 2021, 16, 867–879. [Google Scholar] [CrossRef]
- Sims, I.M.; Carnachan, S.M.; Bell, T.J.; Hinkley, S.F. Methylation analysis of polysaccharides: Technical advice. Carbohydr. Polym. 2018, 188, 1–7. [Google Scholar] [CrossRef]
- Li, J.; Zhong, Y.G.; Liu, C.J. Purification and Electron Microscope Analysis of Lentinan. J. Shanxi Agric. Sci. 2010, 38, 6–9. (In Chinese) [Google Scholar] [CrossRef]
- Li, G.H.; Wang, J.; Gao, X.L.; Guo, W.Y.; Zhang, Y.L.; Wang, Y.H.; Hou, Y.F. Effect of ultrafine grinding of cowpea powder on its physicochemical properties and antioxidant activity. Food Sci. Technol. 2019, 44, 99–103. (In Chinese) [Google Scholar]
- Zeng, Q.; Ko, C.H.; Siu, W.S.; Li, L.F.; Han, X.Q.; Yang, L.; Lau, C.B.S.; Hu, J.M.; Leung, P.C. Polysaccharides of Dendrobium officinale Kimura & Migo protect gastric mucosal cell against oxidative damage-induced apoptosis in vitro and in vivo. J. Ethnopharmacol. 2017, 208, 214–224. [Google Scholar]
- Zhou, D.; Yang, Q.; Tian, T.; Chang, Y.; Li, Y.; Duan, L.R.; Li, H.; Wang, S.W. Gastroprotective effect of gallic acid against ethanol-induced gastric ulcer in rats: Involvement of the Nrf2/HO-1 signaling and anti-apoptosis role. Biomed. Pharmacother. 2020, 126, 110075. [Google Scholar] [CrossRef] [PubMed]
- Beiranvand, M.; Bahramikia, S. Ameliorating and protective effects mesalazine on ethanol-induced gastric ulcers in experimental rats. Eur. J. Pharmacol. 2020, 888, 173573. [Google Scholar] [CrossRef]
- Amirshahrokhi, K.; Khalili, A.-R. The effect of thalidomide on ethanol-induced gastric mucosal damage in mice: Involvement of inflammatory cytokines and nitric oxide. Chem. Interact. 2015, 225, 63–69. [Google Scholar] [CrossRef]
- Konturek, P.C.; Duda, A.; Brzozowski, T.; Konturek, S.J.; Kwiecien, S.; Drozdowicz, D.; Pajdo, R.; Meixner, H.; Hahn, E.G. Activation of genes for superoxide dismutase, interleukin-1beta, tumor necrosis factor-alpha, and intercellular adhesion molecule-1 during healing of ischemia-reperfusion-induced gastric injury. Scand. J. Gastroenterol. 2000, 35, 452–463. [Google Scholar]
- Yang, X.; Yang, L.; Pan, D.; Liu, H.; Xia, H.; Wang, S.; Sun, G. Wheat peptide protects against ethanol-induced gastric mucosal damage through downregulation of TLR4 and MAPK. J. Funct. Foods 2020, 75, 104271. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Arakawa, T.; Fukuda, T.; Sasaki, E.; Nakagawa, K.; Fujiwara, K.; Higuchi, K.; Kobayashi, K.; Tarnawski, A. Interleukin-8 stimulates leukocyte migration across a monolayer of cultured rabbit gastric epithelial cells. Effect associated with the impairment of gastric epithelial barrier function. Dig. Dis. Sci. 1997, 42, 1210–1215. [Google Scholar] [CrossRef]
- Watanabe, T.; Higuchi, K.; Tanigawa, T.; Tominaga, K.; Fujiwara, Y.; Arakawa, T. Mechanisms of peptic ulcer recurrence: Role of inflammation. Inflammopharmacology 2022, 10, 291–302. [Google Scholar] [CrossRef]
- De Araújo, E.R.D.; Guerra, G.C.B.; Araújo, D.F.d.S.; De Araújo, A.A.; Fernandes, J.M.; De Araújo Júnior, R.F.; Da Silva, V.C.; De Carvalho, T.G.; Ferreira, L.D.S.; Zucolotto, S.M. Gastroprotective and Antioxidant Activity of Kalanchoe brasiliensis and Kalanchoe pinnata Leaf Juices against Indomethacin and Ethanol-Induced Gastric Lesions in Rats. Int. J. Mol. Sci. 2018, 19, 1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, K.; Zhan, L.H.; Lu, T.T.; Zhou, C.; Chen, X.; Dong, Y.J.; Lv, G.Y.; Chen, S.H. Polysaccharides of Dendrobium officinale Kimura & Migo Leaves Protect Against Ethanol-Induced Gastric Mucosal Injury via the AMPK/mTOR Signaling Pathway in Vitro and vivo. Front. Pharmacol. 2020, 11, 526349. [Google Scholar]
- Nam, H.H.; Choo, B.K. Geranium koreanum, a medicinal plant Geranii Herba, ameliorate the gastric mucosal injury in gastritis-induced mice. J. Ethnopharmacol. 2021, 265, 113041. [Google Scholar] [CrossRef]
- Dumitriu, S. Polysaccharides: Structural Diversity and Functional Versatility; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Ju, H.; Yu, C.; Zhang, X.D.; Liu, W.; Wu, Y.C.; Gong, P.X.; Li, H.H.; Liu, Y.; Li, H.J. Recent trends in anti-cancer activities of terrestrial plants-based polysaccharides: A review. Carbohydr. Polym. Technol. Appl. 2023, 6, 100341. [Google Scholar] [CrossRef]
- Liao, H.F.; Chen, Y.; Yang, Y. A novel polysaccharide of black soybean promotes myelopoiesis and reconstitutes bone marrow after 5-flurouraciland irradiation-induced myelosuppression. Life Sci. 2005, 77, 400–413. [Google Scholar] [CrossRef]
- Wu, M.-H.; Lee, Y.-C.; Tsai, W.-J.; Yang, W.-B.; Chen, Y.-C.; Chuang, K.-A.; Liao, J.-F.; Wang, C.-C.; Kuo, Y.-C. Characterized Polysaccharides from Black Soybean Induce Granulocyte Colony-Stimulated Factor Gene Expression in a Phosphoinositide 3-Kinase-dependent Manner. Immunol. Investig. 2010, 40, 39–61. [Google Scholar] [CrossRef]
- Yao, Y.; Hu, G.; Gao, Y.; Wen, Z. White lentil polysaccharide inhibits hypoxic necrosis and apoptosis of nerve cells. Pharmacol. Clin. Appl. Tradit. Chin. Med. 2012, 03, 58–62. [Google Scholar] [CrossRef]
- Campos-Vega, R.; Guevara-Gonzalez, R.; Guevara-Olvera, B.; Oomah, B.D.; Loarca-Piña, G. Bean (Phaseolus vulgaris L.) polysaccharides modulate gene expression in human colon cancer cells (HT-29). Food Res. Int. 2010, 43, 1057–1064. [Google Scholar] [CrossRef]
- Cheng, A.; Wu, J.; Qin, H.; Yang, Q.; Liu, C.; Guo, X.; Sun, J. Study on the content and antioxidant activity of polyphenols and flavonoids in four types of legumes. Chin. J. Cereals Oils 2017, 32, 28–32. (In Chinese) [Google Scholar]
- Wu, G.-J.; Liu, D.; Wan, Y.-J.; Huang, X.-J.; Nie, S.-P. Comparison of hypoglycemic effects of polysaccharides from four legume species. Food Hydrocoll. 2018, 90, 299–304. [Google Scholar] [CrossRef]
- Lai, F.; Wen, Q.; Li, L.; Wu, H.; Li, X. Antioxidant activities of water-soluble polysaccharide extracted from mung bean (Vigna radiata L.) hull with ultrasonic assisted treatment. Carbohydr. Polym. 2010, 81, 323–329. [Google Scholar] [CrossRef]
- Qiang, Y.; Zhu, W.; Zhihan, G.; Xin, Q.; Siyu, W.; Tuoping, L.; Suhong, L. Optimization of Ultrasound Assisted Extraction of Cowpea Polysaccharides and in Vitro Antioxidant Study. J. Shenyang Agric. Univ. 2021, 52, 224–231. (In Chinese) [Google Scholar]
- Luo, A.; Fan, Y.; Tan, X.; Zhao, J.; Yang, K.; Wu, S.; Zhang, J.; Pu, S.; Wang, G. Screening and characterization of an acid polysaccharide with antioxidant activity in vitro and in vivo from Dendrobium aurantiacum var. denneanum (Kerr). Pharmacogn. Mag. 2022, 18, 44–51. [Google Scholar] [CrossRef]
- Fan, Y.; He, X.; Zhou, S.; Luo, A.; He, T.; Chun, Z. Composition analysis and antioxidant activity of polysaccharide from Dendrobium denneanum. Int. J. Biol. Macromol. 2009, 45, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Luo, A.; He, X.; Zhou, S.; Fan, Y.; Luo, A.; Chun, Z. Purification, composition analysis and antioxidant activity of the polysaccharides from Dendrobium nobile Lindl. Carbohydr. Polym. 2010, 79, 1014–1019. [Google Scholar] [CrossRef]
- Luo, A.; He, X.; Zhou, S.; Fan, Y.; He, T.; Chun, Z. In vitro antioxidant activities of a water-soluble polysaccharide derived from Dendrobium nobile Lindl. extracts. Int. J. Biol. Macromol. 2009, 45, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, R.; Liu, J.; Cheng, H.; Yu, N. Determination of monosaccharides in Lycium barbarum fruit polysaccharide by an efficient UHPLC-QTRAP-MS/MS method. Phytochem. Anal. 2021, 32, 785–793. [Google Scholar] [CrossRef]
- Liu, W.; Li, Z.; Feng, C.; Hu, S.; Yang, X.; Xiao, K.; Nong, Q.; Xiao, Q.; Wu, K.; Li, X.Q.; et al. The structures of two polysaccharides from Angelica sinensis and their effects on hepatic insulin resistance through blocking RAGE. Carbohydr. Polym. 2022, 280, 119001. [Google Scholar] [CrossRef]
- Wan, X.; Jin, X.; Wu, X.; Yang, X.; Lin, D.; Li, C.; Fu, Y.; Liu, Y.; Liu, X.; Lv, J.; et al. Structural characterisation and antitumor activity against non-small cell lung cancer of polysaccharides from Sanghuangporus vaninii. Carbohydr. Polym. 2021, 276, 118798. [Google Scholar] [CrossRef]
- Fan, Y.; Yu, Q.; Wang, G.; Tan, J.; Liu, S.; Pu, S.; Chen, W.; Xie, P.; Zhang, Y.; Zhang, J.; et al. Effects of Non-thermal Plasma Treatment on the Polysaccharide from Dendrobium nobile Lindl. and Its Immune Activities in vitro. Int. J. Biol. Macromol. 2020, 32, 393–396. [Google Scholar] [CrossRef]
- Quan, Z.; Guan, R.; Huang, H.; Yang, K.; Cai, M.; Meng, X. Antioxidant activity and absorption of cyanidin-3-O-glucoside liposomes in GES-1 cells in vitro. Biosci. Biotechnol. Biochem. 2020, 84, 1239–1249. [Google Scholar] [CrossRef]
- Hany, H.A.; Ahmed, M.A.; Amany, M.G.; Ayman, M.M.; Ahmed, M.K. Activation of AMPK/mTOR-driven autophagy and inhibition of NLRP3 inflammasome by saxagliptin ameliorate ethanol-induced gastric mucosal damage. Life Sci. 2021, 280, 119743. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Linkage Type | Methylation Product | Molecular Weight (MW) | RT (min) | Main MS (m/z) |
|---|---|---|---|---|
| t-Ara(f) | 1,4-di-O-acetyl-2,3,5-tri-O-methyl arabinitol | 279 | 5.71 | 71, 87, 101, 118, 129, 161 |
| t-Glc(p) | 1,5-di-O-acetyl-2,3,4,6-tetra-O-methyl glucitol | 323 | 8.58 | 59, 71, 87, 102, 118, 129, 145, 162, 205 |
| t-Gal(p) | 1,5-di-O-acetyl-2,3,4,6-tetra-O-methyl galactitol | 323 | 9.57 | 59, 71, 87, 102, 118, 129, 145, 157, 205 |
| 5-Ara(f) | 1,4,5-tri-O-acetyl-2,3-di-O-methyl arabinitol | 307 | 10.21 | 71, 87, 102, 118, 129, 189 |
| 2,4-Rha(p) | 1,2,4,5-tetra-O-acetyl-6-deoxy-3-O-methyl rhamnitol | 349 | 12.09 | 59, 74, 88, 101, 130, 143, 190, 203 |
| 4-Gal(p) | 1,4,5-tri-O-acetyl-2,3,6-tri-O-methyl galactitol | 351 | 13.32 | 71, 87, 99, 118, 131, 142, 157, 173, 233 |
| 4-Glc(p) | 1,4,5-tri-O-acetyl-2,3,6-tri-O-methyl glucitol | 351 | 13.65 | 71, 87, 99, 118, 129, 142, 157, 173, 233 |
| 3,4-Glc(p) | 1,3,4,5-tetra-O-acetyl-2,6-di-O-methyl glucitol | 379 | 15.82 | 59, 87, 118, 129, 160, 171, 185, 305 |
| 2,4-Gal(p) | 1,2,4,5-tetra-O-acetyl-3,6-di-O-methyl galactitol | 379 | 16.47 | 59, 71, 87, 99, 113, 130, 190, 233 |
| 4,6-Glc(p) | 1,4,5,6-tetra-O-acetyl-2,3-di-O-methyl glucitol | 379 | 17.88 | 59, 85, 101, 117, 159, 261 |
| Code | Glycosyl Residues | Chemical Shifts (ppm) | |||||
|---|---|---|---|---|---|---|---|
| H1/C1 | H2/C2 | H3/C3 | H4/C4 | H5/C5 | H6a,b/C6 | ||
| A | →4)-α-D-Glcp(1→ | 5.42 | 3.65 | 3.99 | 3.69 | 3.85 | 3.84/n.d |
| 102.45 | 74.38 | 76.1 | 79.46 | 74.03 | 63.24 | ||
| B | →4)-α-D-Galp(1→ | 4.99 | 3.78 | 3.92 | 3.65 | 3.87 | 3.85/n.d |
| 100.54 | 70.71 | 72.99 | 74.19 | 73.83 | 63.2 | ||
| C | α-D-Glcp(1→ | 4.66 | 3.7 | 3.85 | 3.43 | 3.86 | 3.65/n.d |
| 107.22 | 74.7 | 74.04 | 72.11 | 74.06 | 60.24 | ||
| D | →3,4)-β-D-Glcp(1→ | 5.1 | 3.54 | 4.14 | 4.02 | 3.97 | 3.79/n.d |
| 110.36 | 72.27 | 83.85 | 79.36 | 76.26 | 63.62 | ||
| E | →4,6)-α-D-Glcp(1→ | 5.1 | 3.59 | 3.96 | 3.73 | 3.84 | 3.9/n.d |
| 101.86 | 74.21 | 75.54 | 79.49 | 76.19 | 69.67 | ||
| F | →5)-α-L-Araf(1→ | 5.79 | 4.26 | 4.19 | n.d | 3.77,4.01 | - |
| 109.61 | 85.17 | 80.3 | n.d | 68.27 | - | ||
| Inflammatory Factors (pg/mg Prot) | Groups (μg/mL) | ||||
|---|---|---|---|---|---|
| Control | Model | 250 | 500 | 1000 | |
| IL-8 | 48.11 ± 2.78 ** | 125.6 ± 3.85 | 101.99 ± 6.38 | 82.75 ± 5.07 * | 68.05 ± 4.31 ** |
| TNF-α | 15.95 ± 4.74 ** | 58.19 ± 2.69 | 46.91 ± 3.73 | 33.16 ± 3.48 | 25.32 ± 2.68 * |
| IL-1β | 18.96 ± 2.77 ** | 46.34 ± 3.07 | 33.38 ± 1.99 | 28.57 ± 2.07 * | 19.58 ± 1.79 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, Y.; Xu, E.; Ma, J.; Li, X.; Liu, Y.; Xu, L.; Luo, A. Isolation, Structural Characteristics Analysis of a Vigna unguiculata Polysaccharide VUP80-3 and Its Protective Effect on GES-1 Cells In Vitro. Molecules 2023, 28, 5566. https://doi.org/10.3390/molecules28145566
Fan Y, Xu E, Ma J, Li X, Liu Y, Xu L, Luo A. Isolation, Structural Characteristics Analysis of a Vigna unguiculata Polysaccharide VUP80-3 and Its Protective Effect on GES-1 Cells In Vitro. Molecules. 2023; 28(14):5566. https://doi.org/10.3390/molecules28145566
Chicago/Turabian StyleFan, Yijun, Erya Xu, Jie Ma, Xuebing Li, Yuanyuan Liu, Linlong Xu, and Aoxue Luo. 2023. "Isolation, Structural Characteristics Analysis of a Vigna unguiculata Polysaccharide VUP80-3 and Its Protective Effect on GES-1 Cells In Vitro" Molecules 28, no. 14: 5566. https://doi.org/10.3390/molecules28145566