
Epigallocatechin-3-Gallate Therapeutic Potential in Cancer: Mechanism of Action and Clinical Implications
 ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Mechanism of Anticancer Activity of EGCG
2.1. Antioxidant Properties
2.2. Suppression of Inflammation
2.3. Modulation of Epigenetic Targets
2.4. Modulation of Cellular Signaling Molecules by EGCG
2.4.1. EGFR Pathway
2.4.2. JAK/STAT Pathway
2.4.3. MAPKs Pathways
2.4.4. PI3K/AKT/mTOR Pathway
2.4.5. Other Signaling Pathways
3. Emerging EGCG Targets
4. EGCG in Cancer Prevention and Therapy?
4.1. Epidemiological Studies
4.2. Clinical Studies
4.3. Bioavailability of EGCG
4.4. Other Concerns
- Standardization: Clinical research outcomes are difficult to evaluate and duplicate because of variations in the quality and purity of the EGCG preparations used.
- Safety concerns: Even though EGCG is usually believed to be safe, it might lead to liver damage in high doses, and there is a possibility that it could interact with some drugs. Additional research is required to completely understand the safety profile of this compound.
- Lack of clinical data: Although there is some evidence from preclinical research to show that EGCG may have a cancer-preventative effect, there is very little evidence from clinical studies that support its use in humans. More clinical tests that are carefully designed are required to assess the effectiveness and safety of this flavonoid.
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 4E-BP1 | Eukaryotic-translation-initiation-factor-4E-binding protein 1 |
| 67LR | 67-kD laminin receptor |
| ABL1 | Tyrosine-protein kinase ABL1 |
| AIF | Apoptosis-inducing factor |
| AKT | Protein kinase B |
| AP-1 | Activator protein-1 |
| APAF1 | Apoptotic-protease-activating factor 1 |
| APL | Acute promyelocytic leukemia |
| AR | Androgen receptor |
| ASAP | Atypical small acinar proliferation |
| ASM | Acid sphingomyelinase |
| BAX | Bcl-2-associated X protein |
| BCL-2 | B-cell lymphoma 2 |
| BCL-xL | B-cell lymphoma-extra large |
| BCR | B-cell receptor |
| CAFs | Cancer-associated fibroblasts |
| CD3 | Cluster of differentiation 3 |
| CDH1 | Cadherin 1 |
| CDK2 | Cyclin-dependent kinase 2 |
| CDK4/6 | Cyclin-dependent kinases 4/6 |
| c-IAP1/2 | Cellular inhibitor of apoptosis protein 1/2 |
| CLL | Chronic lymphocytic leukemia |
| COMT | Catechol-O-methyltransferase |
| COX-1 | Cyclooxygenase-1 |
| COX-2 | Cyclooxygenase-2 |
| COXs | Cyclooxygenases |
| CpG | Cytosine-phosphate-guanine |
| DAPK1 | Death-associated protein kinase 1 |
| DBGT | Double-brewed green tea |
| DNA-PK | DNA-dependent protein kinase |
| DNMT | DNA methyltransferase |
| DR5 | Death receptor 5 |
| XIAP | E3 ubiquitin-protein-ligase-X-linked inhibitor of apoptosis protein |
| EC | Epicatechin |
| ECG | Epicatechin-3-gallate |
| ECM | Extracellular matrix |
| EGC | Epigallocatechin |
| EGCG | Epigallocatechin-3-gallate |
| EGF | Epidermal growth factor |
| EGFR | Epidermal growth factor receptor |
| ER | Endoplasmic reticulum |
| ERK1/2 | Extracellular-signal-regulated kinases 1 and 2 |
| ETC | Electron transport chain |
| EZH2 | Enhancer of zeste homolog 2 |
| FAK | Focal adhesion kinase |
| FAS | Fatty acid synthase |
| FLIP | Cellular FLICE-like inhibitory protein |
| GADD153 | Growth arrest and DNA-damage-inducible gene 153 |
| GLI1 | GLI-Kruppel family member 1 |
| GLUTs | Glucose transporters |
| GRP78 | Glucose-regulated protein 78 |
| GST P1-1 | Glutathione S-transferase P1-1 |
| GTE | Green tea extract |
| HDAC | Histone deacetylase |
| HER-2 | Human epidermal growth factor receptor 2 |
| HGF | Hepatocyte growth factor |
| HGPIN | High-grade prostatic intraepithelial neoplasia |
| Hh | Hedgehog |
| HIF-1α | Hypoxia-inducible factor 1-alpha |
| HK2 | Hexokinase 2 |
| HUVECs | Human umbilical vein endothelial cells |
| ICAM-1 | Intercellular adhesion molecule-1 |
| IFNs | Interferons |
| IGF-1 | Insulin-like growth factor 1 |
| IGFBP-3 | IGF-binding protein-3 |
| IGF-1R | Insulin-like growth factor-1 receptor |
| IKKβ | Inhibitor of nuclear factor kappa-B kinase subunit beta |
| IL-1β | Interleukin 1β |
| IL-2 | Interleukin 2 |
| IL-4 | Interleukin 4 |
| IL-6 | Interleukin 6 |
| IL-8 | Interleukin 8 |
| iNOS | Inducible nitric oxide synthase |
| IRAK4 | Interleukin-1-receptor-associated kinase 4 |
| IRF1 | Interferon regulatory factor 1 |
| JAK | Tyrosine-protein kinase JAK |
| JNK | c-Jun N-terminal kinase |
| Ki-67 | Antigen Ki-67 |
| KRAS | GTPase KRas |
| MAPK8 | Mitogen-activated protein kinase 8 |
| MCP-1 | Monocyte chemoattractant protein-1 |
| MD | Molecular dynamics |
| MMPs | Matrix metalloproteinases |
| MRP | Multidrug-resistance-associated protein |
| mTOR | Serine/threonine-protein kinase mTOR |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| NF-Kb | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| NO | Nitric oxide |
| NOS | Nitric oxide synthase |
| NOTCH | Neurogenic locus notch homolog protein |
| NPs | Natural products |
| NSCLC | Non-small-cell lung cancer |
| NTRK1 | High-affinity nerve growth factor receptor |
| OATP1B1 | Organic anion-transporting polypeptide 1B1 |
| P300/CBP | CREB-binding protein/p300 |
| PARP | Poly (ADP-ribose) polymerase |
| PcG | Polycomb group |
| PDK1 | Phosphoinositide-dependent kinase-1 |
| PD-L1/2 | Programmed death-ligand 1/2 |
| PFK | Phosphofructokinase |
| PFKP | Platelet-type phosphofructokinase |
| PGE2 | Prostaglandin E2 |
| P-gp | P-glycoprotein |
| PI3K | Phosphatidylinositol-3-kinase |
| PIP2 | Phosphatidylinositol 4,5-bisphosphate |
| PIP3 | Phosphatidylinositol 3,4,5-triphosphate |
| PK | Pyruvate kinase |
| PKC | Protein kinase C |
| PKM2 | Pyruvate kinase M2 |
| PP2A | Protein phosphatase 2A |
| PPE | Green tea polyphenon E |
| PRC2 | Polycomb-repressive complex 2 |
| PSA | Prostate-specific antigen |
| PTEN | Phosphatidylinositol-3,4,5-trisphosphate 3-phosphatase and dual-specificity protein phosphatase |
| RARβ | Retinoic acid receptor beta |
| RAS/MAPK | RAS/mitogen-activated protein kinase |
| ROS | Reactive oxygen species |
| S6K | Ribosomal protein kinase S6 |
| SAM | S-adenosylmethionine |
| SHH | Sonic hedgehog |
| SIRT6 | Sirtuin-6 |
| SLC21A6 | Organic anion transporter |
| SMO | Smoothened |
| SPR | Surface plasmon resonance |
| STAT | Signal transducer and activator of transcription |
| SULTs | Sulphotransferases |
| SYK | Spleen tyrosine kinase |
| TAMs | Tumor-associated macrophages |
| TCR | T-cell receptor |
| TIMP-3 | Tissue inhibitor of matrix metalloproteinase-3 |
| TLR4 | Toll-like receptor 4 |
| TME | Tumor microenvironment |
| TNFR-1/2 | TNF receptor 1/2 |
| TNF-α | Tumor necrosis factor α |
| TP53 | Tumor protein p53 |
| UGTs | UDP-glucuronosyltransferases |
| VEGF | Vascular endothelial growth factor |
| VEGFR | Vascular endothelial growth factor receptor |
| WEE1 | Wee1-like protein kinase |
| ZAP-70 | Zeta-chain-associated protein kinase-70 |
| γ-CD | γ-cyclodextrin |
References
- Hayat, K.; Iqbal, H.; Malik, U.; Bilal, U.; Mushtaq, S. Tea and Its Consumption: Benefits and Risks. Crit. Rev. Food Sci. Nutr. 2015, 55, 939–954. [Google Scholar] [CrossRef]
- Khan, N.; Mukhtar, H. Tea and Health: Studies in Humans. Curr. Pharm. Des. 2013, 19, 6141–6147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakrawarti, L.; Agrawal, R.; Dang, S.; Gupta, S.; Gabrani, R. Therapeutic Effects of EGCG: A Patent Review. Expert Opin. Ther. Pat. 2016, 26, 907–916. [Google Scholar] [CrossRef]
- Alam, M.; Ali, S.; Ashraf, G.M.; Bilgrami, A.L.; Yadav, D.K.; Hassan, M.I. Epigallocatechin 3-Gallate: From Green Tea to Cancer Therapeutics. Food Chem. 2022, 379, 132135. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.D.; Yang, C.S. Cancer Chemopreventive Activity and Bioavailability of Tea and Tea Polyphenols. Mutat. Res. 2003, 523–524, 201–208. [Google Scholar] [CrossRef]
- Yang, C.S.; Lambert, J.D.; Ju, J.; Lu, G.; Sang, S. Tea and Cancer Prevention: Molecular Mechanisms and Human Relevance. Toxicol. Appl. Pharmacol. 2007, 224, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Mukhtar, H.; Ahmad, N. Tea Polyphenols: Prevention of Cancer and Optimizing Health. Am. J. Clin. Nutr. 2000, 71, 1698S–1702S, discussion 1703S–1704S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohishi, T.; Goto, S.; Monira, P.; Isemura, M.; Nakamura, Y. Anti-Inflammatory Action of Green Tea. Anti-Inflamm. Anti-Allergy Agents Med. Chem. 2016, 15, 74–90. [Google Scholar] [CrossRef] [PubMed]
- Oz, H.S. Chronic Inflammatory Diseases and Green Tea Polyphenols. Nutrients 2017, 9, 561. [Google Scholar] [CrossRef] [Green Version]
- Sah, J.F.; Balasubramanian, S.; Eckert, R.L.; Rorke, E.A. Epigallocatechin-3-Gallate Inhibits Epidermal Growth Factor Receptor Signaling Pathway. Evidence for Direct Inhibition of ERK1/2 and AKT Kinases. J. Biol. Chem. 2004, 279, 12755–12762. [Google Scholar] [CrossRef] [Green Version]
- Minnelli, C.; Cianfruglia, L.; Laudadio, E.; Mobbili, G.; Galeazzi, R.; Armeni, T. Effect of Epigallocatechin-3-Gallate on EGFR Signaling and Migration in Non-Small Cell Lung Cancer. Int. J. Mol. Sci. 2021, 22, 11833. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Deguchi, A.; Lim, J.T.E.; Moriwaki, H.; Kopelovich, L.; Weinstein, I.B. (−)-Epigallocatechin Gallate and Polyphenon E Inhibit Growth and Activation of the Epidermal Growth Factor Receptor and Human Epidermal Growth Factor Receptor-2 Signaling Pathways in Human Colon Cancer Cells. Clin. Cancer Res. 2005, 11, 2735–2746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, M.; Suzui, M.; Weinstein, I.B. Effects of Epigallocatechin-3-Gallate on Growth, Epidermal Growth Factor Receptor Signaling Pathways, Gene Expression, and Chemosensitivity in Human Head and Neck Squamous Cell Carcinoma Cell Lines. Clin. Cancer Res. 2001, 7, 4220–4229. [Google Scholar] [PubMed]
- Hamed, F.N.; McDonagh, A.J.G.; Almaghrabi, S.; Bakri, Y.; Messenger, A.G.; Tazi-Ahnini, R. Epigallocatechin-3 Gallate Inhibits STAT-1/JAK2/IRF-1/HLA-DR/HLA-B and Reduces CD8 MKG2D Lymphocytes of Alopecia Areata Patients. Int. J. Environ. Res. Public Health 2018, 15, 2882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravindran Menon, D.; Li, Y.; Yamauchi, T.; Osborne, D.G.; Vaddi, P.K.; Wempe, M.F.; Zhai, Z.; Fujita, M. EGCG Inhibits Tumor Growth in Melanoma by Targeting JAK-STAT Signaling and Its Downstream PD-L1/PD-L2-PD1 Axis in Tumors and Enhancing Cytotoxic T-Cell Responses. Pharmaceuticals 2021, 14, 1081. [Google Scholar] [CrossRef]
- Al-Maghrebi, M.; Alnajem, A.S.; Esmaeil, A. Epigallocatechin-3-Gallate Modulates Germ Cell Apoptosis through the SAFE/Nrf2 Signaling Pathway. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2020, 393, 663–671. [Google Scholar] [CrossRef]
- Barberino, R.S.; Santos, J.M.S.; Lins, T.L.B.G.; Menezes, V.G.; Monte, A.P.O.; Gouveia, B.B.; Palheta, R.C.; Matos, M.H.T. Epigallocatechin-3-Gallate (EGCG) Reduces Apoptosis of Preantral Follicles through the Phosphatidylinositol-3-Kinase/Protein Kinase B (PI3K/AKT) Signaling Pathway after in Vitro Culture of Sheep Ovarian Tissue. Theriogenology 2020, 155, 25–32. [Google Scholar] [CrossRef]
- Van Aller, G.S.; Carson, J.D.; Tang, W.; Peng, H.; Zhao, L.; Copeland, R.A.; Tummino, P.J.; Luo, L. Epigallocatechin Gallate (EGCG), a Major Component of Green Tea, Is a Dual Phosphoinositide-3-Kinase/MTOR Inhibitor. Biochem. Biophys. Res. Commun. 2011, 406, 194–199. [Google Scholar] [CrossRef]
- Tang, G.; Zhang, Z.; Qian, H.; Chen, J.; Wang, Y.; Chen, X.; Chen, B.; Chen, Y. (−)-Epigallocatechin-3-Gallate Inhibits Osteosarcoma Cell Invasiveness by Inhibiting the MEK/ERK Signaling Pathway in Human Osteosarcoma Cells. J. Environ. Pathol. Toxicol. Oncol. 2015, 34, 85–93. [Google Scholar] [CrossRef]
- Wei, R.; Wirkus, J.; Yang, Z.; Machuca, J.; Esparza, Y.; Mackenzie, G.G. EGCG Sensitizes Chemotherapeutic-Induced Cytotoxicity by Targeting the ERK Pathway in Multiple Cancer Cell Lines. Arch. Biochem. Biophys. 2020, 692, 108546. [Google Scholar] [CrossRef]
- Steed, K.L.; Jordan, H.R.; Tollefsbol, T.O. SAHA and EGCG Promote Apoptosis in Triple-Negative Breast Cancer Cells, Possibly Through the Modulation of CIAP2. Anticancer Res. 2020, 40, 9–26. [Google Scholar] [CrossRef]
- Gu, J.-J.; Qiao, K.-S.; Sun, P.; Chen, P.; Li, Q. Study of EGCG Induced Apoptosis in Lung Cancer Cells by Inhibiting PI3K/Akt Signaling Pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 4557–4563. [Google Scholar] [CrossRef]
- Liu, L.; Ju, Y.; Wang, J.; Zhou, R. Epigallocatechin-3-Gallate Promotes Apoptosis and Reversal of Multidrug Resistance in Esophageal Cancer Cells. Pathol. Res. Pract. 2017, 213, 1242–1250. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, R.; Ito, K.; Ishida, T.; Hamanoue, M.; Adachi, S.; Watanabe, T.; Sato, Y. Catechin, Green Tea Component, Causes Caspase-Independent Necrosis-like Cell Death in Chronic Myelogenous Leukemia. Cancer Sci. 2009, 100, 349–356. [Google Scholar] [CrossRef]
- Alam, M.; Ali, S.; Ahmed, S.; Elasbali, A.M.; Adnan, M.; Islam, A.; Hassan, M.I.; Yadav, D.K. Therapeutic Potential of Ursolic Acid in Cancer and Diabetic Neuropathy Diseases. Int. J. Mol. Sci. 2021, 22, 12162. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.; Ahmed, S.; Elasbali, A.M.; Adnan, M.; Alam, S.; Hassan, M.I.; Pasupuleti, V.R. Therapeutic Implications of Caffeic Acid in Cancer and Neurological Diseases. Front. Oncol. 2022, 12, 860508. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.S.; Robertson, A.A.B.; Cooper, M.A. Natural Product and Natural Product Derived Drugs in Clinical Trials. Nat. Prod. Rep. 2014, 31, 1612–1661. [Google Scholar] [CrossRef] [PubMed]
- Gordaliza, M. Natural Products as Leads to Anticancer Drugs. Clin. Transl. Oncol. 2007, 9, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Solowey, E.; Lichtenstein, M.; Sallon, S.; Paavilainen, H.; Solowey, E.; Lorberboum-Galski, H. Evaluating Medicinal Plants for Anticancer Activity. Sci. World J. 2014, 2014, 721402. [Google Scholar] [CrossRef] [Green Version]
- Kebebe, D.; Liu, Y.; Wu, Y.; Vilakhamxay, M.; Liu, Z.; Li, J. Tumor-Targeting Delivery of Herb-Based Drugs with Cell-Penetrating/Tumor-Targeting Peptide-Modified Nanocarriers. Int. J. Nanomed. 2018, 13, 1425–1442. [Google Scholar] [CrossRef] [Green Version]
- Hossain, R.; Jain, D.; Khan, R.A.; Islam, M.T.; Mubarak, M.S.; Mohammad Saikat, A.S. Natural-Derived Molecules as a Potential Adjuvant in Chemotherapy: Normal Cell Protectors and Cancer Cell Sensitizers. Anti-Cancer Agents Med. Chem. 2022, 22, 836–850. [Google Scholar] [CrossRef]
- Diederich, M. Natural Compound Inducers of Immunogenic Cell Death. Arch. Pharm. Res. 2019, 42, 629–645. [Google Scholar] [CrossRef]
- Luo, K.-W.; Chen, W.; Lung, W.-Y.; Wei, X.-Y.; Cheng, B.-H.; Cai, Z.-M.; Huang, W.-R. EGCG Inhibited Bladder Cancer SW780 Cell Proliferation and Migration Both in Vitro and in Vivo via Down-Regulation of NF-ΚB and MMP-9. J. Nutr. Biochem. 2017, 41, 56–64. [Google Scholar] [CrossRef]
- Luo, K.-W.; Zhu, X.-H.; Zhao, T.; Zhong, J.; Gao, H.-C.; Luo, X.-L.; Huang, W.-R. EGCG Enhanced the Anti-Tumor Effect of Doxorubicine in Bladder Cancer via NF-ΚB/MDM2/P53 Pathway. Front. Cell Dev. Biol. 2020, 8, 606123. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Li, J.; Kang, L.; Liu, X.; Luo, J.; Zhang, L.; Li, Y.; Cai, J. Epigallocatechin-3-Gallate Induces Autophagy-Related Apoptosis Associated with LC3B II and Beclin Expression of Bladder Cancer Cells. J. Food Biochem. 2021, 45, e13758. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-Y.; Chen, Y.-J.; Chang, W.-A.; Li, W.-M.; Ke, H.-L.; Wu, W.-J.; Kuo, P.-L. Effects of Epigallocatechin Gallate (EGCG) on Urinary Bladder Urothelial Carcinoma-Next-Generation Sequencing and Bioinformatics Approaches. Medicina 2019, 55, 768. [Google Scholar] [CrossRef] [Green Version]
- Philips, B.J.; Coyle, C.H.; Morrisroe, S.N.; Chancellor, M.B.; Yoshimura, N. Induction of Apoptosis in Human Bladder Cancer Cells by Green Tea Catechins. Biomed. Res. 2009, 30, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Luo, K.-W.; Lung, W.-Y.; Chun-Xie; Luo, X.-L.; Huang, W.-R. EGCG Inhibited Bladder Cancer T24 and 5637 Cell Proliferation and Migration via PI3K/AKT Pathway. Oncotarget 2018, 9, 12261–12272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, D.-S.; Wang, H.; Tan, S.-W.; Huang, Y.-H.; Tsai, C.-Y.; Yeh, M.-K.; Wu, C.-J. The Treatment of Bladder Cancer in a Mouse Model by Epigallocatechin-3-Gallate-Gold Nanoparticles. Biomaterials 2011, 32, 7633–7640. [Google Scholar] [CrossRef]
- Qin, J.; Xie, L.-P.; Zheng, X.-Y.; Wang, Y.-B.; Bai, Y.; Shen, H.-F.; Li, L.-C.; Dahiya, R. A Component of Green Tea, (−)-Epigallocatechin-3-Gallate, Promotes Apoptosis in T24 Human Bladder Cancer Cells via Modulation of the PI3K/Akt Pathway and Bcl-2 Family Proteins. Biochem. Biophys. Res. Commun. 2007, 354, 852–857. [Google Scholar] [CrossRef]
- Sun, X.; Song, J.; Li, E.; Geng, H.; Li, Y.; Yu, D.; Zhong, C. (-)-Epigallocatechin-3-gallate Inhibits Bladder Cancer Stem Cells via Suppression of Sonic Hedgehog Pathway. Oncol. Rep. 2019, 42, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Wang, Y.; Bai, Y.; Yang, K.; Mao, Q.; Lin, Y.; Kong, D.; Zheng, X.; Xie, L. Epigallocatechin-3-Gallate Inhibits Bladder Cancer Cell Invasion via Suppression of NF-ΚB-mediated Matrix Metalloproteinase-9 Expression. Mol. Med. Rep. 2012, 6, 1040–1044. [Google Scholar] [CrossRef] [Green Version]
- Piwowarczyk, L.; Stawny, M.; Mlynarczyk, D.T.; Muszalska-Kolos, I.; Goslinski, T.; Jelińska, A. Role of Curcumin and (-)-Epigallocatechin-3-O-Gallate in Bladder Cancer Treatment: A Review. Cancers 2020, 12, 1801. [Google Scholar] [CrossRef]
- Zan, L.; Chen, Q.; Zhang, L.; Li, X. Epigallocatechin Gallate (EGCG) Suppresses Growth and Tumorigenicity in Breast Cancer Cells by Downregulation of MiR-25. Bioengineered 2019, 10, 374–382. [Google Scholar] [CrossRef] [Green Version]
- Wei, R.; Mao, L.; Xu, P.; Zheng, X.; Hackman, R.M.; Mackenzie, G.G.; Wang, Y. Suppressing Glucose Metabolism with Epigallocatechin-3-Gallate (EGCG) Reduces Breast Cancer Cell Growth in Preclinical Models. Food Funct. 2018, 9, 5682–5696. [Google Scholar] [CrossRef]
- Braicu, C.; Gherman, C.D.; Irimie, A.; Berindan-Neagoe, I. Epigallocatechin-3-Gallate (EGCG) Inhibits Cell Proliferation and Migratory Behaviour of Triple Negative Breast Cancer Cells. J. Nanosci. Nanotechnol. 2013, 13, 632–637. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.-W.; Makey, K.L.; Tucker, K.B.; Chinchar, E.; Mao, X.; Pei, I.; Thomas, E.Y.; Miele, L. EGCG, a Major Green Tea Catechin Suppresses Breast Tumor Angiogenesis and Growth via Inhibiting the Activation of HIF-1α and NFκB, and VEGF Expression. Vasc. Cell 2013, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Thangapazham, R.L.; Passi, N.; Maheshwari, R.K. Green Tea Polyphenol and Epigallocatechin Gallate Induce Apoptosis and Inhibit Invasion in Human Breast Cancer Cells. Cancer Biol. Ther. 2007, 6, 1938–1943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Tian, H.; Ma, X.; Li, L. Epigallocatechin Gallate, the Main Ingredient of Green Tea Induces Apoptosis in Breast Cancer Cells. Front. Biosci. 2006, 11, 2428–2433. [Google Scholar] [CrossRef] [Green Version]
- Mittal, A.; Pate, M.S.; Wylie, R.C.; Tollefsbol, T.O.; Katiyar, S.K. EGCG Down-Regulates Telomerase in Human Breast Carcinoma MCF-7 Cells, Leading to Suppression of Cell Viability and Induction of Apoptosis. Int. J. Oncol. 2004, 24, 703–710. [Google Scholar] [CrossRef]
- Romano, A.; Martel, F. The Role of EGCG in Breast Cancer Prevention and Therapy. Mini Rev. Med. Chem. 2021, 21, 883–898. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Zhao, D.Y.; Elliott, S.; Zhao, W.; Curiel, T.J.; Beckman, B.S.; Burow, M.E. Epigallocatechin-3 Gallate Induces Growth Inhibition and Apoptosis in Human Breast Cancer Cells through Survivin Suppression. Int. J. Oncol. 2007, 31, 705–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, W.S.; Huh, S.W.; Bae, S.-M.; Lee, I.P.; Lee, J.M.; Namkoong, S.E.; Kim, C.K.; Sin, J.-I. A Major Constituent of Green Tea, EGCG, Inhibits the Growth of a Human Cervical Cancer Cell Line, CaSki Cells, through Apoptosis, G(1) Arrest, and Regulation of Gene Expression. DNA Cell Biol. 2003, 22, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Zou, C.; Liu, H.; Feugang, J.M.; Hao, Z.; Chow, H.-H.S.; Garcia, F. Green Tea Compound in Chemoprevention of Cervical Cancer. Int. J. Gynecol. Cancer 2010, 20, 617–624. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Singh, R.; Bhui, K.; Tyagi, S.; Mahmood, Z.; Shukla, Y. Tea Polyphenols Induce Apoptosis through Mitochondrial Pathway and by Inhibiting Nuclear Factor-KappaB and Akt Activation in Human Cervical Cancer Cells. Oncol. Res. 2011, 19, 245–257. [Google Scholar] [CrossRef]
- Sharma, C.; Nusri, Q.E.-A.; Begum, S.; Javed, E.; Rizvi, T.A.; Hussain, A. (−)-Epigallocatechin-3-Gallate Induces Apoptosis and Inhibits Invasion and Migration of Human Cervical Cancer Cells. Asian Pac. J. Cancer Prev. 2012, 13, 4815–4822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.A.; Hussain, A.; Sundaram, M.K.; Alalami, U.; Gunasekera, D.; Ramesh, L.; Hamza, A.; Quraishi, U. (−)-Epigallocatechin-3-Gallate Reverses the Expression of Various Tumor-Suppressor Genes by Inhibiting DNA Methyltransferases and Histone Deacetylases in Human Cervical Cancer Cells. Oncol. Rep. 2015, 33, 1976–1984. [Google Scholar] [CrossRef] [Green Version]
- Almatroodi, S.A.; Almatroudi, A.; Khan, A.A.; Alhumaydhi, F.A.; Alsahli, M.A.; Rahmani, A.H. Potential Therapeutic Targets of Epigallocatechin Gallate (EGCG), the Most Abundant Catechin in Green Tea, and Its Role in the Therapy of Various Types of Cancer. Molecules 2020, 25, 3146. [Google Scholar] [CrossRef]
- Panji, M.; Behmard, V.; Zare, Z.; Malekpour, M.; Nejadbiglari, H.; Yavari, S.; Nayerpour Dizaj, T.; Safaeian, A.; Maleki, N.; Abbasi, M.; et al. Suppressing Effects of Green Tea Extract and Epigallocatechin-3-Gallate (EGCG) on TGF-β- Induced Epithelial-to-Mesenchymal Transition via ROS/Smad Signaling in Human Cervical Cancer Cells. Gene 2021, 794, 145774. [Google Scholar] [CrossRef]
- La, X.; Zhang, L.; Li, Z.; Li, H.; Yang, Y. (−)-Epigallocatechin Gallate (EGCG) Enhances the Sensitivity of Colorectal Cancer Cells to 5-FU by Inhibiting GRP78/NF-ΚB/MiR-155-5p/MDR1 Pathway. J. Agric. Food Chem. 2019, 67, 2510–2518. [Google Scholar] [CrossRef]
- Wu, W.; Dong, J.; Gou, H.; Geng, R.; Yang, X.; Chen, D.; Xiang, B.; Zhang, Z.; Ren, S.; Chen, L.; et al. EGCG Synergizes the Therapeutic Effect of Irinotecan through Enhanced DNA Damage in Human Colorectal Cancer Cells. J. Cell Mol. Med. 2021, 25, 7913–7921. [Google Scholar] [CrossRef]
- Luo, K.-W.; Xia, J.; Cheng, B.-H.; Gao, H.-C.; Fu, L.-W.; Luo, X.-L. Tea Polyphenol EGCG Inhibited Colorectal-Cancer-Cell Proliferation and Migration via Downregulation of STAT3. Gastroenterol. Rep. 2021, 9, 59–70. [Google Scholar] [CrossRef]
- Wu, W.; Gou, H.; Xiang, B.; Geng, R.; Dong, J.; Yang, X.; Chen, D.; Dai, R.; Chen, L.; Liu, J. EGCG Enhances the Chemosensitivity of Colorectal Cancer to Irinotecan through GRP78-MediatedEndoplasmic Reticulum Stress. J. Oncol. 2022, 2022, 7099589. [Google Scholar] [CrossRef]
- Du, G.-J.; Zhang, Z.; Wen, X.-D.; Yu, C.; Calway, T.; Yuan, C.-S.; Wang, C.-Z. Epigallocatechin Gallate (EGCG) Is the Most Effective Cancer Chemopreventive Polyphenol in Green Tea. Nutrients 2012, 4, 1679–1691. [Google Scholar] [CrossRef]
- Jin, H.; Gong, W.; Zhang, C.; Wang, S. Epigallocatechin Gallate Inhibits the Proliferation of Colorectal Cancer Cells by Regulating Notch Signaling. OncoTargets Ther. 2013, 6, 145–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, J.-D.; Yao, J.-J.; Wang, H.; Cui, W.-G.; Leng, J.; Ding, L.-Y.; Fan, K.-Y. Effects of EGCG on Proliferation and Apoptosis of Gastric Cancer SGC7901 Cells via Down-Regulation of HIF-1α and VEGF under a Hypoxic State. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Du, W.; Yang, D. Inhibition of Green Tea Polyphenol EGCG((−)-Epigallocatechin-3-Gallate) on the Proliferation of Gastric Cancer Cells by Suppressing Canonical Wnt/β-Catenin Signalling Pathway. Int. J. Food Sci. Nutr. 2016, 67, 818–827. [Google Scholar] [CrossRef] [PubMed]
- Onoda, C.; Kuribayashi, K.; Nirasawa, S.; Tsuji, N.; Tanaka, M.; Kobayashi, D.; Watanabe, N. (−)-Epigallocatechin-3-Gallate Induces Apoptosis in Gastric Cancer Cell Lines by down-Regulating Survivin Expression. Int. J. Oncol. 2011, 38, 1403–1408. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.-H.; Zhan, W.-H.; Li, Z.-R.; Wang, Z.; He, Y.-L.; Peng, J.-S.; Cai, S.-R.; Ma, J.-P.; Zhang, C.-H. (−)-Epigallocatechin-3-Gallate Inhibits Growth of Gastric Cancer by Reducing VEGF Production and Angiogenesis. World J. Gastroenterol. 2007, 13, 1162–1169. [Google Scholar] [CrossRef] [Green Version]
- Sojoodi, M.; Wei, L.; Erstad, D.J.; Yamada, S.; Fujii, T.; Hirschfield, H.; Kim, R.S.; Lauwers, G.Y.; Lanuti, M.; Hoshida, Y.; et al. Epigallocatechin Gallate Induces Hepatic Stellate Cell Senescence and Attenuates Development of Hepatocellular Carcinoma. Cancer Prev. Res. 2020, 13, 497–508. [Google Scholar] [CrossRef] [Green Version]
- Bimonte, S.; Albino, V.; Piccirillo, M.; Nasto, A.; Molino, C.; Palaia, R.; Cascella, M. Epigallocatechin-3-Gallate in the Prevention and Treatment of Hepatocellular Carcinoma: Experimental Findings and Translational Perspectives. Drug Des. Dev. Ther. 2019, 13, 611–621. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, T.; Nakajima, T.; Moriguchi, M.; Jo, M.; Sekoguchi, S.; Ishii, M.; Takashima, H.; Katagishi, T.; Kimura, H.; Minami, M.; et al. A Green Tea Polyphenol, Epigalocatechin-3-Gallate, Induces Apoptosis of Human Hepatocellular Carcinoma, Possibly through Inhibition of Bcl-2 Family Proteins. J. Hepatol. 2006, 44, 1074–1082. [Google Scholar] [CrossRef]
- Bravi, F.; La Vecchia, C.; Turati, F. Green Tea and Liver Cancer. Hepatobiliary Surg. Nutr. 2017, 6, 127–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakamoto, Y.; Terashita, N.; Muraguchi, T.; Fukusato, T.; Kubota, S. Effects of Epigallocatechin-3-Gallate (EGCG) on A549 Lung Cancer Tumor Growth and Angiogenesis. Biosci. Biotechnol. Biochem. 2013, 77, 1799–1803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Bian, S.; Yang, C.S. Green Tea Polyphenol EGCG Suppresses Lung Cancer Cell Growth through Upregulating MiR-210 Expression Caused by Stabilizing HIF-1α. Carcinogenesis 2011, 32, 1881–1889. [Google Scholar] [CrossRef] [Green Version]
- Dhatwalia, S.K.; Kumar, M.; Dhawan, D.K. Role of EGCG in Containing the Progression of Lung Tumorigenesis—A Multistage Targeting Approach. Nutr. Cancer 2018, 70, 334–349. [Google Scholar] [CrossRef]
- Shin, Y.S.; Kang, S.U.; Park, J.K.; Kim, Y.E.; Kim, Y.S.; Baek, S.J.; Lee, S.-H.; Kim, C.-H. Anti-Cancer Effect of (−)-Epigallocatechin-3-Gallate (EGCG) in Head and Neck Cancer through Repression of Transactivation and Enhanced Degradation of β-Catenin. Phytomedicine 2016, 23, 1344–1355. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, S.K. Emerging Phytochemicals for the Prevention and Treatment of Head and Neck Cancer. Molecules 2016, 21, 1610. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Hao, M.-W.; Dong, K.; Lin, F.; Ren, J.-H.; Zhang, H.-Z. Apoptosis Induction Effects of EGCG in Laryngeal Squamous Cell Carcinoma Cells through Telomerase Repression. Arch. Pharm. Res. 2009, 32, 1263–1269. [Google Scholar] [CrossRef]
- Sarniak, A.; Lipińska, J.; Tytman, K.; Lipińska, S. Endogenous Mechanisms of Reactive Oxygen Species (ROS) Generation. Postepy Hig. Med. Dosw. 2016, 70, 1150–1165. [Google Scholar] [CrossRef]
- Sandalio, L.M.; Rodríguez-Serrano, M.; Romero-Puertas, M.C.; del Río, L.A. Role of Peroxisomes as a Source of Reactive Oxygen Species (ROS) Signaling Molecules. Subcell. Biochem. 2013, 69, 231–255. [Google Scholar] [CrossRef]
- Henle, E.S.; Linn, S. Formation, Prevention, and Repair of DNA Damage by Iron/Hydrogen Peroxide. J. Biol. Chem. 1997, 272, 19095–19098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schieber, M.; Chandel, N.S. ROS Function in Redox Signaling and Oxidative Stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Cadet, J.; Douki, T.; Ravanat, J.-L. Measurement of Oxidatively Generated Base Damage in Cellular DNA. Mutat. Res. 2011, 711, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Cadet, J.; Douki, T.; Ravanat, J.-L. Oxidatively Generated Base Damage to Cellular DNA. Free Radic. Biol. Med. 2010, 49, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Cadet, J.; Ravanat, J.-L.; TavernaPorro, M.; Menoni, H.; Angelov, D. Oxidatively Generated Complex DNA Damage: Tandem and Clustered Lesions. Cancer Lett. 2012, 327, 5–15. [Google Scholar] [CrossRef]
- Nita, M.; Grzybowski, A. The Role of the Reactive Oxygen Species and Oxidative Stress in the Pathomechanism of the Age-Related Ocular Diseases and Other Pathologies of the Anterior and Posterior Eye Segments in Adults. Oxid. Med. Cell Longev. 2016, 2016, 3164734. [Google Scholar] [CrossRef] [Green Version]
- Snezhkina, A.V.; Kudryavtseva, A.V.; Kardymon, O.L.; Savvateeva, M.V.; Melnikova, N.V.; Krasnov, G.S.; Dmitriev, A.A. ROS Generation and Antioxidant Defense Systems in Normal and Malignant Cells. Oxid. Med. Cell Longev. 2019, 2019, 6175804. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, N.; Walker, G.C. Mechanisms of DNA Damage, Repair, and Mutagenesis. Environ. Mol. Mutagen. 2017, 58, 235–263. [Google Scholar] [CrossRef] [Green Version]
- Ljungman, M.; Hanawalt, P.C. Efficient Protection against Oxidative DNA Damage in Chromatin. Mol. Carcinog. 1992, 5, 264–269. [Google Scholar] [CrossRef]
- Kreuz, S.; Fischle, W. Oxidative Stress Signaling to Chromatin in Health and Disease. Epigenomics 2016, 8, 843–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakubczyk, K.; Dec, K.; Kałduńska, J.; Kawczuga, D.; Kochman, J.; Janda, K. Reactive Oxygen Species—Sources, Functions, Oxidative Damage. Pol. Merkur. Lekarski 2020, 48, 124–127. [Google Scholar] [PubMed]
- Bergamini, C.M.; Gambetti, S.; Dondi, A.; Cervellati, C. Oxygen, Reactive Oxygen Species and Tissue Damage. Curr. Pharm. Des. 2004, 10, 1611–1626. [Google Scholar] [CrossRef] [PubMed]
- Mokra, D.; Adamcakova, J.; Mokry, J. Green Tea Polyphenol (−)-Epigallocatechin-3-Gallate (EGCG): A Time for a New Player in the Treatment of Respiratory Diseases? Antioxidants 2022, 11, 1566. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Adhami, V.M.; Mukhtar, H. Review: Green Tea Polyphenols in Chemoprevention of Prostate Cancer: Preclinical and Clinical Studies. Nutr. Cancer 2009, 61, 836–841. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-S.; Quon, M.J.; Kim, J. New Insights into the Mechanisms of Polyphenols beyond Antioxidant Properties; Lessons from the Green Tea Polyphenol, Epigallocatechin 3-Gallate. Redox Biol. 2014, 2, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Legeay, S.; Rodier, M.; Fillon, L.; Faure, S.; Clere, N. Epigallocatechin Gallate: A Review of Its Beneficial Properties to Prevent Metabolic Syndrome. Nutrients 2015, 7, 5443. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Li, Y.; Li, Z.; Ma, C.; Lou, Z.; Yokoyama, W.; Wang, H. Lipase-Catalyzed Synthesis of Acetylated EGCG and Antioxidant Properties of the Acetylated Derivatives. Food Res. Int. 2014, 56, 279–286. [Google Scholar] [CrossRef]
- Greten, F.R.; Grivennikov, S.I. Inflammation and Cancer: Triggers, Mechanisms and Consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef]
- Murata, M. Inflammation and Cancer. Environ. Health Prev. Med. 2018, 23, 50. [Google Scholar] [CrossRef] [Green Version]
- Coussens, L.M.; Werb, Z. Inflammation and Cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Kciuk, M.; Kołat, D.; Kałuzińska-Kołat, Ż.; Gawrysiak, M.; Drozda, R.; Celik, I.; Kontek, R. PD-1/PD-L1 and DNA Damage Response in Cancer. Cells 2023, 12, 530. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lin, Y. Tumor Necrosis Factor and Cancer, Buddies or Foes? Acta Pharmacol. Sin. 2008, 29, 1275–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Horssen, R.; Ten Hagen, T.L.M.; Eggermont, A.M.M. TNF-Alpha in Cancer Treatment: Molecular Insights, Antitumor Effects, and Clinical Utility. Oncologist 2006, 11, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Wajant, H. The Role of TNF in Cancer. Results Probl. Cell Differ. 2009, 49, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Balkwill, F. TNF-Alpha in Promotion and Progression of Cancer. Cancer Metastasis Rev. 2006, 25, 409–416. [Google Scholar] [CrossRef]
- Shen, J.; Xiao, Z.; Zhao, Q.; Li, M.; Wu, X.; Zhang, L.; Hu, W.; Cho, C.H. Anti-Cancer Therapy with TNFα and IFNγ: A Comprehensive Review. Cell Prolif. 2018, 51, e12441. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.-M.; Gao, W.; Wang, H.; Zhao, D.; Nie, Z.-L.; Shi, J.-Q.; Zhao, S.; Lu, X.; Wang, L.-S.; Yang, Z.-J. Green Tea Polyphenol Epigallocatechin-3-Gallate Inhibits TNF-α-Induced Production of Monocyte Chemoattractant Protein-1 in Human Umbilical Vein Endothelial Cells. Cell Physiol. Biochem. 2014, 33, 1349–1358. [Google Scholar] [CrossRef]
- Shin, H.-Y.; Kim, S.-H.; Jeong, H.-J.; Kim, S.-Y.; Shin, T.-Y.; Um, J.-Y.; Hong, S.-H.; Kim, H.-M. Epigallocatechin-3-Gallate Inhibits Secretion of TNF-Alpha, IL-6 and IL-8 through the Attenuation of ERK and NF-KappaB in HMC-1 Cells. Int. Arch. Allergy Immunol. 2007, 142, 335–344. [Google Scholar] [CrossRef]
- Pang, L.Y.; Hurst, E.A.; Argyle, D.J. Cyclooxygenase-2: A Role in Cancer Stem Cell Survival and Repopulation of Cancer Cells during Therapy. Stem Cells Int. 2016, 2016, 2048731. [Google Scholar] [CrossRef] [Green Version]
- Hashemi Goradel, N.; Najafi, M.; Salehi, E.; Farhood, B.; Mortezaee, K. Cyclooxygenase-2 in Cancer: A Review. J. Cell Physiol. 2019, 234, 5683–5699. [Google Scholar] [CrossRef] [PubMed]
- Sinicrope, F.A. Targeting Cyclooxygenase-2 for Prevention and Therapy of Colorectal Cancer. Mol. Carcinog. 2006, 45, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.-C. COX-2 Inhibitors in Cancer Treatment and Prevention, a Recent Development. Anti-Cancer Drugs 2002, 13, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Arun, B.; Goss, P. The Role of COX-2 Inhibition in Breast Cancer Treatment and Prevention. Semin. Oncol. 2004, 31, 22–29. [Google Scholar] [CrossRef]
- Sinicrope, F.A.; Gill, S. Role of Cyclooxygenase-2 in Colorectal Cancer. Cancer Metastasis Rev. 2004, 23, 63–75. [Google Scholar] [CrossRef]
- Peng, G.; Dixon, D.A.; Muga, S.J.; Smith, T.J.; Wargovich, M.J. Green Tea Polyphenol (−)-Epigallocatechin-3-Gallate Inhibits Cyclooxygenase-2 Expression in Colon Carcinogenesis. Mol. Carcinog. 2006, 45, 309–319. [Google Scholar] [CrossRef]
- Hussain, T.; Gupta, S.; Adhami, V.M.; Mukhtar, H. Green Tea Constituent Epigallocatechin-3-Gallate Selectively Inhibits COX-2 without Affecting COX-1 Expression in Human Prostate Carcinoma Cells. Int. J. Cancer 2005, 113, 660–669. [Google Scholar] [CrossRef]
- Adhami, V.M.; Malik, A.; Zaman, N.; Sarfaraz, S.; Siddiqui, I.A.; Syed, D.N.; Afaq, F.; Pasha, F.S.; Saleem, M.; Mukhtar, H. Combined Inhibitory Effects of Green Tea Polyphenols and Selective Cyclooxygenase-2 Inhibitors on the Growth of Human Prostate Cancer Cells Both in Vitro and in Vivo. Clin. Cancer Res. 2007, 13, 1611–1619. [Google Scholar] [CrossRef] [Green Version]
- Harper, C.E.; Patel, B.B.; Wang, J.; Eltoum, I.A.; Lamartiniere, C.A. Epigallocatechin-3-Gallate Suppresses Early Stage, but Not Late Stage Prostate Cancer in TRAMP Mice: Mechanisms of Action. Prostate 2007, 67, 1576–1589. [Google Scholar] [CrossRef]
- Xu, W.; Liu, L.Z.; Loizidou, M.; Ahmed, M.; Charles, I.G. The Role of Nitric Oxide in Cancer. Cell Res. 2002, 12, 311–320. [Google Scholar] [CrossRef] [Green Version]
- Khan, F.H.; Dervan, E.; Bhattacharyya, D.D.; McAuliffe, J.D.; Miranda, K.M.; Glynn, S.A. The Role of Nitric Oxide in Cancer: Master Regulator or NOt? Int. J. Mol. Sci. 2020, 21, 9393. [Google Scholar] [CrossRef] [PubMed]
- Somasundaram, V.; Basudhar, D.; Bharadwaj, G.; No, J.H.; Ridnour, L.A.; Cheng, R.Y.S.; Fujita, M.; Thomas, D.D.; Anderson, S.K.; McVicar, D.W.; et al. Molecular Mechanisms of Nitric Oxide in Cancer Progression, Signal Transduction, and Metabolism. Antioxid. Redox Signal. 2019, 30, 1124–1143. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.M.; Fong, D.; Ho, C.T.; Huang, H.I. Inhibition of Inducible Nitric Oxide Synthase Gene Expression and Enzyme Activity by Epigallocatechin Gallate, a Natural Product from Green Tea. Biochem. Pharmacol. 1997, 54, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.L.; Lin, J.K. (−)-Epigallocatechin-3-Gallate Blocks the Induction of Nitric Oxide Synthase by down-Regulating Lipopolysaccharide-Induced Activity of Transcription Factor Nuclear Factor-KappaB. Mol. Pharmacol. 1997, 52, 465–472. [Google Scholar]
- Xia, L.; Tan, S.; Zhou, Y.; Lin, J.; Wang, H.; Oyang, L.; Tian, Y.; Liu, L.; Su, M.; Wang, H.; et al. Role of the NFκB-Signaling Pathway in Cancer. OncoTargets Ther. 2018, 11, 2063–2073. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Shen, S.; Verma, I.M. NF-ΚB, an Active Player in Human Cancers. Cancer Immunol. Res. 2014, 2, 823. [Google Scholar] [CrossRef] [Green Version]
- Itoh, Y.; Nagase, H. Matrix Metalloproteinases in Cancer. Essays Biochem. 2002, 38, 21–36. [Google Scholar] [CrossRef] [Green Version]
- Gialeli, C.; Theocharis, A.D.; Karamanos, N.K. Roles of Matrix Metalloproteinases in Cancer Progression and Their Pharmacological Targeting. FEBS J. 2011, 278, 16–27. [Google Scholar] [CrossRef]
- Mustafa, S.; Koran, S.; AlOmair, L. Insights Into the Role of Matrix Metalloproteinases in Cancer and Its Various Therapeutic Aspects: A Review. Front. Mol. Biosci. 2022, 9, 896099. [Google Scholar] [CrossRef]
- Chowdhury, A.; Nandy, S.K.; Sarkar, J.; Chakraborti, T.; Chakraborti, S. Inhibition of Pro-/Active MMP-2 by Green Tea Catechins and Prediction of Their Interaction by Molecular Docking Studies. Mol. Cell Biochem. 2017, 427, 111–122. [Google Scholar] [CrossRef]
- Sarkar, J.; Nandy, S.K.; Chowdhury, A.; Chakraborti, T.; Chakraborti, S. Inhibition of MMP-9 by Green Tea Catechins and Prediction of Their Interaction by Molecular Docking Analysis. Biomed. Pharmacother. 2016, 84, 340–347. [Google Scholar] [CrossRef]
- Maeda-Yamamoto, M.; Kawahara, H.; Tahara, N.; Tsuji, K.; Hara, Y.; Isemura, M. Effects of Tea Polyphenols on the Invasion and Matrix Metalloproteinases Activities of Human Fibrosarcoma HT1080 Cells. J. Agric. Food Chem. 1999, 47, 2350–2354. [Google Scholar] [CrossRef]
- Ilango, S.; Paital, B.; Jayachandran, P.; Padma, P.R.; Nirmaladevi, R. Epigenetic Alterations in Cancer. Front. Biosci. 2020, 25, 1058–1109. [Google Scholar] [CrossRef]
- Kanwal, R.; Gupta, S. Epigenetic Modifications in Cancer. Clin. Genet. 2012, 81, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Miranda Furtado, C.L.; Dos Santos Luciano, M.C.; Silva Santos, R.D.; Furtado, G.P.; Moraes, M.O.; Pessoa, C. Epidrugs: Targeting Epigenetic Marks in Cancer Treatment. Epigenetics 2019, 14, 1164–1176. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, C.; Wu, C.; Cui, W.; Wang, L. DNA Methyltransferases in Cancer: Biology, Paradox, Aberrations, and Targeted Therapy. Cancers 2020, 12, 2123. [Google Scholar] [CrossRef] [PubMed]
- Peng, D.-F.; Kanai, Y.; Sawada, M.; Ushijima, S.; Hiraoka, N.; Kitazawa, S.; Hirohashi, S. DNA Methylation of Multiple Tumor-Related Genes in Association with Overexpression of DNA Methyltransferase 1 (DNMT1) during Multistage Carcinogenesis of the Pancreas. Carcinogenesis 2006, 27, 1160–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, Y.; Kanai, Y.; Nakagawa, T.; Sakamoto, M.; Saito, H.; Ishii, H.; Hirohashi, S. Increased Protein Expression of DNA Methyltransferase (DNMT) 1 Is Significantly Correlated with the Malignant Potential and Poor Prognosis of Human Hepatocellular Carcinomas. Int. J. Cancer 2003, 105, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Butcher, D.T.; Rodenhiser, D.I. Epigenetic Inactivation of BRCA1 Is Associated with Aberrant Expression of CTCF and DNA Methyltransferase (DNMT3B) in Some Sporadic Breast Tumours. Eur. J. Cancer 2007, 43, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Linhart, H.G.; Lin, H.; Yamada, Y.; Moran, E.; Steine, E.J.; Gokhale, S.; Lo, G.; Cantu, E.; Ehrich, M.; He, T.; et al. Dnmt3b Promotes Tumorigenesis in Vivo by Gene-Specific de Novo Methylation and Transcriptional Silencing. Genes. Dev. 2007, 21, 3110–3122. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Xu, J. DNA Methyltransferases and Their Roles in Tumorigenesis. Biomark. Res. 2017, 5, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.-H.; Gudas, L.J. Retinoids, Retinoic Acid Receptors, and Cancer. Annu. Rev. Pathol. 2011, 6, 345–364. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, K.; Utsunomiya, H.; Miki, Y.; Hanihara, M.; Fue, M.; Takagi, K.; Nishimoto, M.; Suzuki, F.; Yaegashi, N.; Suzuki, T.; et al. Retinoic Acid Receptor β: A Potential Therapeutic Target in Retinoic Acid Treatment of Endometrial Cancer. Int. J. Gynecol. Cancer 2017, 27, 643–650. [Google Scholar] [CrossRef]
- Berx, G.; van Roy, F. Involvement of Members of the Cadherin Superfamily in Cancer. Cold Spring Harb. Perspect. Biol. 2009, 1, a003129. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Zhou, X.Z.; Lee, T.H. Death-Associated Protein Kinase 1 as a Promising Drug Target in Cancer and Alzheimer’s Disease. Recent Pat. Anti-Cancer Drug Discov. 2019, 14, 144–157. [Google Scholar] [CrossRef]
- Nandakumar, V.; Vaid, M.; Katiyar, S.K. (−)-Epigallocatechin-3-Gallate Reactivates Silenced Tumor Suppressor Genes, Cip1/P21 and P16INK4a, by Reducing DNA Methylation and Increasing Histones Acetylation in Human Skin Cancer Cells. Carcinogenesis 2011, 32, 537–544. [Google Scholar] [CrossRef] [Green Version]
- Barneda-Zahonero, B.; Parra, M. Histone Deacetylases and Cancer. Mol. Oncol. 2012, 6, 579–589. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.-H.; Kwak, J.; Choi, H.-K.; Choi, K.-C.; Kim, S.; Lee, J.; Jun, W.; Park, H.-J.; Yoon, H.-G. EGCG Suppresses Prostate Cancer Cell Growth Modulating Acetylation of Androgen Receptor by Anti-Histone Acetyltransferase Activity. Int. J. Mol. Med. 2012, 30, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.-C.; Jung, M.G.; Lee, Y.-H.; Yoon, J.C.; Kwon, S.H.; Kang, H.-B.; Kim, M.-J.; Cha, J.-H.; Kim, Y.J.; Jun, W.J.; et al. Epigallocatechin-3-Gallate, a Histone Acetyltransferase Inhibitor, Inhibits EBV-Induced B Lymphocyte Transformation via Suppression of RelA Acetylation. Cancer Res. 2009, 69, 583–592. [Google Scholar] [CrossRef] [Green Version]
- Borutinskaitė, V.; Virkšaitė, A.; Gudelytė, G.; Navakauskienė, R. Green Tea Polyphenol EGCG Causes Anti-Cancerous Epigenetic Modulations in Acute Promyelocytic Leukemia Cells. Leuk. Lymphoma 2018, 59, 469–478. [Google Scholar] [CrossRef]
- Deb, G.; Thakur, V.S.; Limaye, A.M.; Gupta, S. Epigenetic Induction of Tissue Inhibitor of Matrix Metalloproteinase-3 by Green Tea Polyphenols in Breast Cancer Cells. Mol. Carcinog. 2015, 54, 485–499. [Google Scholar] [CrossRef]
- Deb, G.; Shankar, E.; Thakur, V.S.; Ponsky, L.E.; Bodner, D.R.; Fu, P.; Gupta, S. Green Tea-Induced Epigenetic Reactivation of Tissue Inhibitor of Matrix Metalloproteinase-3 Suppresses Prostate Cancer Progression through Histone-Modifying Enzymes. Mol. Carcinog. 2019, 58, 1194–1207. [Google Scholar] [CrossRef] [PubMed]
- Oya, Y.; Mondal, A.; Rawangkan, A.; Umsumarng, S.; Iida, K.; Watanabe, T.; Kanno, M.; Suzuki, K.; Li, Z.; Kagechika, H.; et al. Down-Regulation of Histone Deacetylase 4, -5 and -6 as a Mechanism of Synergistic Enhancement of Apoptosis in Human Lung Cancer Cells Treated with the Combination of a Synthetic Retinoid, Am80 and Green Tea Catechin. J. Nutr. Biochem. 2017, 42, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, M.A.; Schlessinger, J. Cell Signaling by Receptor Tyrosine Kinases. Cell 2010, 141, 1117–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlessinger, J. Receptor Tyrosine Kinases: Legacy of the First Two Decades. Cold Spring Harb. Perspect. Biol. 2014, 6, a008912. [Google Scholar] [CrossRef] [Green Version]
- Sigismund, S.; Avanzato, D.; Lanzetti, L. Emerging Functions of the EGFR in Cancer. Mol. Oncol. 2018, 12, 3–20. [Google Scholar] [CrossRef] [Green Version]
- Tan, X.; Lambert, P.F.; Rapraeger, A.C.; Anderson, R.A. Stress-Induced EGFR Trafficking: Mechanisms, Functions, and Therapeutic Implications. Trends Cell Biol. 2016, 26, 352–366. [Google Scholar] [CrossRef] [Green Version]
- Roskoski, R. The ErbB/HER Family of Protein-Tyrosine Kinases and Cancer. Pharmacol. Res. 2014, 79, 34–74. [Google Scholar] [CrossRef]
- Wilson, K.J.; Mill, C.; Lambert, S.; Buchman, J.; Wilson, T.R.; Hernandez-Gordillo, V.; Gallo, R.M.; Ades, L.M.C.; Settleman, J.; Riese, D.J. EGFR Ligands Exhibit Functional Differences in Models of Paracrine and Autocrine Signaling. Growth Factors 2012, 30, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, K.M.; Berger, M.B.; Mendrola, J.M.; Cho, H.S.; Leahy, D.J.; Lemmon, M.A. EGF Activates Its Receptor by Removing Interactions That Autoinhibit Ectodomain Dimerization. Mol. Cell 2003, 11, 507–517. [Google Scholar] [CrossRef]
- Singh, B.; Carpenter, G.; Coffey, R.J. EGF Receptor Ligands: Recent Advances. F1000Research 2016, 5, F1000 Faculty Rev-2270. [Google Scholar] [CrossRef] [Green Version]
- Yarden, Y.; Sliwkowski, M.X. Untangling the ErbB Signalling Network. Nat. Rev. Mol. Cell Biol. 2001, 2, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, M.A.; Schlessinger, J.; Ferguson, K.M. The EGFR Family: Not so Prototypical Receptor Tyrosine Kinases. Cold Spring Harb. Perspect. Biol. 2014, 6, a020768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freed, D.M.; Bessman, N.J.; Kiyatkin, A.; Salazar-Cavazos, E.; Byrne, P.O.; Moore, J.O.; Valley, C.C.; Ferguson, K.M.; Leahy, D.J.; Lidke, D.S.; et al. EGFR Ligands Differentially Stabilize Receptor Dimers to Specify Signaling Kinetics. Cell 2017, 171, 683–695.e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, R.; Benedetti, L.G.; Abera, M.B.; Wang, H.; Abba, M.; Kazanietz, M.G. Protein Kinase C and Cancer: What We Know and What We Do Not. Oncogene 2014, 33, 5225–5237. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Li, Q.; Huang, Q.; Cheng, J. Targeting Protein Kinase C for Cancer Therapy. Cancers 2022, 14, 1104. [Google Scholar] [CrossRef]
- Sun, X.-L.; Xiang, Z.-M.; Xie, Y.-R.; Zhang, N.; Wang, L.-X.; Wu, Y.-L.; Zhang, D.-Y.; Wang, X.-J.; Sheng, J.; Zi, C.-T. Dimeric-(−)-Epigallocatechin-3-Gallate Inhibits the Proliferation of Lung Cancer Cells by Inhibiting the EGFR Signaling Pathway. Chem. Biol. Interact. 2022, 365, 110084. [Google Scholar] [CrossRef]
- Honda, Y.; Takigawa, N.; Ichihara, E.; Ninomiya, T.; Kubo, T.; Ochi, N.; Yasugi, M.; Murakami, T.; Yamane, H.; Tanimoto, M.; et al. Effects of (−)-Epigallocatechin-3-Gallate on EGFR- or Fusion Gene-Driven Lung Cancer Cells. Acta Med. Okayama 2017, 71, 505–512. [Google Scholar] [CrossRef]
- Wu, D.; Liu, Z.; Li, J.; Zhang, Q.; Zhong, P.; Teng, T.; Chen, M.; Xie, Z.; Ji, A.; Li, Y. Epigallocatechin-3-Gallate Inhibits the Growth and Increases the Apoptosis of Human Thyroid Carcinoma Cells through Suppression of EGFR/RAS/RAF/MEK/ERK Signaling Pathway. Cancer Cell Int. 2019, 19, 43. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Chen, L.; Lu, T.; Xie, Y.; Li, C.; Jia, Z.; Cao, J. ERα36 Is an Effective Target of Epigallocatechin-3-Gallate in Hepatocellular Carcinoma. Int. J. Clin. Exp. Pathol. 2019, 12, 3222–3234. [Google Scholar]
- Filippi, A.; Picot, T.; Aanei, C.M.; Nagy, P.; Szöllősi, J.; Campos, L.; Ganea, C.; Mocanu, M.-M. Epigallocatechin-3-O-Gallate Alleviates the Malignant Phenotype in A-431 Epidermoid and SK-BR-3 Breast Cancer Cell Lines. Int. J. Food Sci. Nutr. 2018, 69, 584–597. [Google Scholar] [CrossRef] [PubMed]
- Weng, L.X.; Wang, G.H.; Yao, H.; Yu, M.F.; Lin, J. Epigallocatechin Gallate Inhibits the Growth of Salivary Adenoid Cystic Carcinoma Cells via the EGFR/Erk Signal Transduction Pathway and the Mitochondria Apoptosis Pathway. Neoplasma 2017, 64, 563–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosseini, A.; Gharibi, T.; Marofi, F.; Javadian, M.; Babaloo, Z.; Baradaran, B. Janus Kinase Inhibitors: A Therapeutic Strategy for Cancer and Autoimmune Diseases. J. Cell Physiol. 2020, 235, 5903–5924. [Google Scholar] [CrossRef] [PubMed]
- Thoidingjam, L.K.; Blouin, C.M.; Gaillet, C.; Brion, A.; Solier, S.; Niyomchon, S.; El Marjou, A.; Mouasni, S.; Sepulveda, F.E.; de Saint Basile, G.; et al. Small Molecule Inhibitors of Interferon-Induced JAK-STAT Signalling. Angew. Chem. Int. Ed. Engl. 2022, 61, e202205231. [Google Scholar] [CrossRef]
- Hu, X.; Li, J.; Fu, M.; Zhao, X.; Wang, W. The JAK/STAT Signaling Pathway: From Bench to Clinic. Signal Transduct. Target. Ther. 2021, 6, 402. [Google Scholar] [CrossRef]
- Groner, B.; von Manstein, V. Jak Stat Signaling and Cancer: Opportunities, Benefits and Side Effects of Targeted Inhibition. Mol. Cell Endocrinol. 2017, 451, 1–14. [Google Scholar] [CrossRef]
- Ou, A.; Ott, M.; Fang, D.; Heimberger, A.B. The Role and Therapeutic Targeting of JAK/STAT Signaling in Glioblastoma. Cancers 2021, 13, 437. [Google Scholar] [CrossRef]
- Rah, B.; Rather, R.A.; Bhat, G.R.; Baba, A.B.; Mushtaq, I.; Farooq, M.; Yousuf, T.; Dar, S.B.; Parveen, S.; Hassan, R.; et al. JAK/STAT Signaling: Molecular Targets, Therapeutic Opportunities, and Limitations of Targeted Inhibitions in Solid Malignancies. Front. Pharmacol. 2022, 13, 821344. [Google Scholar] [CrossRef]
- Shao, F.; Pang, X.; Baeg, G.H. Targeting the JAK/STAT Signaling Pathway for Breast Cancer. Curr. Med. Chem. 2021, 28, 5137–5151. [Google Scholar] [CrossRef]
- Tang, S.-N.; Fu, J.; Shankar, S.; Srivastava, R.K. EGCG Enhances the Therapeutic Potential of Gemcitabine and CP690550 by Inhibiting STAT3 Signaling Pathway in Human Pancreatic Cancer. PLoS ONE 2012, 7, e31067. [Google Scholar] [CrossRef] [Green Version]
- Masuda, M.; Suzui, M.; Lim, J.T.E.; Weinstein, I.B. Epigallocatechin-3-Gallate Inhibits Activation of HER-2/Neu and Downstream Signaling Pathways in Human Head and Neck and Breast Carcinoma Cells. Clin. Cancer Res. 2003, 9, 3486–3491. [Google Scholar] [PubMed]
- Xiao, X.; Jiang, K.; Xu, Y.; Peng, H.; Wang, Z.; Liu, S.; Zhang, G. (−)-Epigallocatechin-3-Gallate Induces Cell Apoptosis in Chronic Myeloid Leukaemia by Regulating Bcr/Abl-Mediated P38-MAPK/JNK and JAK2/STAT3/AKT Signalling Pathways. Clin. Exp. Pharmacol. Physiol. 2019, 46, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Senggunprai, L.; Kukongviriyapan, V.; Prawan, A.; Kukongviriyapan, U. Quercetin and EGCG Exhibit Chemopreventive Effects in Cholangiocarcinoma Cells via Suppression of JAK/STAT Signaling Pathway. Phytother. Res. 2014, 28, 841–848. [Google Scholar] [CrossRef]
- Wang, Y.; Ren, X.; Deng, C.; Yang, L.; Yan, E.; Guo, T.; Li, Y.; Xu, M.X. Mechanism of the Inhibition of the STAT3 Signaling Pathway by EGCG. Oncol. Rep. 2013, 30, 2691–2696. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.-W.; Shieh, P.-C.; Lin, Y.-C.; Chen, Y.-J.; Lin, Y.-H.; Kuo, D.-H.; Liu, J.-Y.; Kao, J.-Y.; Kao, M.-C.; Way, T.-D. Indoleamine 2,3-Dioxygenase, an Immunomodulatory Protein, Is Suppressed by (−)-Epigallocatechin-3-Gallate via Blocking of Gamma-Interferon-Induced JAK-PKC-Delta-STAT1 Signaling in Human Oral Cancer Cells. J. Agric. Food Chem. 2010, 58, 887–894. [Google Scholar] [CrossRef]
- Yuan, C.-H.; Horng, C.-T.; Lee, C.-F.; Chiang, N.-N.; Tsai, F.-J.; Lu, C.-C.; Chiang, J.-H.; Hsu, Y.-M.; Yang, J.-S.; Chen, F.-A. Epigallocatechin Gallate Sensitizes Cisplatin-Resistant Oral Cancer CAR Cell Apoptosis and Autophagy through Stimulating AKT/STAT3 Pathway and Suppressing Multidrug Resistance 1 Signaling. Environ. Toxicol. 2017, 32, 845–855. [Google Scholar] [CrossRef]
- Chang, L.; Karin, M. Mammalian MAP Kinase Signalling Cascades. Nature 2001, 410, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Hepworth, E.M.W.; Hinton, S.D. Pseudophosphatases as Regulators of MAPK Signaling. Int. J. Mol. Sci. 2021, 22, 12595. [Google Scholar] [CrossRef]
- Santarpia, L.; Lippman, S.M.; El-Naggar, A.K. Targeting the MAPK-RAS-RAF Signaling Pathway in Cancer Therapy. Expert. Opin. Ther. Targets 2012, 16, 103–119. [Google Scholar] [CrossRef] [Green Version]
- Guan, Y.; Wu, Q.; Li, M.; Chen, D.; Su, J.; Zuo, L.; Zhu, B.; Li, Y. Epigallocatechin-3-Gallate Induced HepG2 Cells Apoptosis through ROS-Mediated AKT /JNK and P53 Signaling Pathway. Curr. Cancer Drug Targets 2022, 23, 447–460. [Google Scholar] [CrossRef]
- Sah, D.K.; Khoi, P.N.; Li, S.; Arjunan, A.; Jeong, J.-U.; Jung, Y.D. (−)-Epigallocatechin-3-Gallate Prevents IL-1β-Induced UPAR Expression and Invasiveness via the Suppression of NF-ΚB and AP-1 in Human Bladder Cancer Cells. Int. J. Mol. Sci. 2022, 23, 14008. [Google Scholar] [CrossRef] [PubMed]
- Ersahin, T.; Tuncbag, N.; Cetin-Atalay, R. The PI3K/AKT/MTOR Interactive Pathway. Mol. Biosyst. 2015, 11, 1946–1954. [Google Scholar] [CrossRef]
- LoRusso, P.M. Inhibition of the PI3K/AKT/MTOR Pathway in Solid Tumors. J. Clin. Oncol. 2016, 34, 3803. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Dai, D.; Chen, B.; Tang, H.; Xie, X.; Wei, W. Efficacy of PI3K/AKT/MTOR Pathway Inhibitors for the Treatment of Advanced Solid Cancers: A Literature-Based Meta-Analysis of 46 Randomised Control Trials. PLoS ONE 2018, 13, e0192464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgensztern, D.; McLeod, H.L. PI3K/Akt/MTOR Pathway as a Target for Cancer Therapy. Anti-Cancer Drugs 2005, 16, 797–803. [Google Scholar] [CrossRef]
- Porta, C.; Paglino, C.; Mosca, A. Targeting PI3K/Akt/MTOR Signaling in Cancer. Front. Oncol. 2014, 4, 64. [Google Scholar] [CrossRef] [Green Version]
- Alzahrani, A.S. PI3K/Akt/MTOR Inhibitors in Cancer: At the Bench and Bedside. Semin. Cancer Biol. 2019, 59, 125–132. [Google Scholar] [CrossRef]
- Shorning, B.Y.; Dass, M.S.; Smalley, M.J.; Pearson, H.B. The PI3K-AKT-MTOR Pathway and Prostate Cancer: At the Crossroads of AR, MAPK, and WNT Signaling. Int. J. Mol. Sci. 2020, 21, 4507. [Google Scholar] [CrossRef]
- Liu, S.; Wang, X.-J.; Liu, Y.; Cui, Y.-F. PI3K/AKT/MTOR Signaling Is Involved in (−)-Epigallocatechin-3-Gallate-Induced Apoptosis of Human Pancreatic Carcinoma Cells. Am. J. Chin. Med. 2013, 41, 629–642. [Google Scholar] [CrossRef]
- Ding, F.; Yang, S. Epigallocatechin-3-Gallate Inhibits Proliferation and Triggers Apoptosis in Colon Cancer via the Hedgehog/Phosphoinositide 3-Kinase Pathways. Can. J. Physiol. Pharmacol. 2021, 99, 910–920. [Google Scholar] [CrossRef]
- Ferrari, E.; Bettuzzi, S.; Naponelli, V. The Potential of Epigallocatechin Gallate (EGCG) in Targeting Autophagy for Cancer Treatment: A Narrative Review. Int. J. Mol. Sci. 2022, 23, 6075. [Google Scholar] [CrossRef]
- Ménard, S.; Castronovo, V.; Tagliabue, E.; Sobel, M.E. New Insights into the Metastasis-Associated 67 KD Laminin Receptor. J. Cell Biochem. 1997, 67, 155–165. [Google Scholar] [CrossRef]
- Tanaka, M.; Narumi, K.; Isemura, M.; Abe, M.; Sato, Y.; Abe, T.; Saijo, Y.; Nukiwa, T.; Satoh, K. Expression of the 37-KDa Laminin Binding Protein in Murine Lung Tumor Cell Correlates with Tumor Angiogenesis. Cancer Lett. 2000, 153, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, H.; Koga, K.; Fujimura, Y.; Yamada, K. A Receptor for Green Tea Polyphenol EGCG. Nat. Struct. Mol. Biol. 2004, 11, 380–381. [Google Scholar] [CrossRef]
- Umeda, D.; Yano, S.; Yamada, K.; Tachibana, H. Green Tea Polyphenol Epigallocatechin-3-Gallate Signaling Pathway through 67-KDa Laminin Receptor. J. Biol. Chem. 2008, 283, 3050–3058. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Zhao, J.; Wang, Q.; Chen, P.; Hong, Y.; He, X.; Chen, D.; Liu, H.; Wang, Y.; Cai, X. The Invasive Potential of Hepatoma Cells Induced by Radiotherapy Is Related to the Activation of Hepatic Stellate Cells and Could Be Inhibited by EGCG through the TLR4 Signaling Pathway. Radiat. Res. 2022, 197, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Badana, A.; Chintala, M.; Varikuti, G.; Pudi, N.; Kumari, S.; Kappala, V.R.; Malla, R.R. Lipid Raft Integrity Is Required for Survival of Triple Negative Breast Cancer Cells. J. Breast Cancer 2016, 19, 372–384. [Google Scholar] [CrossRef]
- Greenlee, J.D.; Subramanian, T.; Liu, K.; King, M.R. Rafting down the Metastatic Cascade: The Role of Lipid Rafts in Cancer Metastasis, Cell Death, and Clinical Outcomes. Cancer Res. 2021, 81, 5–17. [Google Scholar] [CrossRef]
- Mollinedo, F.; Gajate, C. Lipid Rafts as Signaling Hubs in Cancer Cell Survival/Death and Invasion: Implications in Tumor Progression and Therapy. J. Lipid Res. 2020, 61, 611–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vona, R.; Iessi, E.; Matarrese, P. Role of Cholesterol and Lipid Rafts in Cancer Signaling: A Promising Therapeutic Opportunity? Front. Cell Dev. Biol. 2021, 9, 622908. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Huang, Y.; Kumazoe, M.; Lesnick, C.; Yamada, S.; Ueda, N.; Suzuki, T.; Yamashita, S.; Kim, Y.H.; Fujimura, Y.; et al. Sphingosine Kinase-1 Protects Multiple Myeloma from Apoptosis Driven by Cancer-Specific Inhibition of RTKs. Mol. Cancer Ther. 2015, 14, 2303–2312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukamoto, S.; Huang, Y.; Umeda, D.; Yamada, S.; Yamashita, S.; Kumazoe, M.; Kim, Y.; Murata, M.; Yamada, K.; Tachibana, H. 67-KDa Laminin Receptor-Dependent Protein Phosphatase 2A (PP2A) Activation Elicits Melanoma-Specific Antitumor Activity Overcoming Drug Resistance. J. Biol. Chem. 2014, 289, 32671–32681. [Google Scholar] [CrossRef] [Green Version]
- Hardie, D.G. AMPK: A Target for Drugs and Natural Products with Effects on Both Diabetes and Cancer. Diabetes 2013, 62, 2164–2172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salminen, A.; Kaarniranta, K. AMP-Activated Protein Kinase (AMPK) Controls the Aging Process via an Integrated Signaling Network. Ageing Res. Rev. 2012, 11, 230–241. [Google Scholar] [CrossRef]
- Winder, W.W.; Hardie, D.G. AMP-Activated Protein Kinase, a Metabolic Master Switch: Possible Roles in Type 2 Diabetes. Am. J. Physiol. 1999, 277, E1–E10. [Google Scholar] [CrossRef]
- Yang, C.S.; Zhang, J.; Zhang, L.; Huang, J.; Wang, Y. Mechanisms of Body Weight Reduction and Metabolic Syndrome Alleviation by Tea. Mol. Nutr. Food Res. 2016, 60, 160–174. [Google Scholar] [CrossRef] [Green Version]
- Mokra, D.; Joskova, M.; Mokry, J. Therapeutic Effects of Green Tea Polyphenol (−)-Epigallocatechin-3-Gallate (EGCG) in Relation to Molecular Pathways Controlling Inflammation, Oxidative Stress, and Apoptosis. Int. J. Mol. Sci. 2023, 24, 340. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Saud, S.M.; Young, M.R.; Chen, G.; Hua, B. Targeting AMPK for Cancer Prevention and Treatment. Oncotarget 2015, 6, 7365–7378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultze, A.; Fiedler, W. Therapeutic Potential and Limitations of New FAK Inhibitors in the Treatment of Cancer. Expert. Opin. Investig. Drugs 2010, 19, 777–788. [Google Scholar] [CrossRef]
- Lv, P.-C.; Jiang, A.-Q.; Zhang, W.-M.; Zhu, H.-L. FAK Inhibitors in Cancer, a Patent Review. Expert. Opin. Ther. Pat. 2018, 28, 139–145. [Google Scholar] [CrossRef]
- Schaller, M.D. Cellular Functions of FAK Kinases: Insight into Molecular Mechanisms and Novel Functions. J. Cell Sci. 2010, 123, 1007–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumazoe, M.; Kadomatsu, M.; Bae, J.; Otsuka, Y.; Fujimura, Y.; Tachibana, H. Src Mediates Epigallocatechin-3-O-Gallate-Elicited Acid Sphingomyelinase Activation. Molecules 2020, 25, 5481. [Google Scholar] [CrossRef] [PubMed]
- Sen, T.; Chatterjee, A. Epigallocatechin-3-Gallate (EGCG) Downregulates EGF-Induced MMP-9 in Breast Cancer Cells: Involvement of Integrin Receptor A5β1 in the Process. Eur. J. Nutr. 2011, 50, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.D.; Chen, S.H.; Lin, C.L.; Tsai, S.H.; Liang, Y.C. Inhibition of Melanoma Growth and Metastasis by Combination with (−)-Epigallocatechin-3-Gallate and Dacarbazine in Mice. J. Cell Biochem. 2001, 83, 631–642. [Google Scholar] [CrossRef]
- Jiang, J.; Hui, C.-C. Hedgehog Signaling in Development and Cancer. Dev. Cell 2008, 15, 801–812. [Google Scholar] [CrossRef] [Green Version]
- Pasca di Magliano, M.; Hebrok, M. Hedgehog Signalling in Cancer Formation and Maintenance. Nat. Rev. Cancer 2003, 3, 903–911. [Google Scholar] [CrossRef]
- Tang, S.-N.; Fu, J.; Nall, D.; Rodova, M.; Shankar, S.; Srivastava, R.K. Inhibition of Sonic Hedgehog Pathway and Pluripotency Maintaining Factors Regulate Human Pancreatic Cancer Stem Cell Characteristics. Int. J. Cancer 2012, 131, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Andersson, E.R.; Lendahl, U. Therapeutic Modulation of Notch Signalling—Are We There Yet? Nat. Rev. Drug Discov. 2014, 13, 357–378. [Google Scholar] [CrossRef]
- Bray, S.J. Notch Signalling: A Simple Pathway Becomes Complex. Nat. Rev. Mol. Cell Biol. 2006, 7, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Purow, B. NOTCH Inhibition as a Promising New Approach to Cancer Therapy. Adv. Exp. Med. Biol. 2012, 727, 305–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farooqi, A.A.; Pinheiro, M.; Granja, A.; Farabegoli, F.; Reis, S.; Attar, R.; Sabitaliyevich, U.Y.; Xu, B.; Ahmad, A. EGCG Mediated Targeting of Deregulated Signaling Pathways and Non-Coding RNAs in Different Cancers: Focus on JAK/STAT, Wnt/β-Catenin, TGF/SMAD, NOTCH, SHH/GLI, and TRAIL Mediated Signaling Pathways. Cancers 2020, 12, 951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, H.; Ge, Q.; Zhang, L.-Y.; Xie, J.; Gan, R.-H.; Lu, Y.-G.; Zheng, D.-L. EGCG Inhibits Growth of Tumoral Lesions on Lip and Tongue of K-Ras Transgenic Mice through the Notch Pathway. J. Nutr. Biochem. 2022, 99, 108843. [Google Scholar] [CrossRef]
- Anastas, J.N.; Moon, R.T. WNT Signalling Pathways as Therapeutic Targets in Cancer. Nat. Rev. Cancer 2013, 13, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Zhan, T.; Rindtorff, N.; Boutros, M. Wnt Signaling in Cancer. Oncogene 2017, 36, 1461–1473. [Google Scholar] [CrossRef]
- Zhu, J.; Jiang, Y.; Yang, X.; Wang, S.; Xie, C.; Li, X.; Li, Y.; Chen, Y.; Wang, X.; Meng, Y.; et al. Wnt/β-Catenin Pathway Mediates (−)-Epigallocatechin-3-Gallate (EGCG) Inhibition of Lung Cancer Stem Cells. Biochem. Biophys. Res. Commun. 2017, 482, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.H.S.; Ferreira, R.S.; Caffarena, E.R. Integrating Molecular Docking and Molecular Dynamics Simulations. Methods Mol. Biol. 2019, 2053, 13–34. [Google Scholar] [CrossRef]
- Talele, T.T.; Khedkar, S.A.; Rigby, A.C. Successful Applications of Computer Aided Drug Discovery: Moving Drugs from Concept to the Clinic. Curr. Top. Med. Chem. 2010, 10, 127–141. [Google Scholar] [CrossRef]
- Bhattacharjee, R.; Devi, A.; Mishra, S. Molecular Docking and Molecular Dynamics Studies Reveal Structural Basis of Inhibition and Selectivity of Inhibitors EGCG and OSU-03012 toward Glucose Regulated Protein-78 (GRP78) Overexpressed in Glioblastoma. J. Mol. Model. 2015, 21, 272. [Google Scholar] [CrossRef]
- Wang, J.; Yin, Y.; Hua, H.; Li, M.; Luo, T.; Xu, L.; Wang, R.; Liu, D.; Zhang, Y.; Jiang, Y. Blockade of GRP78 Sensitizes Breast Cancer Cells to Microtubules-Interfering Agents That Induce the Unfolded Protein Response. J. Cell Mol. Med. 2009, 13, 3888–3897. [Google Scholar] [CrossRef] [Green Version]
- Luo, T.; Wang, J.; Yin, Y.; Hua, H.; Jing, J.; Sun, X.; Li, M.; Zhang, Y.; Jiang, Y. (−)-Epigallocatechin Gallate Sensitizes Breast Cancer Cells to Paclitaxel in a Murine Model of Breast Carcinoma. Breast Cancer Res. 2010, 12, R8. [Google Scholar] [CrossRef]
- Khan, A.; Mohammad, T.; Shamsi, A.; Hussain, A.; Alajmi, M.F.; Husain, S.A.; Iqbal, M.A.; Hassan, M.I. Identification of Plant-Based Hexokinase 2 Inhibitors: Combined Molecular Docking and Dynamics Simulation Studies. J. Biomol. Struct. Dyn. 2022, 40, 10319–10331. [Google Scholar] [CrossRef] [PubMed]
- Ciscato, F.; Ferrone, L.; Masgras, I.; Laquatra, C.; Rasola, A. Hexokinase 2 in Cancer: A Prima Donna Playing Multiple Characters. Int. J. Mol. Sci. 2021, 22, 4716. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Li, M.; Liu, W.-B.; Zhou, Z.-S.; Zhang, R.; Li, J.-L.; Zhou, K.-C. Epigallocatechin Gallate Inhibits Human Tongue Carcinoma Cells via HK2-mediated Glycolysis. Oncol. Rep. 2015, 33, 1533–1539. [Google Scholar] [CrossRef] [Green Version]
- Wei, R.; Hackman, R.M.; Wang, Y.; Mackenzie, G.G. Targeting Glycolysis with Epigallocatechin-3-Gallate Enhances the Efficacy of Chemotherapeutics in Pancreatic Cancer Cells and Xenografts. Cancers 2019, 11, 1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gobessi, S.; Laurenti, L.; Longo, P.G.; Sica, S.; Leone, G.; Efremov, D.G. ZAP-70 Enhances B-Cell-Receptor Signaling despite Absent or Inefficient Tyrosine Kinase Activation in Chronic Lymphocytic Leukemia and Lymphoma B Cells. Blood 2007, 109, 2032–2039. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Moore, A.; Ringshausen, I. ZAP-70 Shapes the Immune Microenvironment in B Cell Malignancies. Front. Oncol. 2020, 10, 595832. [Google Scholar] [CrossRef]
- Shim, J.-H.; Choi, H.S.; Pugliese, A.; Lee, S.-Y.; Chae, J.-I.; Choi, B.Y.; Bode, A.M.; Dong, Z. (−)-Epigallocatechin Gallate Regulates CD3-Mediated T Cell Receptor Signaling in Leukemia through the Inhibition of ZAP-70 Kinase. J. Biol. Chem. 2008, 283, 28370–28379. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Xiong, X.; Li, X.; Zhang, Q.; Yang, W.; Du, L. In Silico Investigation of the Anti-Tumor Mechanisms of Epigallocatechin-3-Gallate. Molecules 2019, 24, 1445. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Tang, F.; Ermakova, S.; Li, M.; Zhao, Q.; Cho, Y.-Y.; Ma, W.-Y.; Choi, H.-S.; Bode, A.M.; Yang, C.S.; et al. Fyn Is a Novel Target of (−)-Epigallocatechin Gallate in the Inhibition of JB6 Cl41 Cell Transformation. Mol. Carcinog. 2008, 47, 172–183. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.; Rahman, A.; Hasnain, A.; Lalonde, M.; Goldberg, V.M.; Haqqi, T.M. Green Tea Polyphenol Epigallocatechin-3-Gallate Inhibits the IL-1 Beta-Induced Activity and Expression of Cyclooxygenase-2 and Nitric Oxide Synthase-2 in Human Chondrocytes. Free Radic. Biol. Med. 2002, 33, 1097–1105. [Google Scholar] [CrossRef]
- Hung, P.-F.; Wu, B.-T.; Chen, H.-C.; Chen, Y.-H.; Chen, C.-L.; Wu, M.-H.; Liu, H.-C.; Lee, M.-J.; Kao, Y.-H. Antimitogenic Effect of Green Tea (−)-Epigallocatechin Gallate on 3T3-L1 Preadipocytes Depends on the ERK and Cdk2 Pathways. Am. J. Physiol. Cell Physiol. 2005, 288, C1094–C1108. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.-T.; Hung, P.-F.; Chen, H.-C.; Huang, R.-N.; Chang, H.-H.; Kao, Y.-H. The Apoptotic Effect of Green Tea (−)-Epigallocatechin Gallate on 3T3-L1 Preadipocytes Depends on the Cdk2 Pathway. J. Agric. Food Chem. 2005, 53, 5695–5701. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Adams, E.; Van Schepdael, A. Study of Abl1 Tyrosine Kinase Inhibitors by Liquid Chromatography-Electrospray Ionization-Mass Spectrometry. Talanta 2013, 107, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Deana, R.; Turetta, L.; Donella-Deana, A.; Donà, M.; Brunati, A.M.; De Michiel, L.; Garbisa, S. Green Tea Epigallocatechin-3-Gallate Inhibits Platelet Signalling Pathways Triggered by Both Proteolytic and Non-Proteolytic Agonists. Thromb. Haemost. 2003, 89, 866–874. [Google Scholar] [CrossRef] [Green Version]
- Park, S.Y.; Lee, Y.-K.; Kim, Y.-M.; Park, O.J.; Shin, J.-I. Control of AMP-Activated Protein Kinase, Akt, and MTOR in EGCG-Treated HT-29 Colon Cancer Cells. Food Sci. Biotechnol. 2013, 22, 147–151. [Google Scholar] [CrossRef]
- Park, S.B.; Bae, J.W.; Kim, J.M.; Lee, S.G.; Han, M. Antiproliferative and Apoptotic Effect of Epigallocatechin-3-Gallate on Ishikawa Cells Is Accompanied by Sex Steroid Receptor Downregulation. Int. J. Mol. Med. 2012, 30, 1211–1218. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.K.; Umar, S.; Riegsecker, S.; Chourasia, M.; Ahmed, S. Regulation of Transforming Growth Factor β-Activated Kinase Activation by Epigallocatechin-3-Gallate in Rheumatoid Arthritis Synovial Fibroblasts: Suppression of K(63) -Linked Autoubiquitination of Tumor Necrosis Factor Receptor-Associated Factor 6. Arthritis Rheumatol. 2016, 68, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.-P.; Kuo, S.-C.; Huang, W.-W.; Yang, J.-S.; Lai, K.-C.; Chen, H.-J.; Lin, K.-L.; Chiu, Y.-J.; Huang, L.-J.; Chung, J.-G. (−)-Epigallocatechin Gallate Induced Apoptosis in Human Adrenal Cancer NCI-H295 Cells through Caspase-Dependent and Caspase-Independent Pathway. Anticancer Res. 2009, 29, 1435–1442. [Google Scholar]
- Matheson, C.J.; Backos, D.S.; Reigan, P. Targeting WEE1 Kinase in Cancer. Trends Pharmacol. Sci. 2016, 37, 872–881. [Google Scholar] [CrossRef]
- Zhu, G.; Pei, L.; Xia, H.; Tang, Q.; Bi, F. Role of Oncogenic KRAS in the Prognosis, Diagnosis and Treatment of Colorectal Cancer. Mol. Cancer 2021, 20, 143. [Google Scholar] [CrossRef]
- Huang, L.; Guo, Z.; Wang, F.; Fu, L. KRAS Mutation: From Undruggable to Druggable in Cancer. Signal Transduct. Target. Ther. 2021, 6, 386. [Google Scholar] [CrossRef] [PubMed]
- Pierotti, M.A.; Greco, A. Oncogenic Rearrangements of the NTRK1/NGF Receptor. Cancer Lett. 2006, 232, 90–98. [Google Scholar] [CrossRef]
- Cui, F.; Yang, K.; Li, Y. Investigate the Binding of Catechins to Trypsin Using Docking and Molecular Dynamics Simulation. PLoS ONE 2015, 10, e0125848. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.M.; Daniel, K.G.; Wang, Z.; Guida, W.C.; Chan, T.-H.; Dou, Q.P. Docking Studies and Model Development of Tea Polyphenol Proteasome Inhibitors: Applications to Rational Drug Design. Proteins 2004, 54, 58–70. [Google Scholar] [CrossRef] [PubMed]
- Kuban-Jankowska, A.; Kostrzewa, T.; Musial, C.; Barone, G.; Lo-Bosco, G.; Lo-Celso, F.; Gorska-Ponikowska, M. Green Tea Catechins Induce Inhibition of PTP1B Phosphatase in Breast Cancer Cells with Potent Anti-Cancer Properties: In Vitro Assay, Molecular Docking, and Dynamics Studies. Antioxidants 2020, 9, 1208. [Google Scholar] [CrossRef]
- Artali, R.; Beretta, G.; Morazzoni, P.; Bombardelli, E.; Meneghetti, F. Green Tea Catechins in Chemoprevention of Cancer: A Molecular Docking Investigation into Their Interaction with Glutathione S-Transferase (GST P1-1). J. Enzyme Inhib. Med. Chem. 2009, 24, 287–295. [Google Scholar] [CrossRef]
- Fang, M.Z.; Wang, Y.; Ai, N.; Hou, Z.; Sun, Y.; Lu, H.; Welsh, W.; Yang, C.S. Tea Polyphenol (−)-Epigallocatechin-3-Gallate Inhibits DNA Methyltransferase and Reactivates Methylation-Silenced Genes in Cancer Cell Lines. Cancer Res. 2003, 63, 7563–7570. [Google Scholar]
- Rahnasto-Rilla, M.; Tyni, J.; Huovinen, M.; Jarho, E.; Kulikowicz, T.; Ravichandran, S.; Bohr, V.A.; Ferrucci, L.; Lahtela-Kakkonen, M.; Moaddel, R. Natural Polyphenols as Sirtuin 6 Modulators. Sci. Rep. 2018, 8, 4163. [Google Scholar] [CrossRef] [Green Version]
- Saeki, K.; Hayakawa, S.; Nakano, S.; Ito, S.; Oishi, Y.; Suzuki, Y.; Isemura, M. In Vitro and In Silico Studies of the Molecular Interactions of Epigallocatechin-3-O-Gallate (EGCG) with Proteins That Explain the Health Benefits of Green Tea. Molecules 2018, 23, 1295. [Google Scholar] [CrossRef] [Green Version]
- Nakano, S.; Megro, S.; Hase, T.; Suzuki, T.; Isemura, M.; Nakamura, Y.; Ito, S. Computational Molecular Docking and X-Ray Crystallographic Studies of Catechins in New Drug Design Strategies. Molecules 2018, 23, 2020. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.-M.; Sun, C.; Butler, L.M. Tea and Cancer Prevention: Epidemiological Studies. Pharmacol. Res. 2011, 64, 123–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.-F.; Xu, Q.; Lu, J.; Wang, P.; Zhang, H.-W.; Zhou, L.; Ma, X.-Q.; Zhou, Y.-H. Tea Consumption and the Incidence of Cancer: A Systematic Review and Meta-Analysis of Prospective Observational Studies. Eur. J. Cancer Prev. 2015, 24, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Lee, J.E.; Loftfield, E.; Shu, X.-O.; Abe, S.K.; Rahman, M.S.; Saito, E.; Islam, M.R.; Tsugane, S.; Sawada, N.; et al. Coffee and Tea Consumption and Mortality from All Causes, Cardiovascular Disease and Cancer: A Pooled Analysis of Prospective Studies from the Asia Cohort Consortium. Int. J. Epidemiol. 2022, 51, 626–640. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, J.; Oh, J.H.; Chang, H.J.; Sohn, D.K.; Shin, A.; Kim, J. Protective Effect of Green Tea Consumption on Colorectal Cancer Varies by Lifestyle Factors. Nutrients 2019, 11, 2612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, P.; Zheng, H.-M.; Deng, X.-M.; Zhang, Y. Green Tea Consumption and Risk of Esophageal Cancer: A Meta-Analysis of Epidemiologic Studies. BMC Gastroenterol. 2012, 12, 165. [Google Scholar] [CrossRef] [Green Version]
- Sang, L.-X.; Chang, B.; Li, X.-H.; Jiang, M. Green Tea Consumption and Risk of Esophageal Cancer: A Meta-Analysis of Published Epidemiological Studies. Nutr. Cancer 2013, 65, 802–812. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, N.; Zhuang, W.; Liu, G.; Wu, T.; Yao, X.; Du, L.; Wei, M.; Wu, X. Green Tea and Gastric Cancer Risk: Meta-Analysis of Epidemiologic Studies. Asia Pac. J. Clin. Nutr. 2008, 17, 159–165. [Google Scholar]
- Meshitsuka, S.; Shingaki, S.; Hotta, M.; Goto, M.; Kobayashi, M.; Ukawa, Y.; Sagesaka, Y.M.; Wada, Y.; Nojima, M.; Suzuki, K. Phase 2 Trial of Daily, Oral Epigallocatechin Gallate in Patients with Light-Chain Amyloidosis. Int. J. Hematol. 2017, 105, 295–308. [Google Scholar] [CrossRef]
- Widmer, R.J.; Freund, M.A.; Flammer, A.J.; Sexton, J.; Lennon, R.; Romani, A.; Mulinacci, N.; Vinceri, F.F.; Lerman, L.O.; Lerman, A. Beneficial Effects of Polyphenol-Rich Olive Oil in Patients with Early Atherosclerosis. Eur. J. Nutr. 2013, 52, 1223–1231. [Google Scholar] [CrossRef] [Green Version]
- De Stefano, D.; Villella, V.R.; Esposito, S.; Tosco, A.; Sepe, A.; De Gregorio, F.; Salvadori, L.; Grassia, R.; Leone, C.A.; De Rosa, G.; et al. Restoration of CFTR Function in Patients with Cystic Fibrosis Carrying the F508del-CFTR Mutation. Autophagy 2014, 10, 2053–2074. [Google Scholar] [CrossRef] [Green Version]
- Tosco, A.; De Gregorio, F.; Esposito, S.; De Stefano, D.; Sana, I.; Ferrari, E.; Sepe, A.; Salvadori, L.; Buonpensiero, P.; Di Pasqua, A.; et al. A Novel Treatment of Cystic Fibrosis Acting On-Target: Cysteamine plus Epigallocatechin Gallate for the Autophagy-Dependent Rescue of Class II-Mutated CFTR. Cell Death Differ. 2016, 23, 1380–1393. [Google Scholar] [CrossRef] [Green Version]
- Joe, A.K.; Schnoll-Sussman, F.; Bresalier, R.S.; Abrams, J.A.; Hibshoosh, H.; Cheung, K.; Friedman, R.A.; Yang, C.S.; Milne, G.L.; Liu, D.D.; et al. Phase Ib Randomized, Double-Blinded, Placebo-Controlled, Dose Escalation Study of Polyphenon E in Patients with Barrett’s Esophagus. Cancer Prev. Res. 2015, 8, 1131–1137. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.-M.; Chang, Y.-H.; Chao, Y.-C.; Lin, J.-A.; Wu, C.-H.; Lai, C.-Y.; Chan, K.-C.; Tseng, S.-T.; Yen, G.-C. EGCG-Rich Green Tea Extract Stimulates SRAGE Secretion to Inhibit S100A12-RAGE Axis through ADAM10-Mediated Ectodomain Shedding of Extracellular RAGE in Type 2 Diabetes. Mol. Nutr. Food Res. 2013, 57, 2264–2268. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Su, S.; Yu, X.; Li, Y. Dietary Epigallocatechin 3-Gallate Supplement Improves Maternal and Neonatal Treatment Outcome of Gestational Diabetes Mellitus: A Double-Blind Randomised Controlled Trial. J. Hum. Nutr. Diet. 2017, 30, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Bazyar, H.; Hosseini, S.A.; Saradar, S.; Mombaini, D.; Allivand, M.; Labibzadeh, M.; Alipour, M. Effects of Epigallocatechin-3-Gallate of Camellia sinensis Leaves on Blood Pressure, Lipid Profile, Atherogenic Index of Plasma and Some Inflammatory and Antioxidant Markers in Type 2 Diabetes Mellitus Patients: A Clinical Trial. J. Complement. Integr. Med. 2020, 18, 405–411. [Google Scholar] [CrossRef] [PubMed]
- De la Torre, R.; De Sola, S.; Pons, M.; Duchon, A.; de Lagran, M.M.; Farré, M.; Fitó, M.; Benejam, B.; Langohr, K.; Rodriguez, J.; et al. Epigallocatechin-3-Gallate, a DYRK1A Inhibitor, Rescues Cognitive Deficits in Down Syndrome Mouse Models and in Humans. Mol. Nutr. Food Res. 2014, 58, 278–288. [Google Scholar] [CrossRef]
- de la Torre, R.; de Sola, S.; Hernandez, G.; Farré, M.; Pujol, J.; Rodriguez, J.; Espadaler, J.M.; Langohr, K.; Cuenca-Royo, A.; Principe, A.; et al. Safety and Efficacy of Cognitive Training plus Epigallocatechin-3-Gallate in Young Adults with Down’s Syndrome (TESDAD): A Double-Blind, Randomised, Placebo-Controlled, Phase 2 Trial. Lancet Neurol. 2016, 15, 801–810. [Google Scholar] [CrossRef]
- Xicota, L.; Rodríguez, J.; Langohr, K.; Fitó, M.; Dierssen, M.; de la Torre, R.; TESDAD Study Group. Effect of Epigallocatechin Gallate on the Body Composition and Lipid Profile of down Syndrome Individuals: Implications for Clinical Management. Clin. Nutr. 2020, 39, 1292–1300. [Google Scholar] [CrossRef]
- Cieuta-Walti, C.; Cuenca-Royo, A.; Langohr, K.; Rakic, C.; López-Vílchez, M.Á.; Lirio, J.; González-Lamuño Leguina, D.; González, T.B.; García, J.G.; Roure, M.R.; et al. Safety and Preliminary Efficacy on Cognitive Performance and Adaptive Functionality of Epigallocatechin Gallate (EGCG) in Children with Down Syndrome. A Randomized Phase Ib Clinical Trial (PERSEUS Study). Genet. Med. 2022, 24, 2004–2013. [Google Scholar] [CrossRef]
- Chiaverini, C.; Roger, C.; Fontas, E.; Bourrat, E.; Bourdon-Lanoy, E.; Labrèze, C.; Mazereeuw, J.; Vabres, P.; Bodemer, C.; Lacour, J.-P. Oral Epigallocatechin-3-Gallate for Treatment of Dystrophic Epidermolysis Bullosa: A Multicentre, Randomized, Crossover, Double-Blind, Placebo-Controlled Clinical Trial. Orphanet J. Rare Dis. 2016, 11, 31. [Google Scholar] [CrossRef] [Green Version]
- Arazi, H.; Samami, N.; Kheirkhah, J.; Taati, B. The Effect of Three Weeks Green Tea Extract Consumption on Blood Pressure, Heart Rate Responses to a Single Bout Resistance Exercise in Hypertensive Women. High Blood Press. Cardiovasc. Prev. 2014, 21, 213–219. [Google Scholar] [CrossRef]
- Maeda-Yamamoto, M.; Nishimura, M.; Kitaichi, N.; Nesumi, A.; Monobe, M.; Nomura, S.; Horie, Y.; Tachibana, H.; Nishihira, J. A Randomized, Placebo-Controlled Study on the Safety and Efficacy of Daily Ingestion of Green Tea (Camellia sinensis L.) Cv. “Yabukita” and “Sunrouge” on Eyestrain and Blood Pressure in Healthy Adults. Nutrients 2018, 10, 569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mähler, A.; Steiniger, J.; Bock, M.; Klug, L.; Parreidt, N.; Lorenz, M.; Zimmermann, B.F.; Krannich, A.; Paul, F.; Boschmann, M. Metabolic Response to Epigallocatechin-3-Gallate in Relapsing-Remitting Multiple Sclerosis: A Randomized Clinical Trial. Am. J. Clin. Nutr. 2015, 101, 487–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovera, J.; Ramos, A.; Devier, D.; Garrison, V.; Kovner, B.; Reza, T.; Koop, D.; Rooney, W.; Foundas, A.; Bourdette, D. Polyphenon E, Non-Futile at Neuroprotection in Multiple Sclerosis but Unpredictably Hepatotoxic: Phase I Single Group and Phase II Randomized Placebo-Controlled Studies. J. Neurol. Sci. 2015, 358, 46–52. [Google Scholar] [CrossRef] [Green Version]
- Platero, J.L.; Cuerda-Ballester, M.; Ibáñez, V.; Sancho, D.; Lopez-Rodríguez, M.M.; Drehmer, E.; de la Rubia Ortí, J.E. The Impact of Coconut Oil and Epigallocatechin Gallate on the Levels of IL-6, Anxiety and Disability in Multiple Sclerosis Patients. Nutrients 2020, 12, 305. [Google Scholar] [CrossRef] [Green Version]
- Benlloch, M.; Cuerda Ballester, M.; Drehmer, E.; Platero, J.L.; Carrera-Juliá, S.; López-Rodríguez, M.M.; Ceron, J.J.; Tvarijonaviciute, A.; Navarro, M.Á.; Moreno, M.L.; et al. Possible Reduction of Cardiac Risk after Supplementation with Epigallocatechin Gallate and Increase of Ketone Bodies in the Blood in Patients with Multiple Sclerosis. A Pilot Study. Nutrients 2020, 12, 3792. [Google Scholar] [CrossRef] [PubMed]
- Rust, R.; Chien, C.; Scheel, M.; Brandt, A.U.; Dörr, J.; Wuerfel, J.; Klumbies, K.; Zimmermann, H.; Lorenz, M.; Wernecke, K.-D.; et al. Epigallocatechin Gallate in Progressive MS: A Randomized, Placebo-Controlled Trial. Neurol. Neuroimmunol. Neuroinflamm. 2021, 8, e964. [Google Scholar] [CrossRef] [PubMed]
- Bellmann-Strobl, J.; Paul, F.; Wuerfel, J.; Dörr, J.; Infante-Duarte, C.; Heidrich, E.; Körtgen, B.; Brandt, A.; Pfüller, C.; Radbruch, H.; et al. Epigallocatechin Gallate in Relapsing-Remitting Multiple Sclerosis: A Randomized, Placebo-Controlled Trial. Neurol. Neuroimmunol. Neuroinflamm. 2021, 8, e981. [Google Scholar] [CrossRef]
- de la Rubia Ortí, J.E.; Platero, J.L.; Yang, I.H.; Ceron, J.J.; Tvarijonaviciute, A.; Sabater, P.S.; Benlloch, M.; Sancho-Cantus, D.; Sancho, S. Possible Role of Butyrylcholinesterase in Fat Loss and Decreases in Inflammatory Levels in Patients with Multiple Sclerosis after Treatment with Epigallocatechin Gallate and Coconut Oil: A Pilot Study. Nutrients 2021, 13, 3230. [Google Scholar] [CrossRef]
- Cuerda-Ballester, M.; Proaño, B.; Alarcón-Jimenez, J.; de Bernardo, N.; Villaron-Casales, C.; Lajara Romance, J.M.; de la Rubia Ortí, J.E. Improvements in Gait and Balance in Patients with Multiple Sclerosis after Treatment with Coconut Oil and Epigallocatechin Gallate. A Pilot Study. Food Funct. 2023, 14, 1062–1071. [Google Scholar] [CrossRef]
- Mielgo-Ayuso, J.; Barrenechea, L.; Alcorta, P.; Larrarte, E.; Margareto, J.; Labayen, I. Effects of Dietary Supplementation with Epigallocatechin-3-Gallate on Weight Loss, Energy Homeostasis, Cardiometabolic Risk Factors and Liver Function in Obese Women: Randomised, Double-Blind, Placebo-Controlled Clinical Trial. Br. J. Nutr. 2014, 111, 1263–1271. [Google Scholar] [CrossRef] [Green Version]
- Chen, I.-J.; Liu, C.-Y.; Chiu, J.-P.; Hsu, C.-H. Therapeutic Effect of High-Dose Green Tea Extract on Weight Reduction: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Clin. Nutr. 2016, 35, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Most, J.; Timmers, S.; Warnke, I.; Jocken, J.W.; van Boekschoten, M.; de Groot, P.; Bendik, I.; Schrauwen, P.; Goossens, G.H.; Blaak, E.E. Combined Epigallocatechin-3-Gallate and Resveratrol Supplementation for 12 Wk Increases Mitochondrial Capacity and Fat Oxidation, but Not Insulin Sensitivity, in Obese Humans: A Randomized Controlled Trial. Am. J. Clin. Nutr. 2016, 104, 215–227. [Google Scholar] [CrossRef] [Green Version]
- Most, J.; Warnke, I.; Boekschoten, M.V.; Jocken, J.W.E.; de Groot, P.; Friedel, A.; Bendik, I.; Goossens, G.H.; Blaak, E.E. The Effects of Polyphenol Supplementation on Adipose Tissue Morphology and Gene Expression in Overweight and Obese Humans. Adipocyte 2018, 7, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Cazzola, R.; Rondanelli, M. N-Oleoyl-Phosphatidyl-Ethanolamine and Epigallo Catechin-3-Gallate Mitigate Oxidative Stress in Overweight and Class I Obese People on a Low-Calorie Diet. J. Med. Food 2020, 23, 319–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatree, S.; Sitticharoon, C.; Maikaew, P.; Pongwattanapakin, K.; Keadkraichaiwat, I.; Churintaraphan, M.; Sripong, C.; Sririwichitchai, R.; Tapechum, S. Epigallocatechin Gallate Decreases Plasma Triglyceride, Blood Pressure, and Serum Kisspeptin in Obese Human Subjects. Exp. Biol. Med. 2021, 246, 163–176. [Google Scholar] [CrossRef]
- Roberts, J.D.; Willmott, A.G.B.; Beasley, L.; Boal, M.; Davies, R.; Martin, L.; Chichger, H.; Gautam, L.; Del Coso, J. The Impact of Decaffeinated Green Tea Extract on Fat Oxidation, Body Composition and Cardio-Metabolic Health in Overweight, Recreationally Active Individuals. Nutrients 2021, 13, 764. [Google Scholar] [CrossRef]
- Gu, Q.; Wang, X.; Xie, L.; Yao, X.; Qian, L.; Yu, Z.; Shen, X. Green Tea Catechin EGCG Could Prevent Obesity-Related Precocious Puberty through NKB/NK3R Signaling Pathway. J. Nutr. Biochem. 2022, 108, 109085. [Google Scholar] [CrossRef]
- Zhao, H.; Zhu, W.; Xie, P.; Li, H.; Zhang, X.; Sun, X.; Yu, J.; Xing, L. A Phase I Study of Concurrent Chemotherapy and Thoracic Radiotherapy with Oral Epigallocatechin-3-Gallate Protection in Patients with Locally Advanced Stage III Non-Small-Cell Lung Cancer. Radiother. Oncol. 2014, 110, 132–136. [Google Scholar] [CrossRef]
- Zhao, H.; Xie, P.; Li, X.; Zhu, W.; Sun, X.; Sun, X.; Chen, X.; Xing, L.; Yu, J. A Prospective Phase II Trial of EGCG in Treatment of Acute Radiation-Induced Esophagitis for Stage III Lung Cancer. Radiother. Oncol. 2015, 114, 351–356. [Google Scholar] [CrossRef]
- Zhao, H.; Jia, L.; Chen, G.; Li, X.; Meng, X.; Zhao, X.; Xing, L.; Zhu, W. A Prospective, Three-Arm, Randomized Trial of EGCG for Preventing Radiation-Induced Esophagitis in Lung Cancer Patients Receiving Radiotherapy. Radiother. Oncol. 2019, 137, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xing, L.; Zhang, Y.; Xie, P.; Zhu, W.; Meng, X.; Wang, Y.; Kong, L.; Zhao, H.; Yu, J. Phase II Trial of Epigallocatechin-3-Gallate in Acute Radiation-Induced Esophagitis for Esophagus Cancer. J. Med. Food 2020, 23, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Dryden, G.W.; Lam, A.; Beatty, K.; Qazzaz, H.H.; McClain, C.J. A Pilot Study to Evaluate the Safety and Efficacy of an Oral Dose of (−)-Epigallocatechin-3-Gallate-Rich Polyphenon E in Patients with Mild to Moderate Ulcerative Colitis. Inflamm. Bowel Dis. 2013, 19, 1904–1912. [Google Scholar] [CrossRef]
- Kowalska, J.; Marzec, A.; Domian, E.; Galus, S.; Ciurzyńska, A.; Brzezińska, R.; Kowalska, H. Influence of Tea Brewing Parameters on the Antioxidant Potential of Infusions and Extracts Depending on the Degree of Processing of the Leaves of Camellia sinensis. Molecules 2021, 26, 4773. [Google Scholar] [CrossRef]
- Lakenbrink, C.; Lapczynski, S.; Maiwald, B.; Engelhardt, U.H. Flavonoids and Other Polyphenols in Consumer Brews of Tea and Other Caffeinated Beverages. J. Agric. Food Chem. 2000, 48, 2848–2852. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhou, D.; Chen, Y. Preparation and Antioxidant Activity of Green Tea Extract Enriched in Epigallocatechin (EGC) and Epigallocatechin Gallate (EGCG). J. Agric. Food Chem. 2009, 57, 1349–1353. [Google Scholar] [CrossRef]
- Saklar, S.; Ertas, E.; Ozdemir, I.S.; Karadeniz, B. Effects of Different Brewing Conditions on Catechin Content and Sensory Acceptance in Turkish Green Tea Infusions. J. Food Sci. Technol. 2015, 52, 6639–6646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Xu, Y.-F.; Feng, Y.; Yang, F.-Q.; Luo, J.; Zhai, W.; Che, J.-P.; Wang, G.-C.; Zheng, J.-H. Epigallocatechin Gallate Attenuates Interstitial Cystitis in Human Bladder Urothelium Cells by Modulating Purinergic Receptors. J. Surg. Res. 2013, 183, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Trudel, D.; Labbé, D.P.; Araya-Farias, M.; Doyen, A.; Bazinet, L.; Duchesne, T.; Plante, M.; Grégoire, J.; Renaud, M.-C.; Bachvarov, D.; et al. A Two-Stage, Single-Arm, Phase II Study of EGCG-Enriched Green Tea Drink as a Maintenance Therapy in Women with Advanced Stage Ovarian Cancer. Gynecol. Oncol. 2013, 131, 357–361. [Google Scholar] [CrossRef]
- Kumar, N.B.; Pow-Sang, J.; Egan, K.M.; Spiess, P.E.; Dickinson, S.; Salup, R.; Helal, M.; McLarty, J.; Williams, C.R.; Schreiber, F.; et al. Randomized, Placebo-Controlled Trial of Green Tea Catechins for Prostate Cancer Prevention. Cancer Prev. Res. 2015, 8, 879–887. [Google Scholar] [CrossRef] [Green Version]
- Tsampoukas, G.; Manolas, V.; Brown, D.; Dellis, A.; Deliveliotis, K.; Moussa, M.; Papatsoris, A. Atypical Small Acinar Proliferation and Its Significance in Pathological Reports in Modern Urological Times. Asian J. Urol. 2022, 9, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Andras, I.; Telecan, T.; Crisan, D.; Cata, E.; Kadula, P.; Andras, D.; Bungardean, M.; Coman, I.; Crisan, N. Different Clinical Significance of ASAP/HGPIN Pattern in Systematic vs. MRI-US Fusion Guided Prostate Biopsy. Exp. Ther. Med. 2020, 20, 195. [Google Scholar] [CrossRef]
- Bettuzzi, S.; Brausi, M.; Rizzi, F.; Castagnetti, G.; Peracchia, G.; Corti, A. Chemoprevention of Human Prostate Cancer by Oral Administration of Green Tea Catechins in Volunteers with High-Grade Prostate Intraepithelial Neoplasia: A Preliminary Report from a One-Year Proof-of-Principle Study. Cancer Res. 2006, 66, 1234–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brausi, M.; Rizzi, F.; Bettuzzi, S. Chemoprevention of Human Prostate Cancer by Green Tea Catechins: Two Years Later. A Follow-up Update. Eur. Urol. 2008, 54, 472–473. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.B.; Pow-Sang, J.; Spiess, P.E.; Park, J.; Salup, R.; Williams, C.R.; Parnes, H.; Schell, M.J. Randomized, Placebo-Controlled Trial Evaluating the Safety of One-Year Administration of Green Tea Catechins. Oncotarget 2016, 7, 70794–70802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perletti, G.; Magri, V.; Vral, A.; Stamatiou, K.; Trinchieri, A. Green Tea Catechins for Chemoprevention of Prostate Cancer in Patients with Histologically-Proven HG-PIN or ASAP. Concise Review and Meta-Analysis. Arch. Ital. Urol. Androl. 2019, 91. [Google Scholar] [CrossRef] [Green Version]
- Henning, S.M.; Aronson, W.; Niu, Y.; Conde, F.; Lee, N.H.; Seeram, N.P.; Lee, R.-P.; Lu, J.; Harris, D.M.; Moro, A.; et al. Tea Polyphenols and Theaflavins Are Present in Prostate Tissue of Humans and Mice after Green and Black Tea Consumption. J. Nutr. 2006, 136, 1839–1843. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Aronson, W.J.; Huang, M.; Zhang, Y.; Lee, R.-P.; Heber, D.; Henning, S.M. Green Tea Polyphenols and Metabolites in Prostatectomy Tissue: Implications for Cancer Prevention. Cancer Prev. Res. 2010, 3, 985–993. [Google Scholar] [CrossRef] [Green Version]
- McLarty, J.; Bigelow, R.L.H.; Smith, M.; Elmajian, D.; Ankem, M.; Cardelli, J.A. Tea Polyphenols Decrease Serum Levels of Prostate-Specific Antigen, Hepatocyte Growth Factor, and Vascular Endothelial Growth Factor in Prostate Cancer Patients and Inhibit Production of Hepatocyte Growth Factor and Vascular Endothelial Growth Factor in Vitro. Cancer Prev. Res. 2009, 2, 673–682. [Google Scholar] [CrossRef] [Green Version]
- Fhu, C.W.; Ali, A. Fatty Acid Synthase: An Emerging Target in Cancer. Molecules 2020, 25, 3935. [Google Scholar] [CrossRef]
- Sun, X.; Kaufman, P.D. Ki-67: More than a Proliferation Marker. Chromosoma 2018, 127, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Garzotto, M.; Beer, T.M.; Thuillier, P.; Lieberman, S.; Mori, M.; Stoller, W.A.; Farris, P.E.; Shannon, J. Effects of ω-3 Fatty Acids and Catechins on Fatty Acid Synthase in the Prostate: A Randomized Controlled Trial. Nutr. Cancer 2016, 68, 1309–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; McIntosh, G.H.; Le Leu, R.K.; Somashekar, R.; Meng, X.Q.; Gopalsamy, G.; Bambaca, L.; McKinnon, R.A.; Young, G.P. Supplementation with Brazil Nuts and Green Tea Extract Regulates Targeted Biomarkers Related to Colorectal Cancer Risk in Humans. Br. J. Nutr. 2016, 116, 1901–1911. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, H.; Tighiouart, M.; Lee, J.E.; Shin, H.J.; Khuri, F.R.; Yang, C.S.; Chen, Z.; Shin, D.M. Synergistic Inhibition of Head and Neck Tumor Growth by Green Tea (−)-Epigallocatechin-3-Gallate and EGFR Tyrosine Kinase Inhibitor. Int. J. Cancer 2008, 123, 1005–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, D.M.; Nannapaneni, S.; Patel, M.R.; Shi, Q.; Liu, Y.; Chen, Z.; Chen, A.Y.; El-Deiry, M.W.; Beitler, J.J.; Steuer, C.E.; et al. Phase Ib Study of Chemoprevention with Green Tea Polyphenon E and Erlotinib in Patients with Advanced Premalignant Lesions (APL) of the Head and Neck. Clin. Cancer Res. 2020, 26, 5860–5868. [Google Scholar] [CrossRef] [PubMed]
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural Products for Drug Discovery in the 21st Century: Innovations for Novel Drug Discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dube, A.; Nicolazzo, J.A.; Larson, I. Chitosan Nanoparticles Enhance the Plasma Exposure of (−)-Epigallocatechin Gallate in Mice through an Enhancement in Intestinal Stability. Eur. J. Pharm. Sci. Off. J. Eur. Fed. Pharm. Sci. 2011, 44, 422–426. [Google Scholar] [CrossRef]
- Son, Y.-R.; Chung, J.-H.; Ko, S.; Shim, S.-M. Combinational Enhancing Effects of Formulation and Encapsulation on Digestive Stability and Intestinal Transport of Green Tea Catechins. J. Microencapsul. 2016, 33, 183–190. [Google Scholar] [CrossRef]
- Chung, J.-H.; Lee, S.-J.; Chung, J.-O.; Oh, Y.-J.; Hwang, J.-A.; Kim, Y.-K.; Ko, S.; Shim, S.-M. Effect of Hydroxypropyl Methyl Cellulose Phthalate Coating on Digestive Stability and Intestinal Transport of Green Tea Catechins. Integr. Med. Res. 2014, 3, 34–37. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.B.; Tsai, M.J.; Wu, P.C.; Tsai, Y.H.; Wu, Y.H.; Fang, J.Y. Elastic Liposomes as Carriers for Oral Delivery and the Brain Distribution of (+)-Catechin. J. Drug Target. 2011, 19, 709–718. [Google Scholar] [CrossRef]
- Fang, J.Y.; Lee, W.R.; Shen, S.C.; Huang, Y.L. Effect of Liposome Encapsulation of Tea Catechins on Their Accumulation in Basal Cell Carcinomas. J. Dermatol. Sci. 2006, 42, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.Q.; Liu, W.; Liu, W.L.; Liang, R.H.; Li, T.; Liu, C.M.; Cao, Y.L.; Niu, J.; Liu, Z. Characterization and Bioavailability of Tea Polyphenol Nanoliposome Prepared by Combining an Ethanol Injection Method with Dynamic High-Pressure Microfluidization. J. Agric. Food Chem. 2014, 62, 934–941. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Huang, L.-Y.; Nie, N.; Ye, J.-H.; Zheng, X.-Q.; Lu, J.-L.; Liang, Y.-R. Binding of Tea Catechins to Rice Bran Protein Isolate: Interaction and Protective Effect during in Vitro Digestion. Food Res. Int. 2017, 93, 1–7. [Google Scholar] [CrossRef]
- Li, Y.; Lim, L.-T.; Kakuda, Y. Electrospun Zein Fibers as Carriers to Stabilize (−)-Epigallocatechin Gallate. J. Food Sci. 2009, 74, C233–C240. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Ha, J.; Zou, T.; Gu, L. Fabrication of Coated Bovine Serum Albumin (BSA)-Epigallocatechin Gallate (EGCG) Nanoparticles and Their Transport across Monolayers of Human Intestinal Epithelial Caco-2 Cells. Food Funct. 2014, 5, 1278–1285. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Gu, L. Fabrication of Self-Assembled (−)-Epigallocatechin Gallate (EGCG) Ovalbumin-Dextran Conjugate Nanoparticles and Their Transport across Monolayers of Human Intestinal Epithelial Caco-2 Cells. J. Agric. Food Chem. 2014, 62, 1301–1309. [Google Scholar] [CrossRef]
- Sahadevan, R.; Singh, S.; Binoy, A.; Sadhukhan, S. Chemico-Biological Aspects of (−)-Epigallocatechin-3-Gallate (EGCG) to Improve Its Stability, Bioavailability and Membrane Permeability: Current Status and Future Prospects. Crit. Rev. Food Sci. Nutr. 2022, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.-Y.; Li, X.-M.; Liang, J.-P.; Xiang, L.-P.; Wang, K.-R.; Shi, Y.-L.; Yang, R.; Shi, M.; Ye, J.-H.; Lu, J.-L.; et al. Bioavailability of Tea Catechins and Its Improvement. Molecules 2018, 23, 2346. [Google Scholar] [CrossRef] [Green Version]
- Janle, E.M.; Morré, D.M.; Morré, D.J.; Zhou, Q.; Zhu, Y. Pharmacokinetics of Green Tea Catechins in Extract and Sustained-Release Preparations. J. Diet. Suppl. 2008, 5, 248–263. [Google Scholar] [CrossRef] [Green Version]
- Andreu Fernández, V.; Almeida Toledano, L.; Pizarro Lozano, N.; Navarro Tapia, E.; Gómez Roig, M.D.; De la Torre Fornell, R.; García Algar, Ó. Bioavailability of Epigallocatechin Gallate Administered with Different Nutritional Strategies in Healthy Volunteers. Antioxidants 2020, 9, 440. [Google Scholar] [CrossRef]
- Mereles, D.; Hunstein, W. Epigallocatechin-3-Gallate (EGCG) for Clinical Trials: More Pitfalls than Promises? Int. J. Mol. Sci. 2011, 12, 5592–5603. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Lee, M.J.; Li, H.; Yang, C.S. Absorption, Distribution, Elimination of Tea Polyphenols in Rats. Drug Metab. Dispos. 1997, 25, 1045–1050. [Google Scholar]
- Oketch-Rabah, H.A.; Roe, A.L.; Rider, C.V.; Bonkovsky, H.L.; Giancaspro, G.I.; Navarro, V.; Paine, M.F.; Betz, J.M.; Marles, R.J.; Casper, S.; et al. United States Pharmacopeia (USP) Comprehensive Review of the Hepatotoxicity of Green Tea Extracts. Toxicol. Rep. 2020, 7, 386–402. [Google Scholar] [CrossRef]
- Siblini, H.; Al-Hendy, A.; Segars, J.; González, F.; Taylor, H.S.; Singh, B.; Flaminia, A.; Flores, V.A.; Christman, G.M.; Huang, H.; et al. Assessing the Hepatic Safety of Epigallocatechin Gallate (EGCG) in Reproductive-Aged Women. Nutrients 2023, 15, 320. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.D.; Kennett, M.J.; Sang, S.; Reuhl, K.R.; Ju, J.; Yang, C.S. Hepatotoxicity of High Oral Dose (−)-Epigallocatechin-3-Gallate in Mice. Food Chem. Toxicol. 2010, 48, 409–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isbrucker, R.A.; Edwards, J.A.; Wolz, E.; Davidovich, A.; Bausch, J. Safety Studies on Epigallocatechin Gallate (EGCG) Preparations. Part 2: Dermal, Acute and Short-Term Toxicity Studies. Food Chem. Toxicol. 2006, 44, 636–650. [Google Scholar] [CrossRef] [PubMed]
- Kapetanovic, I.M.; Crowell, J.A.; Krishnaraj, R.; Zakharov, A.; Lindeblad, M.; Lyubimov, A. Exposure and Toxicity of Green Tea Polyphenols in Fasted and Non-Fasted Dogs. Toxicology 2009, 260, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Isbrucker, R.A.; Bausch, J.; Edwards, J.A.; Wolz, E. Safety Studies on Epigallocatechin Gallate (EGCG) Preparations. Part 1: Genotoxicity. Food Chem. Toxicol. 2006, 44, 626–635. [Google Scholar] [CrossRef]
- Isbrucker, R.A.; Edwards, J.A.; Wolz, E.; Davidovich, A.; Bausch, J. Safety Studies on Epigallocatechin Gallate (EGCG) Preparations. Part 3: Teratogenicity and Reproductive Toxicity Studies in Rats. Food Chem. Toxicol. 2006, 44, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Webster, D.; Cao, J.; Shao, A. The Safety of Green Tea and Green Tea Extract Consumption in Adults—Results of a Systematic Review. Regul. Toxicol. Pharmacol. 2018, 95, 412–433. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Lambert, J.D.; Lee, S.H.; Sinko, P.J.; Yang, C.S. Involvement of Multidrug Resistance-Associated Proteins in Regulating Cellular Levels of (−)-Epigallocatechin-3-Gallate and Its Methyl Metabolites. Biochem. Biophys. Res. Commun. 2003, 310, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Jodoin, J.; Demeule, M.; Beliveau, R. Inhibition of the Multidrug Resistance P-Glycoprotein Activity by Green Tea Polyphenols. Biochim. Biophys. Acta 2002, 1542, 149–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rein, M.J.; Renouf, M.; Cruz-Hernandez, C.; Actis-Goretta, L.; Thakkar, S.K.; da Silva Pinto, M. Bioavailability of Bioactive Food Compounds: A Challenging Journey to Bioefficacy. Br. J. Clin. Pharmacol. 2013, 75, 588–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholl, C.; Lepper, A.; Lehr, T.; Hanke, N.; Schneider, K.L.; Brockmöller, J.; Seufferlein, T.; Stingl, J.C. Population Nutrikinetics of Green Tea Extract. PLoS ONE 2018, 13, e0193074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misaka, S.; Abe, O.; Sato, H.; Ono, T.; Shikama, Y.; Onoue, S.; Yabe, H.; Kimura, J. Lack of Pharmacokinetic Interaction between Fluvastatin and Green Tea in Healthy Volunteers. Eur. J. Clin. Pharmacol. 2018, 74, 601–609. [Google Scholar] [CrossRef]
- Abe, O.; Ono, T.; Sato, H.; Müller, F.; Ogata, H.; Miura, I.; Shikama, Y.; Yabe, H.; Onoue, S.; Fromm, M.F.; et al. Role of (−)-Epigallocatechin Gallate in the Pharmacokinetic Interaction between Nadolol and Green Tea in Healthy Volunteers. Eur. J. Clin. Pharmacol. 2018, 74, 775–783. [Google Scholar] [CrossRef]
- Veerman, G.D.M.; van der Werff, S.C.; Koolen, S.L.W.; Miedema, J.R.; Oomen-de Hoop, E.; van der Mark, S.C.; Chandoesing, P.P.; de Bruijn, P.; Wijsenbeek, M.S.; Mathijssen, R.H.J. The Influence of Green Tea Extract on Nintedanib’s Bioavailability in Patients with Pulmonary Fibrosis. Biomed. Pharmacother. 2022, 151, 113101. [Google Scholar] [CrossRef]
- Misaka, S.; Ono, Y.; Taudte, R.V.; Hoier, E.; Ogata, H.; Ono, T.; König, J.; Watanabe, H.; Fromm, M.F.; Shimomura, K. Exposure of Fexofenadine, but Not Pseudoephedrine, Is Markedly Decreased by Green Tea Extract in Healthy Volunteers. Clin. Pharmacol. Ther. 2022, 112, 627–634. [Google Scholar] [CrossRef]
- Lambert, J.D.; Sang, S.; Yang, C.S. Biotransformation of Green Tea Polyphenols and the Biological Activities of Those Metabolites. Mol. Pharm. 2007, 4, 819–825. [Google Scholar] [CrossRef]
- Lee, M.-J.; Maliakal, P.; Chen, L.; Meng, X.; Bondoc, F.Y.; Prabhu, S.; Lambert, G.; Mohr, S.; Yang, C.S. Pharmacokinetics of Tea Catechins after Ingestion of Green Tea and (−)-Epigallocatechin-3-Gallate by Humans: Formation of Different Metabolites and Individual Variability. Cancer Epidemiol. Biomark. Prev. 2002, 11, 1025–1032. [Google Scholar]
- Nagle, D.G.; Ferreira, D.; Zhou, Y.-D. Epigallocatechin-3-Gallate (EGCG): Chemical and Biomedical Perspectives. Phytochemistry 2006, 67, 1849–1855. [Google Scholar] [CrossRef] [Green Version]
- Dai, W.; Ruan, C.; Zhang, Y.; Wang, J.; Han, J.; Shao, Z.; Sun, Y.; Liang, J. Bioavailability Enhancement of EGCG by Structural Modification and Nano-Delivery: A Review. J. Funct. Foods 2020, 65, 103732. [Google Scholar] [CrossRef]
- Hayashi, A.; Terasaka, S.; Nukada, Y.; Kameyama, A.; Yamane, M.; Shioi, R.; Iwashita, M.; Hashizume, K.; Morita, O. 4″-Sulfation Is the Major Metabolic Pathway of Epigallocatechin-3-Gallate in Humans: Characterization of Metabolites, Enzymatic Analysis, and Pharmacokinetic Profiling. J. Agric. Food Chem. 2022, 70, 8264–8273. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Gao, X.; Lou, Y. Interactions of Tea Polyphenols with Intestinal Microbiota and Their Implication for Cellular Signal Conditioning Mechanism. J. Food Biochem. 2019, 43, e12953. [Google Scholar] [CrossRef] [PubMed]
- Naumovski, N.; Blades, B.L.; Roach, P.D. Food Inhibits the Oral Bioavailability of the Major Green Tea Antioxidant Epigallocatechin Gallate in Humans. Antioxidants 2015, 4, 373–393. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Heber, D.; Henning, S.M. Quercetin Increased Bioavailability and Decreased Methylation of Green Tea Polyphenols in Vitro and in Vivo. Food Funct. 2012, 3, 635–642. [Google Scholar] [CrossRef] [Green Version]
- Peters, C.M.; Green, R.J.; Janle, E.M.; Ferruzzi, M.G. Formulation with Ascorbic Acid and Sucrose Modulates Catechin Bioavailability from Green Tea. Food Res. Int. 2010, 43, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Lazzeroni, M.; Guerrieri-Gonzaga, A.; Gandini, S.; Johansson, H.; Serrano, D.; Cazzaniga, M.; Aristarco, V.; Macis, D.; Mora, S.; Caldarella, P.; et al. A Presurgical Study of Lecithin Formulation of Green Tea Extract in Women with Early Breast Cancer. Cancer Prev. Res. 2017, 10, 363–370. [Google Scholar] [CrossRef] [Green Version]
- Hayasaka, S.; Goto, Y.; Maeda-Yamamoto, M. The Effects of Bathing in Hot Springs on the Absorption of Green Tea Catechin: A Pilot Study. Complement. Ther. Clin. Pract. 2013, 19, 243–245. [Google Scholar] [CrossRef]







{kind=link}
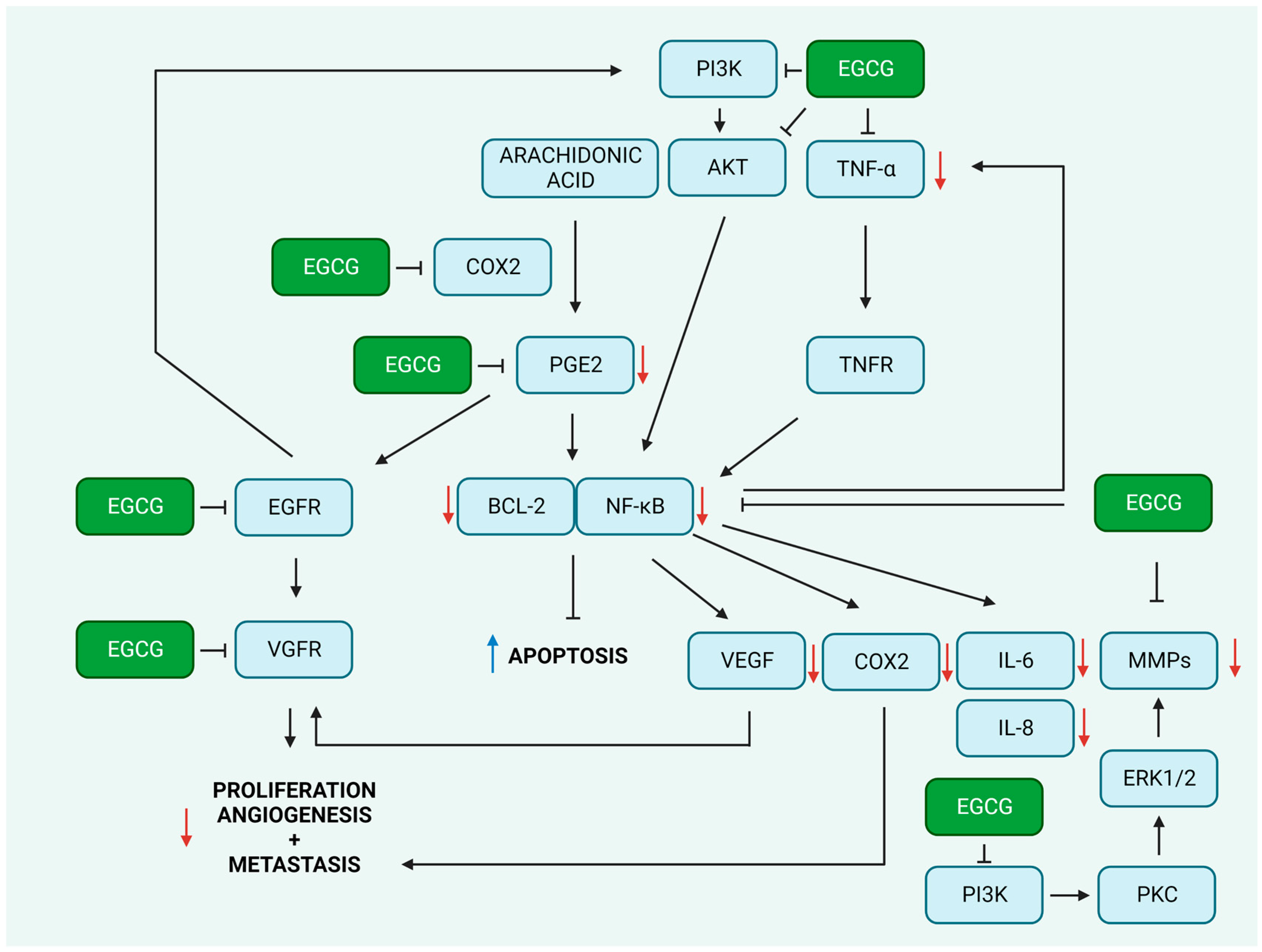{kind=link}
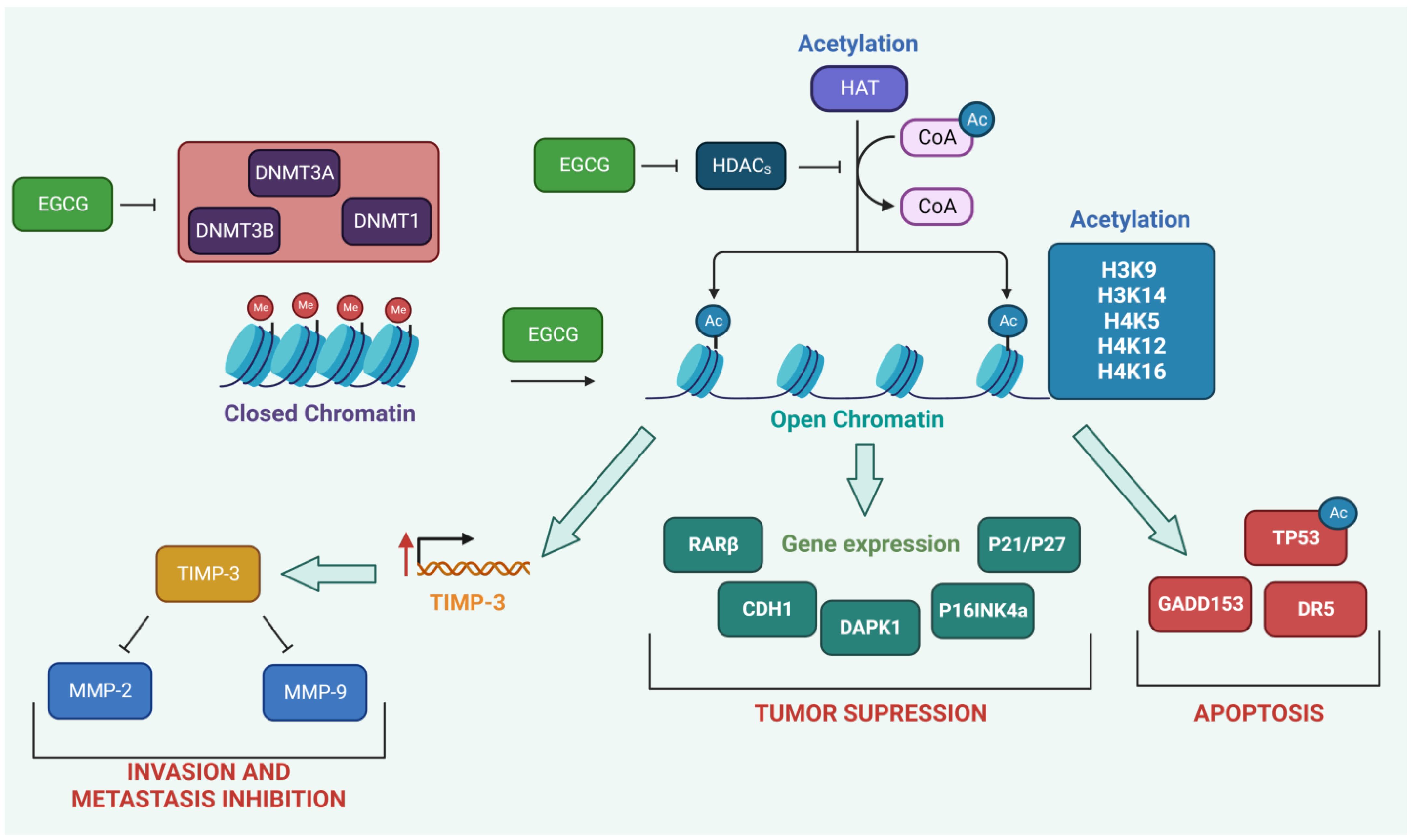{kind=link}
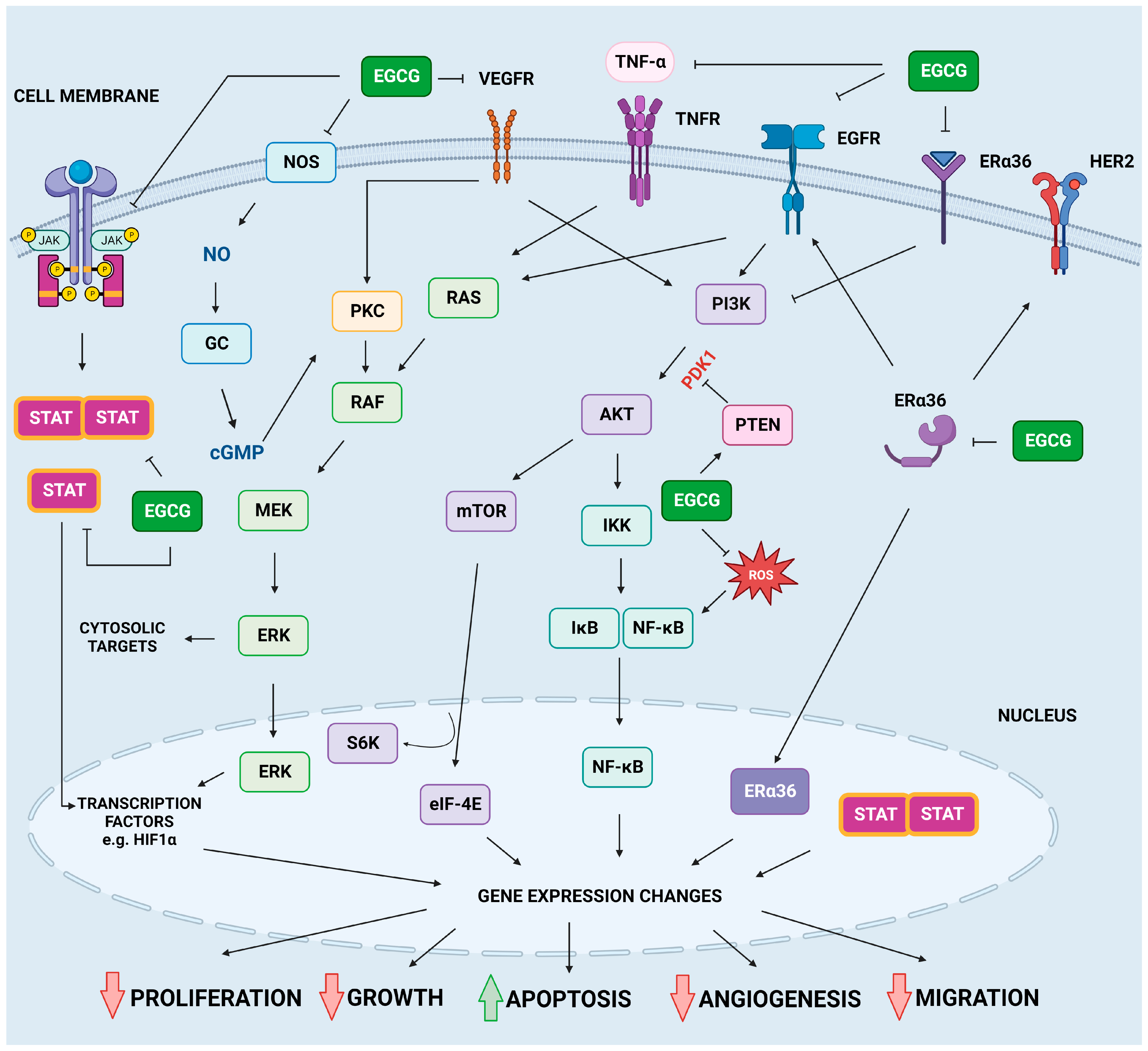{kind=link}
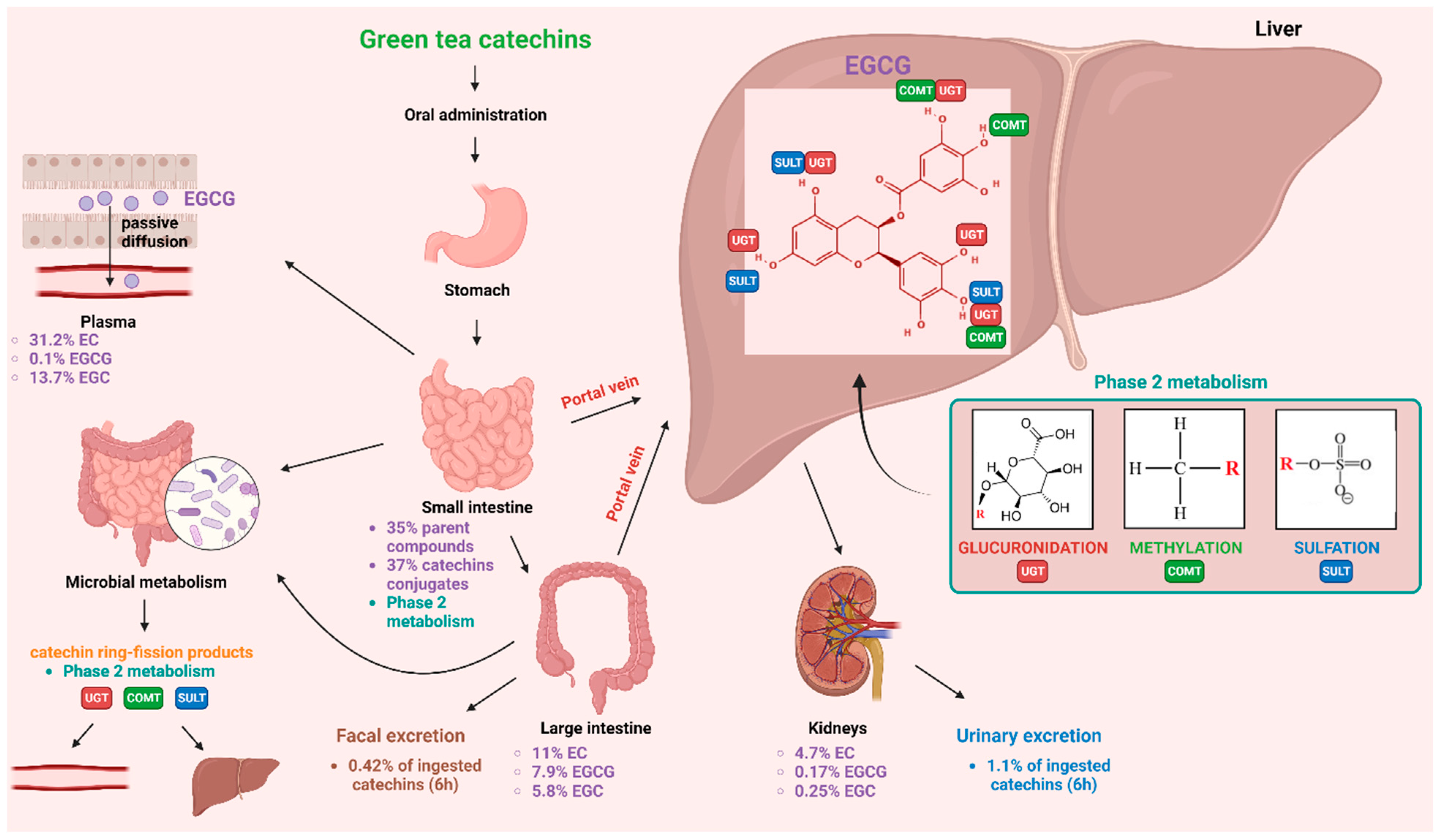{kind=link}
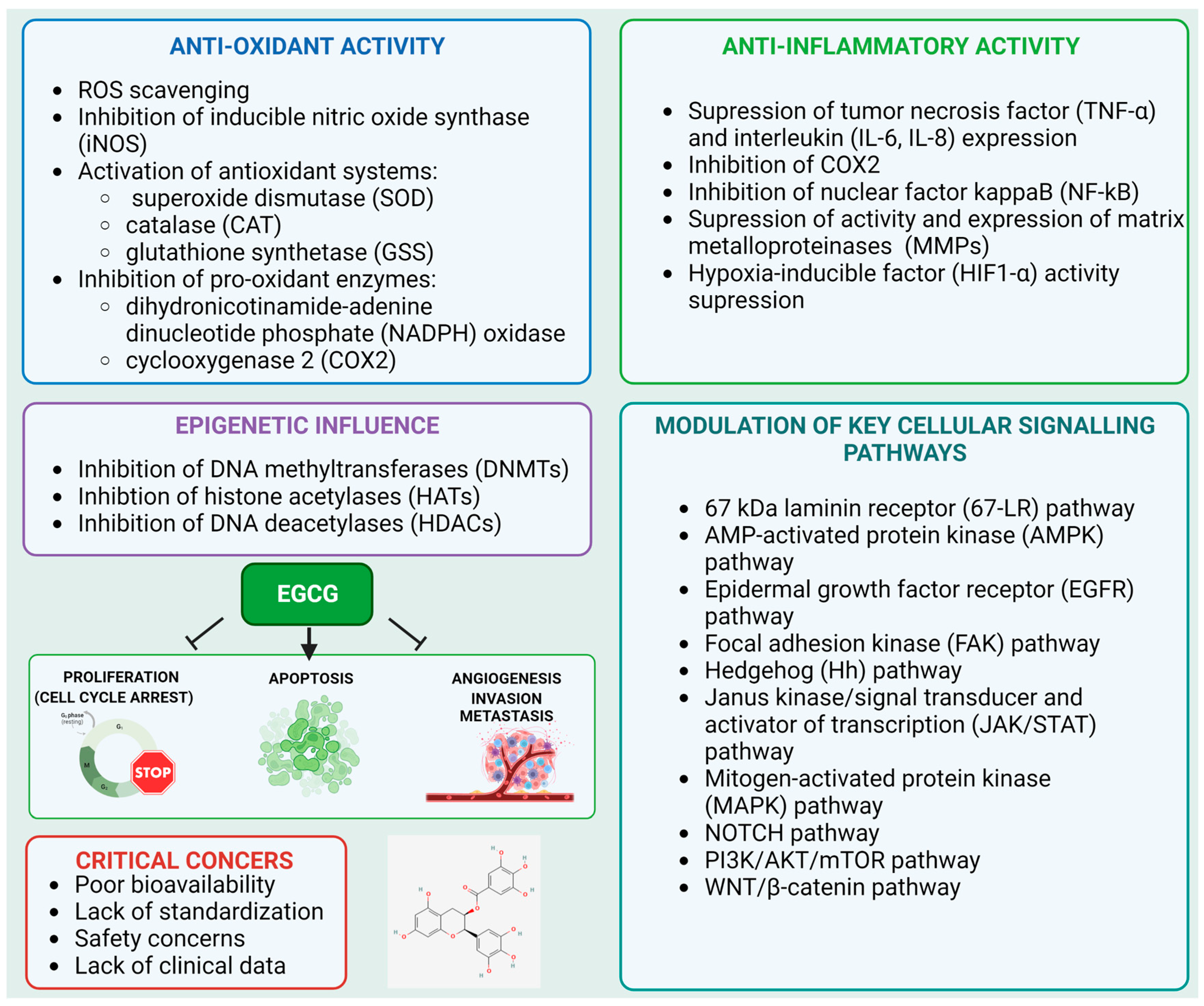{kind=link}
| Title | Number | Status | Estimated Enrollment | Objective |
|---|---|---|---|---|
| A Pilot Study to Evaluate the Chemopreventive Effects of Epigallocatechin Gallate (EGCG) in Colorectal Cancer (CRC) Patients with Curative Resections. | NCT02891538 | Recruiting | 50 | Evaluating cthe hemopreventive properties of EGCG in CRC patients with curative resections by comparing DNA methylation change during 1 year of EGCG treatment. |
| The Study of Quadruple Therapy Quercetin, Zinc, Metformin, and EGCG as Adjuvant Therapy for Early, Metastatic Breast Cancer and Triple-negative Breast Cancer, a Novel Mechanism | NCT05680662 | Not yet recruiting | 200 | Achieving the best efficacy for different stages breast cancer and triple-negative breast cancer treatment in female patients by applying various combinations of adjuvants. |
| Study of Epigallocatechin-3-gallate (EGCG) for Supportive and Symptomatic Management in Patients with Esophageal Cancer | NCT05039983 | Recruiting | 15 | Assessing the application of EGCG for alleviating esophageal stenosis, choking and pain while swallowing, weight change, and blood parameter changes in pathological esophageal squamous cell carcinoma patients. |
| Reducing Frailty for Older Cancer Survivors Using Supplements (ReFOCUS): A Phase 2 Randomized Controlled Trial of Epigallocatechin-3-Gallate (EGCG) on Frailty and Inflammation in Older Survivors of Cancer | NCT04553666 | Recruiting | 40 | Investigating the beneficial properties and safety of EGCG treatment in older cancer survivors with frailty and inflammation. |
| A Phase II Randomized Double Blinded Study of Green Tea Catechins (GTC) vs. Placebo in Men on Active Surveillance for Prostate Cancer: Modulation of Biological and Clinical Intermediate Biomarkers | NCT04597359 | Recruiting | 360 | Assessing the abilities of tea catechins in preventing the progression of prostate cancer using various genetic, biochemical, and clinical markers. |
| Phase II Clinical Trial of Green Tea Catechins in Men on Active Surveillance (AS) | NCT04300855 | Recruiting | 135 | An evaluation of the bioavailability, safety, effectiveness, and validation of the acting mechanism of a drug containing EGCG applied for 24 months to men with adenocarcinoma of the prostate with cancer present. |
| Pioneering Pre- and Post-Operative Integrative Care to Improve Thoracic Cancer Quality of Care—The Thoracic Peri-Operative Integrative Surgical Care Evaluation (POISE) Trial—Stage II | NCT04871412 | Recruiting | 20 | Improving health-related quality of life, decreasing surgical adverse events, prolonging overall survival, and pioneering integrative care delivery for thoracic cancer patients. |
| Fibroids and Unexplained Infertility Treatment With Epigallocatechin Gallate; A Natural CompounD in Green Tea (FRIEND) | NCT05364008 | Recruiting | 200 | Determining the effect of low-caffeine green tea extract containing 45% EGCG on fibroids and subsequent pregnancy and live births in women seeking fertility treatment. |
| The Use of Vitamin D in Combination With Epigallocatechin Gallate, D-chiro-inositol and Vitamin B6 in the Treatment of Women With Uterine Fibroid | NCT05448365 | Recruiting | 60 | Evaluating the impact on uterine fibroid volume of a combination of natural molecules including epigallocatechin gallate, vitamin D, D-chiro-inositol, and vitamin B6. |
| Effects of Vitamin D, Epigallocatechin Gallate, Vitamin B6, and D-Chiro-inositol Combination on Uterine Fibroids: a Randomized Controlled Trial | NCT05409872 | Recruiting | 108 | Evaluating the efficacy of a combination of epigallocatechin gallate, vitamin D3, D-chiro-inositol, and vitamin B6 as a treatment for uterine fibroids. |
| A Phase I Single-arm, Multicenter Pilot Study Aimed at Validating γ-OHPdG as a Biomarker and Testing the Effects of Polyphenon E on Its Levels in Patients With Cirrhosis | NCT03278925 | Active, not recruiting | 48 | Investigating the side effects and best dose of defined green tea catechin extract and verifying its ability to prevent liver cancer in participants with cirrhosis. |
| A Pilot Study of Gemcitabine, Abraxane, Metformin and a Standardized Dietary Supplement (DS) in Patients With Unresectable Pancreatic Cancer | NCT02336087 | Active, not recruiting | 21 | Investigating the side effects of gemcitabine hydrochloride, nab-paclitaxel, metformin hydrochloride, and a standardized dietary supplement including EGCG in treating patients with pancreatic cancer that cannot be removed by surgery. |
| Phase ⅠStudy of Oral Green Tea Extract as Maintenance Therapy for Extensive-stage Small Cell Lung Cancer | NCT01317953 | Available | No information | Assessing the safety of the application of EGCG for extensive-stage small lung cancer patients who have achieved an objective tumor response after first-line therapy. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kciuk, M.; Alam, M.; Ali, N.; Rashid, S.; Głowacka, P.; Sundaraj, R.; Celik, I.; Yahya, E.B.; Dubey, A.; Zerroug, E.; et al. Epigallocatechin-3-Gallate Therapeutic Potential in Cancer: Mechanism of Action and Clinical Implications. Molecules 2023, 28, 5246. https://doi.org/10.3390/molecules28135246
Kciuk M, Alam M, Ali N, Rashid S, Głowacka P, Sundaraj R, Celik I, Yahya EB, Dubey A, Zerroug E, et al. Epigallocatechin-3-Gallate Therapeutic Potential in Cancer: Mechanism of Action and Clinical Implications. Molecules. 2023; 28(13):5246. https://doi.org/10.3390/molecules28135246
Chicago/Turabian StyleKciuk, Mateusz, Manzar Alam, Nemat Ali, Summya Rashid, Pola Głowacka, Rajamanikandan Sundaraj, Ismail Celik, Esam Bashir Yahya, Amit Dubey, Enfale Zerroug, and et al. 2023. "Epigallocatechin-3-Gallate Therapeutic Potential in Cancer: Mechanism of Action and Clinical Implications" Molecules 28, no. 13: 5246. https://doi.org/10.3390/molecules28135246