

Effects of Solid Fermentation on Polygonatum cyrtonema Polysaccharides: Isolation, Characterization and Bioactivities

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Extraction, Purification and Preliminary Characterization of P. cyrtonema Polysaccharide
2.1.1. Extraction of Crude Polysaccharides
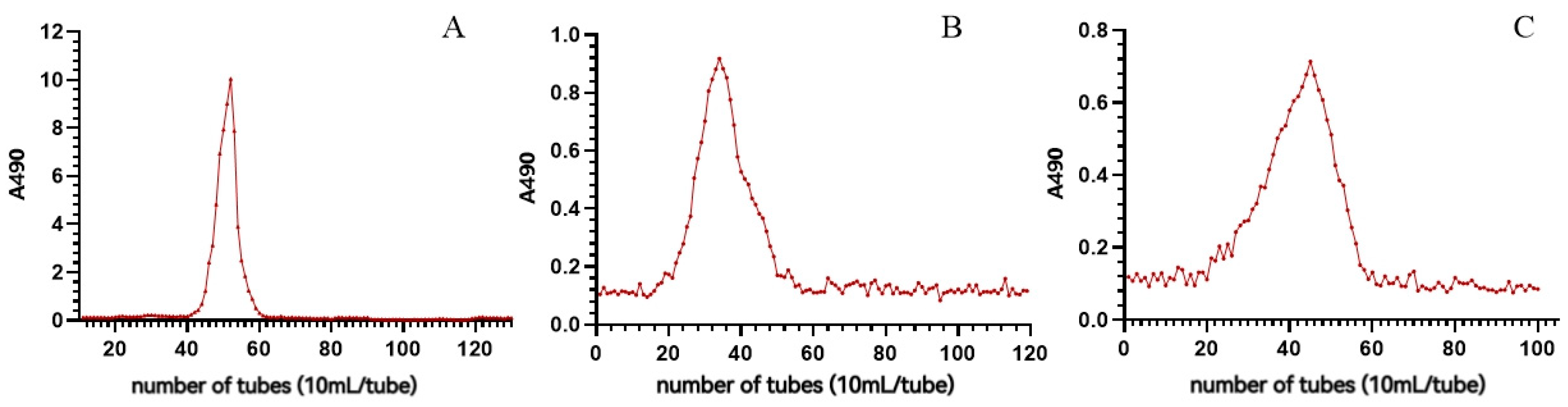
2.1.2. Purification of P. cyrtonema Polysaccharide
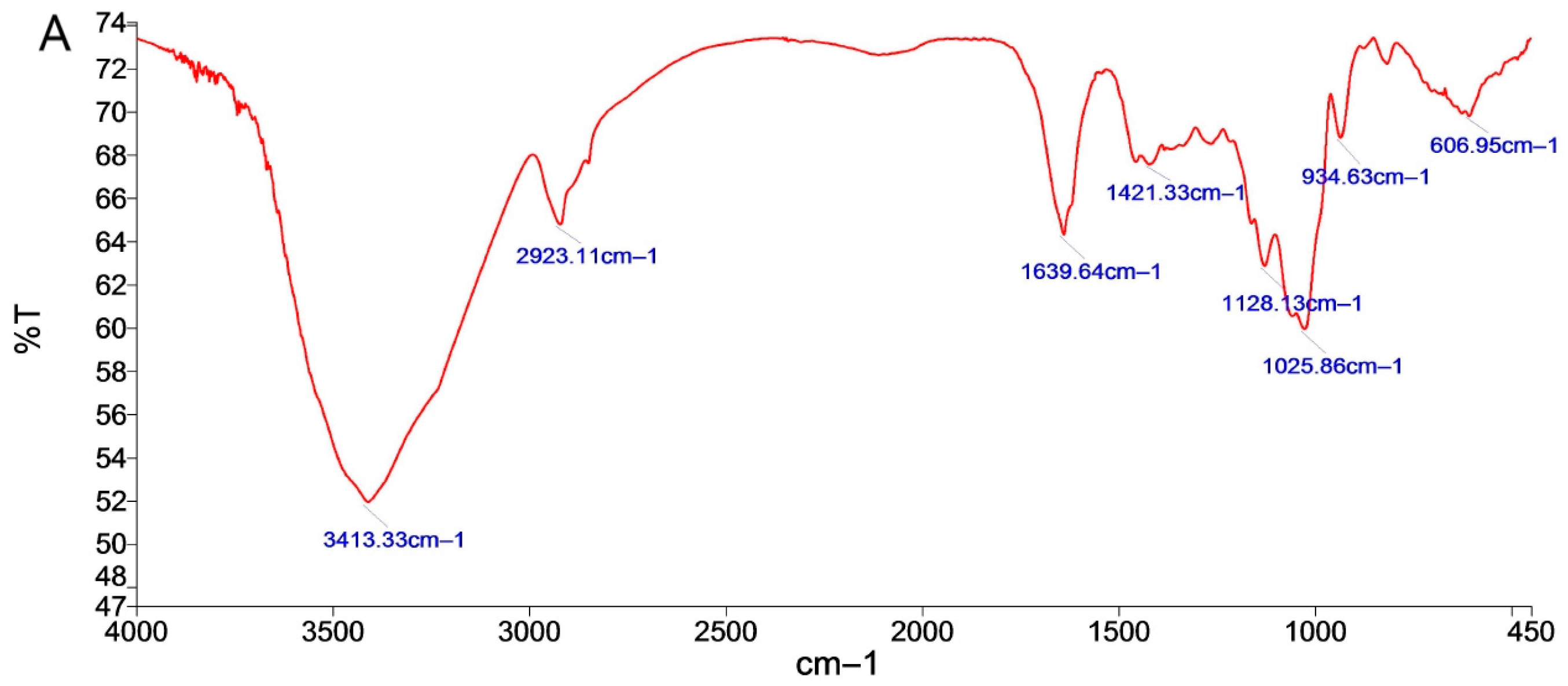
2.1.3. FT-IR Analysis of P. cyrtonema Neutral Polysaccharide
2.1.4. FT-IR Analysis of P. cyrtonema Neutral Polysaccharides
2.2. Fermentation Affects the In Vitro Bioactivities of P. cyrtonema Polysaccharides
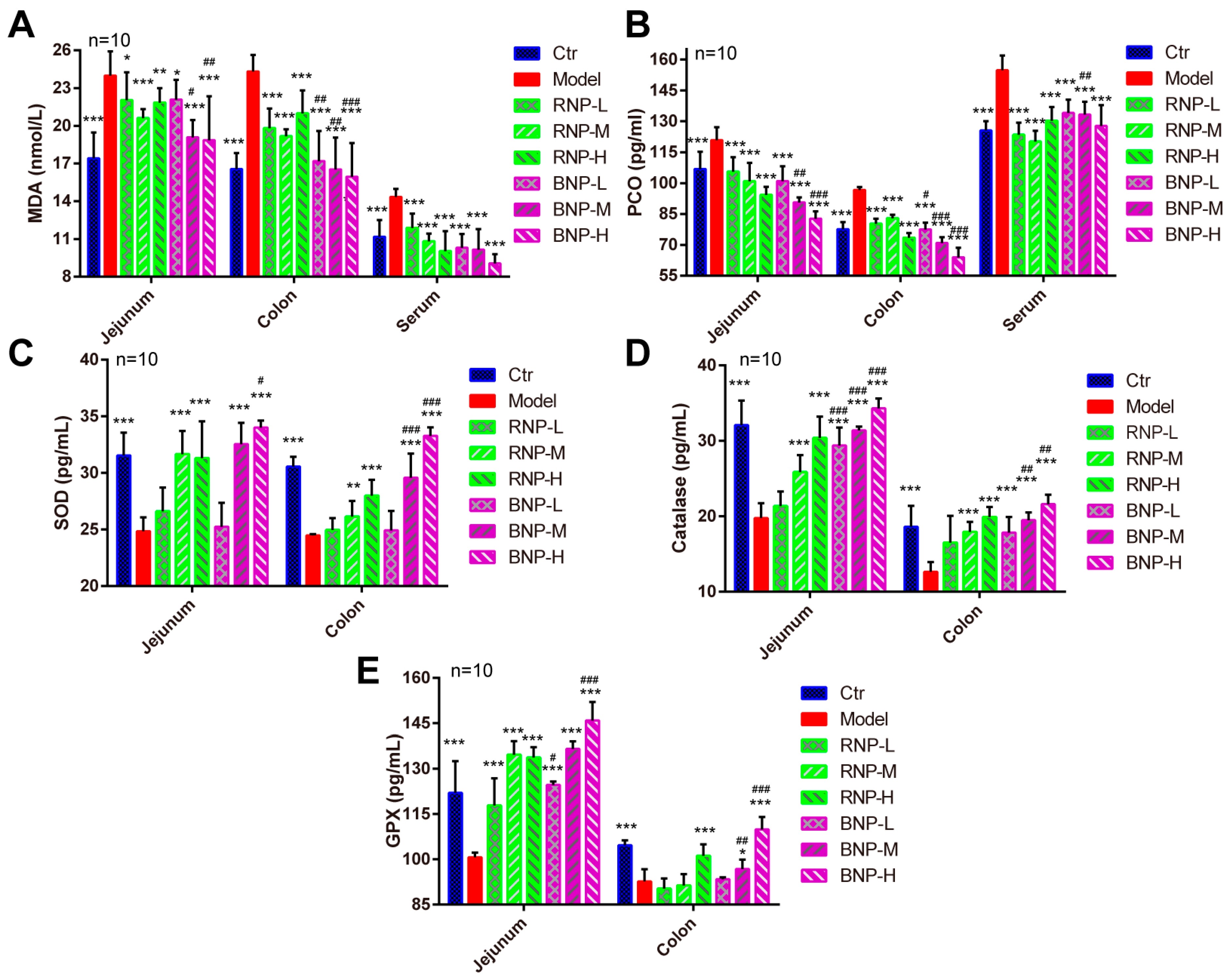
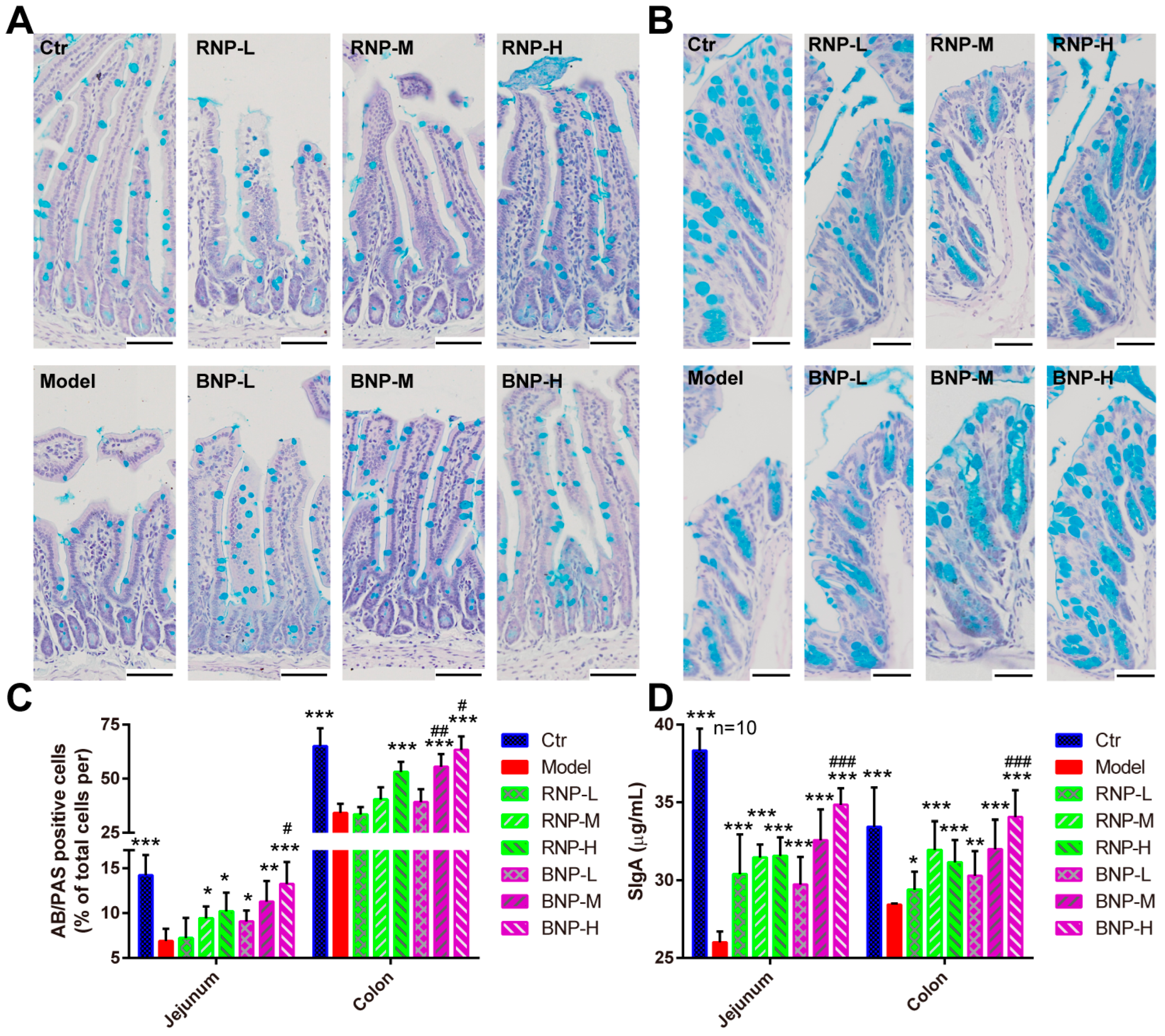
2.3. BNP Attenuates Intestinal Oxidative Stress in the Aging Mouse Model More Effectively
2.4. Immunomodulation Effects of BNP in the Immunosuppressed Mouse Model
2.5. BNP Modulates the Homeostasis of Gut Microbiota Affected by CTX in Mice
3. Discussion
4. Materials and Methods
4.1. Solid Fermentation of P. cyrtonema
4.2. Extraction, Purification and Preliminary Characterization of Polygonati Rhizoma Polysaccharide
4.2.1. Extraction of Crude Polysaccharides
4.2.2. Purification of Crude Polysaccharides
4.2.3. Chemical Composition and FT-IR Analysis of Polysaccharide Fractions
4.3. Animals and Treatments
4.3.1. Animals
4.3.2. Generation of Aging Mouse Model
4.3.3. Generation of Immunosuppressed Mouse Model
4.4. Biochemical Assay and Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Gut Microbiota Analysis
4.6. Histological Staining
4.7. Cell Culture
4.8. Quantitative Realtime PCR
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xie, Y.; Mu, C.; Kazybay, B.; Sun, Q.; Kutzhanova, A.; Nazarbek, G.; Xu, N.; Nurtay, L.; Wang, Q.; Amin, A.; et al. Network pharmacology and experimental investigation of Rhizoma polygonati extract targeted kinase with herbzyme activity for potent drug delivery. Drug Deliv. 2021, 28, 2187–2197. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Jia, X.; Liu, P.; Zhang, Q.; Liu, F.; Ma, C.; Xi, P.; Liang, Z. Species identification of Polygonati Rhizoma in China by both morphological and molecular marker methods. Comptes Rendus Biol. 2018, 341, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Xu, C.; Zhou, W.; Li, Y.; Xie, Y.; Hu, H.; Wang, Z. Polygonati Rhizoma with the homology of medicine and food: A review of ethnopharmacology, botany, phytochemistry, pharmacology and applications. J. Ethnopharmacol. 2023, 309, 116296. [Google Scholar] [CrossRef]
- Li, L.X.; Feng, X.; Tao, M.T.; Paulsen, B.S.; Huang, C.; Feng, B.; Liu, W.; Yin, Z.Q.; Song, X.; Zhao, X.; et al. Benefits of neutral polysaccharide from rhizomes of Polygonatum sibiricum to intestinal function of aged mice. Front. Nutr. 2022, 9, 992102. [Google Scholar] [CrossRef]
- Liu, N.; Dong, Z.; Zhu, X.; Xu, H.; Zhao, Z. Characterization and protective effect of Polygonatum sibiricum polysaccharide against cyclophosphamide-induced immunosuppression in Balb/c mice. Int. J. Biol. Macromol. 2018, 107 Pt A, 796–802. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, N.; Sun, C.; Sun, D.; Wang, Y. Polysaccharides from Polygonatum sibiricum Delar. ex Redoute induce an immune response in the RAW264.7 cell line via an NF-kappaB/MAPK pathway. RSC Adv. 2019, 9, 17988–17994. [Google Scholar] [CrossRef]
- Yelithao, K.; Surayot, U.; Park, W.; Lee, S.; Lee, D.H.; You, S. Effect of sulfation and partial hydrolysis of polysaccharides from Polygonatum sibiricum on immune-enhancement. Int. J. Biol. Macromol. 2019, 122, 10–18. [Google Scholar] [CrossRef]
- Li, L.; Thakur, K.; Liao, B.Y.; Zhang, J.G.; Wei, Z.J. Antioxidant and antimicrobial potential of polysaccharides sequentially extracted from Polygonatum cyrtonema Hua. Int. J. Biol. Macromol. 2018, 114, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Lv, Y.; Dai, W.; Miao, X.; Zhong, D. Extraction and bioactivity of polygonatum polysaccharides. Int. J. Biol. Macromol. 2013, 54, 131–135. [Google Scholar] [CrossRef]
- He, Y.; Chen, Z.; Nie, X.; Wang, D.; Zhang, Q.; Peng, T.; Zhang, C.; Wu, D.; Zhang, J. Recent advances in polysaccharides from edible and medicinal Polygonati Rhizoma: From bench to market. Int. J. Biol. Macromol. 2022, 195, 102–116. [Google Scholar] [CrossRef]
- Li, L.; Wang, L.; Fan, W.; Jiang, Y.; Zhang, C.; Li, J.; Peng, W.; Wu, C. The Application of Fermentation Technology in Traditional Chinese Medicine: A Review. Am. J. Chin. Med. 2020, 48, 899–921. [Google Scholar] [CrossRef]
- Wu, T.X.; Wang, N.; Zhang, Y.; Xu, X.B. Advances in the study on microbial fermentation and transformation of traditional Chinese medicine. Afr. J. Microbiol. Res. 2013, 7, 1644–1650. [Google Scholar]
- Chen, H. Modern Solid State Fermentation; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Yao, X.; Deng, Z.; Li, H.; Zhang, B. Effect of processing cycles on the composition of Polygonatum cyrtonema Hua during nine-steam-nine-bask processing. Food Biosci. 2022, 50, 102081. [Google Scholar] [CrossRef]
- Wang, Z.; Lao, J.; Kang, X.; Xie, Z.; He, W.; Liu, X.; Zhong, C.; Zhang, S.; Jin, J. Insights into the metabolic profiling of Polygonati Rhizoma fermented by Lactiplantibacillus plantarum under aerobic and anaerobic conditions using a UHPLC-QE-MS/MS system. Front. Nutr. 2023, 10, 1093761. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.-L.; Liu, L.; Li, Z.; Li, Y.; Han, B. Correlation between Effective Constituent Content and Color During Fermentation of Polygonati Rhizoma. Chin. J. Exp. Tradit. Med. Formulae 2020, 24, 169–176. [Google Scholar]
- Han, L.; Suo, Y.; Yang, Y.; Meng, J.; Hu, N. Optimization, characterization, and biological activity of polysaccharides from Berberis dasystachya Maxim. Int. J. Biol. Macromol. 2016, 85, 655–666. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, X.; Zhang, J.; Liu, G.; Liu, Y.; Wang, K.; Yang, M.; Cheng, H.; Zhao, Z. Structural characterization and in vitro antitumor activity of polysaccharides from Zizyphus jujuba cv. Muzao. RSC Adv. 2015, 5, 7860–7867. [Google Scholar] [CrossRef]
- Yan, W.; Peng, D.; Chenzhong, J.; Jinhe, K.; Yihong, H.; Zheng, W. Analysis on Composition of Polygonatum sibiricum Polysaccharide and Its Anti-oxidant Activity. Genom. Appl. Biol. 2019, 38, 9. [Google Scholar]
- Liu, W.; Wang, H.; Yu, J.; Liu, Y.; Lu, W.; Chai, Y.; Liu, C.; Pan, C.; Yao, W.; Gao, X. Structure, chain conformation, and immunomodulatory activity of the polysaccharide purified from Bacillus Calmette Guerin formulation. Carbohydr. Polym. 2016, 150, 149–158. [Google Scholar] [CrossRef]
- Wu, Q.; Hu, S.; Yang, G.; Mei, Z. Separation, Purification and Structural Studies of Polysaccharides from Polygonatum kingianum. Chem. Ind. For. Prod. 2005, 25, 3. [Google Scholar]
- Yi, W.; Xiaohua, Y.; Huafeng, Z.; Lei, P.; Zhezhi, W. Structure of Polygonatum kingjanttm Polysaccharides and Its Inhibitory Activity against α-Glucosidase. Fine Chem. 2019, 36, 6. [Google Scholar]
- Wan, L.; Liu, Y.; Liu, W.; Hu, Y.; Du, L.; He, X. Identification of Red Ginseng Polysaccharides in Shenshu Compound Polysaccharides by Infrared Spectroscopy. Chin. Tradit. Pat. Med. 2006, 28, 2. [Google Scholar]
- Santos, J.; Escobar-Avello, D.; Magalhães, P.; Magalhães, F.D.; Martins, J.M.; González-Álvarez, J.; Carvalho, L.H. High-value compounds obtained from grape canes (Vitis vinifera L.) by steam pressure alkali extraction. Food Bioprod. Process. 2022, 133, 153–167. [Google Scholar] [CrossRef]
- Xu, M.; Yin, J.; Wan, Y.; Niu, S.; Xie, M. Physicochemical property of different water-soluble polysaccharide from carrot juice fermented by probiotics. Sci. Technol. Food Ind. 2016, 37, 4. [Google Scholar]
- Wang, F.; Jiang, Y.; Jin, S.; Wang, D.; Wu, K.; Yang, Q.; Han, R.; Chen, S.; Liang, Z.; Jia, Q. Structure characterization and bioactivity of neutral polysaccharides from different sources of Polygonatum Mill. Biopolymers 2022, 113, e23490. [Google Scholar] [CrossRef]
- Luan, Y.; Jiang, Y.; Huang, R.; Wang, X.; He, X.; Liu, Y.; Tan, P. Polygonati Rhizoma Polysaccharide Prolongs Lifespan and Healthspan in Caenorhabditis elegans. Molecules 2023, 28, 2235. [Google Scholar] [CrossRef]
- Ilyasov, I.R.; Beloborodov, V.L.; Selivanova, I.A.; Terekhov, R.P. ABTS/PP Decolorization Assay of Antioxidant Capacity Reaction Pathways. Int. J. Mol. Sci. 2020, 21, 1131. [Google Scholar] [CrossRef] [Green Version]
- Ozyurt, D.; Demirata, B.; Apak, R. Determination of total antioxidant capacity by a new spectrofluorometric method based on Ce(IV) reduction: Ce(III) fluorescence probe for CERAC assay. J. Fluoresc. 2011, 21, 2069–2076. [Google Scholar] [CrossRef]
- Azman, K.F.; Zakaria, R. D-Galactose-induced accelerated aging model: An overview. Biogerontology 2019, 20, 763–782. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef]
- Pirinccioglu, A.G.Z.; Gökalp, D.; Pirinccioglu, M.; Kizil, G.K.; Kizil, M. Malondialdehyde (MDA) and protein carbonyl (PCO) levels as biomarkers of oxidative stress in subjects with familial hypercholesterolemia. Clin. Biochem. 2010, 43, 1220–1224. [Google Scholar] [CrossRef]
- Yan, H.; Lu, J.; Wang, J.; Chen, L.; Wang, Y.; Li, L.; Miao, L.; Zhang, H. Prevention of Cyclophosphamide-Induced Immunosuppression in Mice with Traditional Chinese Medicine Xuanfei Baidu Decoction. Front. Pharmacol. 2021, 12, 730567. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Sun, L.; Gonzalez, F.J. Gut microbiota-derived bile acids in intestinal immunity, inflammation, and tumorigenesis. Cell Host Microbe 2022, 30, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhang, Z.; Cheng, L.; Zhang, X.; Liu, Y.; Zhang, R.; Weng, P.; Wu, Z. Polysaccharides confer benefits in immune regulation and multiple sclerosis by interacting with gut microbiota. Food Res. Int. 2021, 149, 110675. [Google Scholar] [CrossRef]
- Song, Q.; Wang, Y.; Huang, L.; Shen, M.; Yu, Y.; Yu, Q.; Chen, Y.; Xie, J. Review of the relationships among polysaccharides, gut microbiota, and human health. Food Res. Int. 2021, 140, 109858. [Google Scholar] [CrossRef]
- Murray, B.R.; Rice, B.L.; Keith, D.A.; Myerscough, P.J.; Howell, J.; Floyd, A.G.; Mills, K.; Westoby, M. Species in the tail of rank–abundance curves. Ecology 1999, 80, 1806–1816. [Google Scholar]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Statist. 1984, 11, 265–270. [Google Scholar]
- Wang, S.; Schuurmans, D.; Peng, F.; Zhao, Y. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar]
- Mello, J.F.; Buzas, M.A. An application of cluster analysis as a method of determining biofacies. J. Paleontol. 1968, 42, 747–758. [Google Scholar]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruskal, J.B.; Wish, M. Multidimensional Scaling; Sage: Newbury Park, CA, USA, 1978; Volume 11. [Google Scholar]
- Zhang, L.X.; Gao, W.Y.; Wang, H.Y. Review of traditional Chinese medicine processed by fermentation. China J. Chin. Mater. Medica 2012, 37, 3695–3700. [Google Scholar]
- Guo, X.; Yan, Z.; Wang, J.; Fan, X.; Kang, J.; Niu, R.; Sun, Z. Effect of traditional chinese medicine (TCM) and its fermentation using Lactobacillus plantarum on ceftriaxone sodium-induced dysbacteriotic diarrhea in mice. Chin. Med. 2022, 17, 20. [Google Scholar] [CrossRef] [PubMed]
- Duan, W.; Guan, Q.; Zhang, H.-L.; Wang, F.-Z.; Lu, R.; Li, D.-M.; Geng, Y.; Xu, Z.-H. Improving flavor, bioactivity, and changing metabolic profiles of goji juice by selected lactic acid bacteria fermentation. Food Chem. 2023, 408, 135–155. [Google Scholar] [CrossRef]
- Liu, Y.; Cheng, H.; Liu, H.; Ma, R.; Ma, J.; Fang, H. Fermentation by Multiple Bacterial Strains Improves the Production of Bioactive Compounds and Antioxidant Activity of Goji Juice. Molecules 2019, 24, 3519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Qu, Q.; Zhao, C.; Liu, X.; Yang, P.; Li, Z.; Han, L.; Shi, X. Paecilomyces cicadae-fermented Radix astragali activates podocyte autophagy by attenuating PI3K/AKT/mTOR pathways to protect against diabetic nephropathy in mice. Biomed. Pharmacother. 2020, 129, 110479. [Google Scholar] [CrossRef]
- Mei, Z.G.; Tan, L.J.; Wang, J.F.; Li, X.L.; Huang, W.F.; Zhou, H.J. Fermented Chinese formula Shuan-Tong-Ling attenuates ischemic stroke by inhibiting inflammation and apoptosis. Neural Regen. Res. 2017, 12, 425–432. [Google Scholar] [CrossRef]
- Sánchez, O.J.; Montoya, S.; Vargas, L.M. Polysaccharide production by submerged fermentation. In Polysaccharides; Springer: Berlin/Heidelberg, Germany, 2015; pp. 451–473. [Google Scholar]
- Sun, T.; Wang, R.; Sun, D.; Li, S.; Xu, H.; Qiu, Y.; Lei, P.; Sun, L.; Xu, X.; Zhu, Y. High-efficiency production of Tremella aurantialba polysaccharide through basidiospore fermentation. Bioresour. Technol. 2020, 318, 124268. [Google Scholar] [CrossRef]
- Wang, Y.; Shao, S.; Xu, P.; Chen, H.; Lin-Shiau, S.-Y.; Deng, Y.-T.; Lin, J.-K. Fermentation process enhanced production and bioactivities of oolong tea polysaccharides. Food Res. Int. 2012, 46, 158–166. [Google Scholar] [CrossRef]
- Wang, S.; Liu, L. Fermentation effect of Bacillus subtilis on traditional Chinese medicine. Chin. Community Dr. 2019, 35, 15. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, R.; Wang, Y.; An, X.; Liu, N.; Song, M.; Yang, Y.; Yin, N.; Qi, J. Characterization and antioxidant activity of wheat bran polysaccharides modified by Saccharomyces cerevisiae and Bacillus subtilis fermentation. J. Cereal Sci. 2021, 97, 103157. [Google Scholar] [CrossRef]
- Cockburn, D.W.; Koropatkin, N.M. Polysaccharide Degradation by the Intestinal Microbiota and Its Influence on Human Health and Disease. J. Mol. Biol. 2016, 428, 3230–3252. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Willför, S.; Xu, C. A review of bioactive plant polysaccharides: Biological activities, functionalization, and biomedical applications. Bioact. Carbohydr. Diet. Fibre 2015, 5, 31–61. [Google Scholar] [CrossRef]
- Foster, J.A.; Rinaman, L.; Cryan, J.F. Stress & the gut-brain axis: Regulation by the microbiome. Neurobiol. Stress 2017, 7, 124–136. [Google Scholar] [PubMed] [Green Version]
- Tripathi, A.; Debelius, J.; Brenner, D.A.; Karin, M.; Loomba, R.; Schnabl, B.; Knight, R. The gut-liver axis and the intersection with the microbiome. Nature reviews. Gastroenterol. Hepatol. 2018, 15, 397–411. [Google Scholar]
- Marsland, B.J.; Trompette, A.; Gollwitzer, E.S. The Gut-Lung Axis in Respiratory Disease. Ann. Am. Thorac. Soc. 2015, 12 (Suppl. 2), S150–S156. [Google Scholar] [CrossRef]
- Salem, I.; Ramser, A.; Isham, N.; Ghannoum, M.A. The gut microbiome as a major regulator of the gut-skin axis. Front. Microbiol. 2018, 9, 1459. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Lan, X.; Peng, X.; Shi, S.; Zhao, Y.; Liu, W.; Luo, Q.; Jia, L.; Feng, B.; Chen, Z.; et al. Polysaccharide from Salviae miltiorrhizae Radix et Rhizoma Attenuates the Progress of Obesity-Induced Non-Alcoholic Fatty Liver Disease through Modulating Intestinal Microbiota-Related Gut-Liver Axis. Int. J. Mol. Sci. 2022, 23, 10620. [Google Scholar] [CrossRef]
- Huang, P.; Wang, P.; Xu, J.; Sun, M.; Liu, X.; Lin, Q.; Liu, W.; Qing, Z.; Zeng, J. Fermented traditional Chinese medicine in diet altered the composition of intestine microbiota in broiler chickens. Res. Vet. Sci. 2020, 135, 8–14. [Google Scholar] [CrossRef]
- Lv, W.; Ma, Y.; Zhang, Y.; Wang, T.; Huang, J.; He, S.; Du, H.; Guo, S. Effects of Lactobacillus plantarum fermented Shenling Baizhu San on gut microbiota, antioxidant capacity, and intestinal barrier function of yellow-plumed broilers. Front. Vet. Sci. 2023, 10, 1103023. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Yi, X.; Yang, J.; Zhu, Z.; Wang, Y.; Liu, X.; Huang, X.; Wan, Y.; Fu, X.; Shu, W.; et al. Gut Microbiome Signatures in the Progression of Hepatitis B Virus-Induced Liver Disease. Front. Microbiol. 2022, 13, 916061. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.F.; Trevisi, P.; Coelho, D.F.; Correa, F.; Ribeiro, D.M.; Alfaia, C.M.; Pinho, M.; Pestana, J.M.; Mourato, M.P.; Almeida, A.M.; et al. Influence of Chlorella vulgaris on growth, digestibility and gut morphology and microbiota of weaned piglet. Sci. Rep. 2022, 12, 6012. [Google Scholar] [CrossRef] [PubMed]
- Mager, L.F.; Burkhard, R.; Pett, N.; Cooke, N.C.A.; Brown, K.; Ramay, H.; Paik, S.; Stagg, J.; Groves, R.A.; Gallo, M.; et al. Microbiome-derived inosine modulates response to checkpoint inhibitor immunotherapy. Science 2020, 369, 1481–1489. [Google Scholar] [CrossRef]
- Truax, A.D.; Chen, L.; Tam, J.W.; Cheng, N.; Guo, H.; Koblansky, A.A.; Chou, W.C.; Wilson, J.E.; Brickey, W.J.; Petrucelli, A.; et al. The Inhibitory Innate Immune Sensor NLRP12 Maintains a Threshold against Obesity by Regulating Gut Microbiota Homeostasis. Cell Host Microbe 2018, 24, 364–378.e6. [Google Scholar] [CrossRef] [Green Version]
- Sinha, S.R.; Haileselassie, Y.; Nguyen, L.P.; Tropini, C.; Wang, M.; Becker, L.S.; Sim, D.; Jarr, K.; Spear, E.T.; Singh, G.; et al. Dysbiosis-Induced Secondary Bile Acid Deficiency Promotes Intestinal Inflammation. Cell Host Microbe 2020, 27, 659–670.e5. [Google Scholar] [CrossRef]
- Shang, Q.; Shan, X.; Cai, C.; Hao, J.; Li, G.; Yu, G. Dietary fucoidan modulates the gut microbiota in mice by increasing the abundance of Lactobacillus and Ruminococcaceae. Food Funct. 2016, 7, 3224–3232. [Google Scholar] [CrossRef]
- Takahashi, K.; Nishida, A.; Fujimoto, T.; Fujii, M.; Shioya, M.; Imaeda, H.; Inatomi, O.; Bamba, S.; Sugimoto, M.; Andoh, A. Reduced Abundance of Butyrate-Producing Bacteria Species in the Fecal Microbial Community in Crohn’s Disease. Digestion 2016, 93, 59–65. [Google Scholar] [CrossRef]
- Rangel, I.; Sundin, J.; Fuentes, S.; Repsilber, D.; Vos, W.; Brummer, R.J. The relationship between faecal-associated and mucosal-associated microbiota in irritable bowel syndrome patients and healthy subjects. Aliment. Pharmacol. Ther. 2015, 42, 1211–1221. [Google Scholar] [CrossRef] [Green Version]
- Walters, W.A.; Xu, Z.; Knight, R. Meta-analyses of human gut microbes associated with obesity and IBD. FEBS Lett. 2014, 588, 4223–4233. [Google Scholar] [CrossRef] [Green Version]
- Keren, N.; Konikoff, F.M.; Paitan, Y.; Gabay, G.; Reshef, L.; Naftali, T.; Gophna, U. Interactions between the intestinal microbiota and bile acids in gallstones patients. Environ. Microbiol. Rep. 2015, 7, 874–880. [Google Scholar] [CrossRef] [PubMed]
- Lam, Y.Y.; Ha, C.W.Y.; Campbell, C.R.; Mitchell, A.J.; Dinudom, A.; Oscarsson, J.; Cook, D.I.; Hunt, N.H.; Caterson, I.D.; Holmes, A.J. Increased Gut Permeability and Microbiota Change Associate with Mesenteric Fat Inflammation and Metabolic Dysfunction in Diet-Induced Obese Mice. PLoS ONE 2012, 7, e34233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konikoff, T.; Gophna, U. Oscillospira: A Central, Enigmatic Component of the Human Gut Microbiota. Trends Microbiol. 2016, 24, 523–524. [Google Scholar] [CrossRef]
- Boutard, M.; Cerisy, T.; Nogue, P.Y.; Alberti, A.; Weissenbach, J.; Salanoubat, M.; Tolonen, A.C. Functional diversity of carbohydrate-active enzymes enabling a bacterium to ferment plant biomass. PLoS Genet. 2014, 10, e1004773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rostaher, A.; Morsy, Y.; Favrot, C.; Unterer, S.; Schnyder, M.; Scharl, M.; Fischer, N.M. Comparison of the Gut Microbiome between Atopic and Healthy Dogs—Preliminary Data. Animals 2022, 12, 2377. [Google Scholar] [CrossRef]
- Kalyana Chakravarthy, S.; Jayasudha, R.; Sai Prashanthi, G.; Ali, M.H.; Sharma, S.; Tyagi, M.; Shivaji, S. Dysbiosis in the Gut Bacterial Microbiome of Patients with Uveitis, an Inflammatory Disease of the Eye. Indian J. Microbiol. 2018, 58, 457–469. [Google Scholar] [CrossRef]
- Khattab, M.; Tawab, A.; Fouad, M.T. Isolation and Characterization of Anaerobic Bacteria from Frozen Rumen Liquid and its Potential Characterizations. Int. J. Dairy Sci. 2017, 12, 47–51. [Google Scholar] [CrossRef] [Green Version]
- Beerens, H.; Romond, C. Sulfate-reducing anaerobic bacteria in human feces. Am. J. Clin. Nutr. 1977, 30, 1770–1776. [Google Scholar] [CrossRef]
- Kushkevych, I.; Dordevic, D.; Kollar, P. Analysis of Physiological Parameters of Desulfovibrio Strains from Individuals with Colitis. Open Life Sci. 2018, 13, 481–488. [Google Scholar] [CrossRef]
- Zhai, Z.; Zhang, F.; Cao, R.; Ni, X.; Deng, B.; Cecropin, A. Alleviates Inflammation Through Modulating the Gut Microbiota of C57BL/6 Mice With DSS-Induced IBD. Front. Microbiol. 2019, 10, 1595. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, S.; Tang, Y.; You, T.; Xu, H. Lactobacillus rhamnosus GG Ameliorated Long-Term Exposure to TiO2 Nanoparticles Induced Microbiota-Mediated Liver and Colon Inflammation and Fructose-Caused Metabolic Abnormality in Metabolism Syndrome Mice. J. Agric. Food Chem. 2021, 69, 9788–9799. [Google Scholar] [CrossRef]
- Papapetropoulos, A.; Pyriochou, A.; Szabo, C. Hydrogen sulfide is an endogenous stimulator of angiogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 21972–21977. [Google Scholar] [CrossRef]
- Waters, J.L.; Ley, R.E. The human gut bacteria Christensenellaceae are widespread, heritable, and associated with health. BMC Biol. 2019, 17, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhu, L.; Li, X.; Wang, X.; Hao, R.; Li, J. Effects of high fructose corn syrup on intestinal microbiota structure and obesity in mice. NPJ Sci. Food 2022, 6, 17. [Google Scholar] [CrossRef]
- Ye, J.; Zhang, C.; Fan, Q.; Lin, X.; Wang, Y.; Azzam, M.; Alhotan, R.; Alqhtani, A.; Jiang, S. Antrodia cinnamomea polysaccharide improves liver antioxidant, anti-inflammatory capacity, and cecal flora structure of slow-growing broiler breeds challenged with lipopolysaccharide. Front. Vet. Sci. 2022, 9, 994782. [Google Scholar] [CrossRef] [PubMed]
- Lozano, C.P.; Wilkens, L.R.; Shvetsov, Y.B.; Maskarinec, G.; Park, S.Y.; Shepherd, J.A.; Boushey, C.J.; Hebert, J.R.; Wirth, M.D.; Ernst, T.; et al. Associations of the Dietary Inflammatory Index with total adiposity and ectopic fat through the gut microbiota, LPS, and C-reactive protein in the Multiethnic Cohort-Adiposity Phenotype Study. Am. J. Clin. Nutr. 2022, 115, 1344–1356. [Google Scholar] [CrossRef]
- Hao, G.; Li, P.; Huang, J.; Cui, K.; Liang, L.; Lin, F.; Lu, Z.; Sun, S. Research Note: Therapeutic effect of a Salmonella phage combination on chicks infected with Salmonella typhimurium. Poult. Sci. 2023, 7, 102715. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Peng, X.; Pang, D.-J.; Li, J.; Paulsen, B.S.; Rise, F.; Chen, Y.-L.; Chen, Z.-L.; Jia, R.-Y.; Li, L.-X.; et al. Pectic polysaccharide from Nelumbo nucifera leaves promotes intestinal antioxidant defense in vitro and in vivo. Food Funct. 2021, 12, 10828–10841. [Google Scholar] [CrossRef]
- Singleton, V.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Yield (%) | Polysaccharide Content (%) | Polyphenol Content (%) | Protein Content (%) |
|---|---|---|---|---|
| R | 15.57 ± 0.30 | 61.18 ± 0.18 | 0.15 ± 0.01 | 0.24 ± 0.04 |
| B | 12.57 ± 0.52 | 56.41 ± 0.13 | 0.67 ± 0.05 | 0.42 ± 0.03 |
| S | 13.22 ± 0.48 | 50.04 ± 0.23 | 0.68 ± 0.02 | 0.28 ± 0.06 |
| Samples | Neutral Component | Neutral Sugar (%) | Acidic Component | Acidic Sugar (%) |
|---|---|---|---|---|
| RP | RNP | 68.48 ± 0.09 | RAP | 31.52 ± 0.10 |
| BP | BNP | 64.10 ± 0.13 | BAP | 35.90 ± 0.21 |
| SP | SNP | 47.80 ± 0.17 | SAP | 52.20 ± 0.11 |
| Samples | Polysaccharide Content (%) | Polyphenol Content (%) | Protein Content (%) |
|---|---|---|---|
| RNP | 73.90 ± 0.14 | 0.24 ± 0.04 | 0.39 ± 0.04 |
| RAP | 64.38 ± 0.09 | 0.18 ± 0.02 | 0.42 ± 0.02 |
| BNP | 68.51 ± 0.15 | 0.32 ± 0.04 | 0.53 ± 0.05 |
| BAP | 59.87 ± 0.12 | 0.21 ± 0.01 | 0.47 ± 0.06 |
| SNP | 68.69 ± 0.17 | 0.26 ± 0.02 | 0.35 ± 0.02 |
| SAP | 65.24 ± 0.14 | 0.15 ± 0.02 | 0.29 ± 0.02 |
| Name | Company | Article Number |
|---|---|---|
| Antioxidant Capacity Assay Kit with ABTS | Mlbio (Shanghai, China) | ml094998 |
| TAC assay kit | Mlbio (Shanghai, China) | ml076332 |
| IL-2 ELISA kit | Mlbio (Shanghai, China) | ml063136 |
| sIgA ELISA kit | Mlbio (Shanghai, China) | ml001917 |
| IFN-γ ELISA kit | Mlbio (Shanghai, China) | ml002277 |
| GSH-Px ELISA kit | Mlbio (Shanghai, China) | ml058194 |
| Occludin ELISA kit | Mlbio (Shanghai, China) | ml063481 |
| SOD activity assay kit | Mlbio (Shanghai, China) | ml095266 |
| SOD ELISA kit | Mlbio (Shanghai, China) | ml643059 |
| IgG ELISA kit | Mlbio (Shanghai, China) | ml037601 |
| TNF-α ELISA kit | Mlbio (Shanghai, China) | ml002095 |
| MDA assay kit | Mlbio (Shanghai, China) | ml094963 |
| Catalase ELISA kit | Mlbio (Shanghai, China) | ml037752 |
| PCO ELISA kit | Mlbio (Shanghai, China) | ml058321 |
| Gene | Primer | Sequence |
|---|---|---|
| β-actin | F | 5′-CATCCTGCGTCTGGACCTGG-3′ |
| R | 5′-CAATAGTGATGACCTGGCCGT-3′ | |
| IL-4 | F | 5′-TTCGGCACATCTACAGACACC-3′ |
| R | 5′-TTCATGCACAGAACAGGTCA-3′ | |
| IL-10 | F | 5′-CTGCCTCCCACTTTCTCTTG-3′ |
| R | 5′-TCAAAGGGGCTCCCTAGTTT-3′ | |
| TGF-β | F | 5′-AGGGCTACCATGCCAATTTCT-3′ |
| R | 5′-CCGGGTTGTGCTGGTTGTACA-3′ | |
| SOD1 | F | 5′-ACCTGGGCAATGTGACTG-3′ |
| R | 5′-TCCAGCATTTCCCGTCT-3′ | |
| Catalase | F | 5′-AACTGTCCCTTCCGTGCTA-3′ |
| R | 5′-CCTGGGTGACATTATCTTCG-3′ | |
| GPX1 | F | 5′-CGGACCACCTGTTGAAAGCTC-3′ |
| R | 5′-TCCGCCAGTTCTTGTTGTCCA-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, Y.; Huang, X.; Li, L.; Liu, L.; Zhang, C.; Fan, X.; Xie, Y.; Zou, Y.; Geng, Z.; Huang, C. Effects of Solid Fermentation on Polygonatum cyrtonema Polysaccharides: Isolation, Characterization and Bioactivities. Molecules 2023, 28, 5498. https://doi.org/10.3390/molecules28145498
Cheng Y, Huang X, Li L, Liu L, Zhang C, Fan X, Xie Y, Zou Y, Geng Z, Huang C. Effects of Solid Fermentation on Polygonatum cyrtonema Polysaccharides: Isolation, Characterization and Bioactivities. Molecules. 2023; 28(14):5498. https://doi.org/10.3390/molecules28145498
Chicago/Turabian StyleCheng, Yi, Xueyuan Huang, Lixia Li, Lu Liu, Chunsheng Zhang, Xiang Fan, Yu Xie, Yuanfeng Zou, Zhe Geng, and Chao Huang. 2023. "Effects of Solid Fermentation on Polygonatum cyrtonema Polysaccharides: Isolation, Characterization and Bioactivities" Molecules 28, no. 14: 5498. https://doi.org/10.3390/molecules28145498