Author Contributions
Conceptualization, J.Y.Y. and C.B.-F.; methodology, C.B.-F. and J.Y.Y.; software, J.Y.Y., A.M., R.F. and H.K.; validation, C.B.-F., A.M. and R.F.; formal analysis, C.B.-F., A.M., R.F. and H.K.; investigation, C.B.-F.; resources, J.Y.Y.; data curation, C.B.-F. and E.A.; writing—original draft preparation, C.B.-F. and H.K.; writing—review and editing, J.Y.Y., E.A., A.M. and R.F.; visualization, C.B.-F. and J.Y.Y.; supervision, J.Y.Y.; project administration, J.Y.Y. and E.A. All authors have read and agreed to the published version of the manuscript.
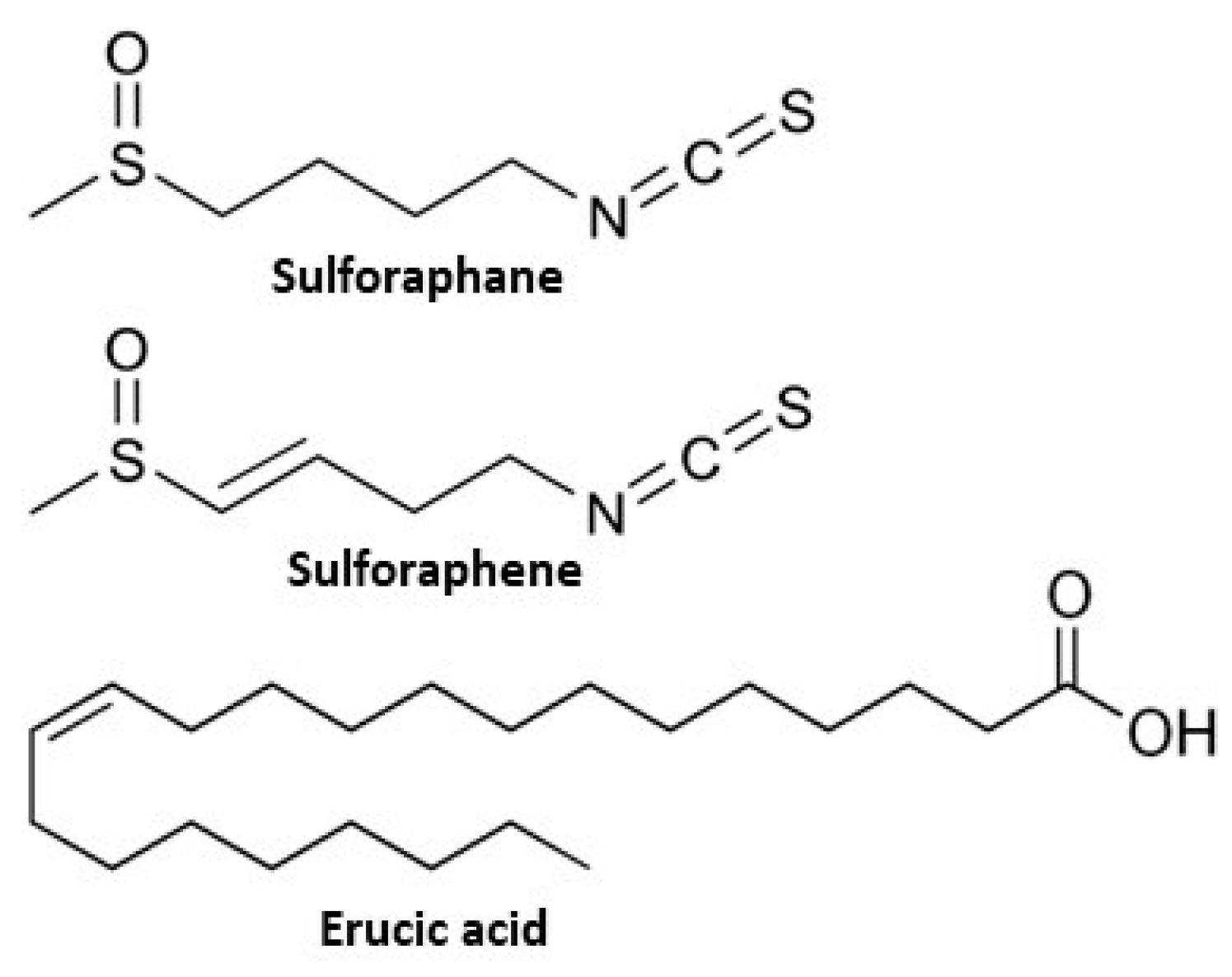
Figure 1.
Structures of sulforaphane, sulforaphene, and erucic acid.
Figure 1.
Structures of sulforaphane, sulforaphene, and erucic acid.
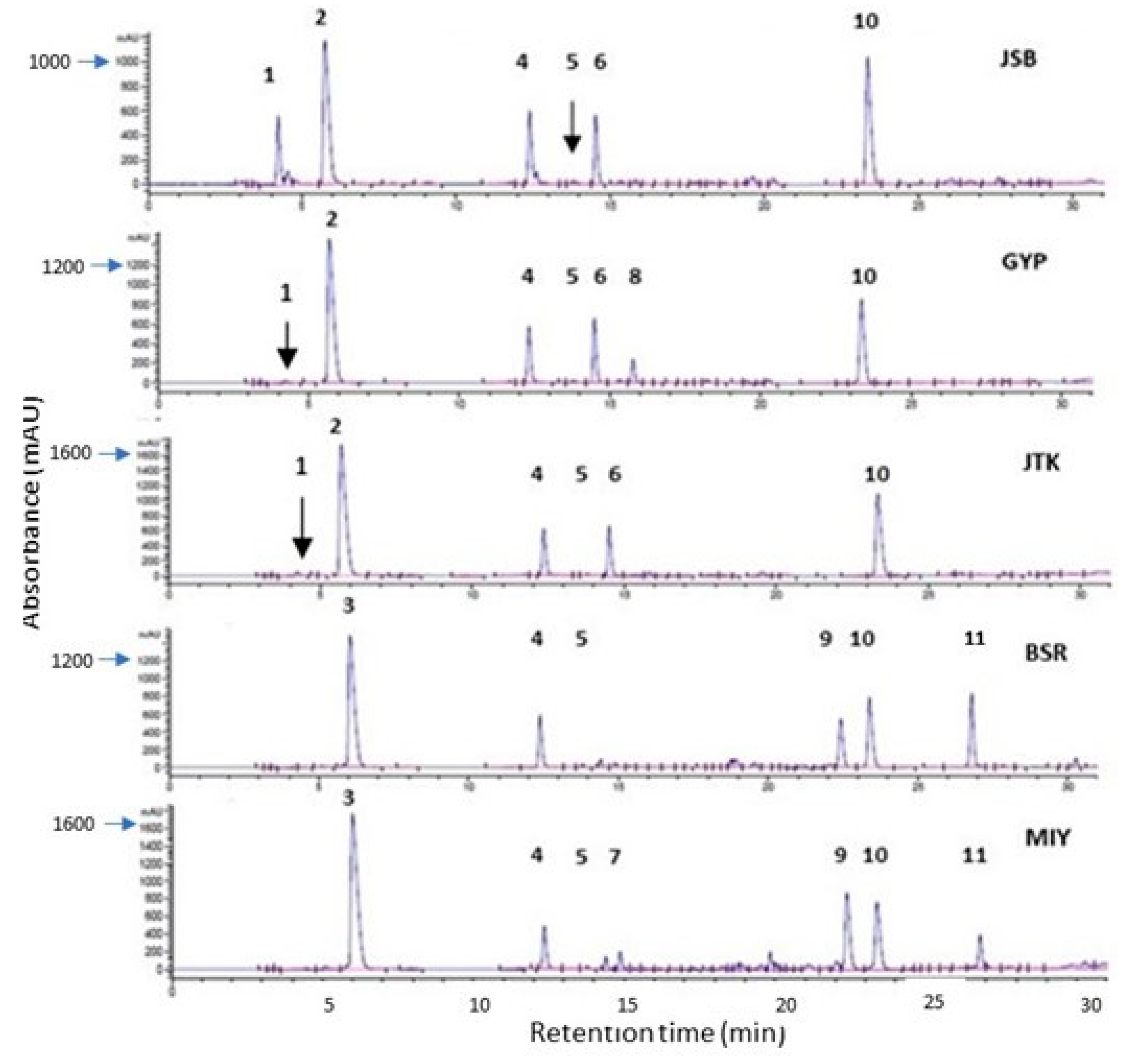
Figure 2.
Chromatograms of selected one-day-old broccoli, kale, and broccoli cultivars (227 nm) showing glucosinolates and phenolic compounds. Broccoli: Johnny’s Sprouting Broccoli (JSB), Gypsy F1 (GYP); Kale: Johnny’s Toscano Kale (JTK); Radish: Black Spanish Round (BSR), and Miyashige (MIY). Compounds: 1: Glucoiberin; 2: glucoraphanin; 3: glucoraphenin; 4: 4-hydroxyglucobrassicin; 5: glucoiberverin; 6: glucoerucin; 7: glucoraphasatin; 8: glucobrassicin; 9–11: Unidentified phenolics.
Figure 2.
Chromatograms of selected one-day-old broccoli, kale, and broccoli cultivars (227 nm) showing glucosinolates and phenolic compounds. Broccoli: Johnny’s Sprouting Broccoli (JSB), Gypsy F1 (GYP); Kale: Johnny’s Toscano Kale (JTK); Radish: Black Spanish Round (BSR), and Miyashige (MIY). Compounds: 1: Glucoiberin; 2: glucoraphanin; 3: glucoraphenin; 4: 4-hydroxyglucobrassicin; 5: glucoiberverin; 6: glucoerucin; 7: glucoraphasatin; 8: glucobrassicin; 9–11: Unidentified phenolics.
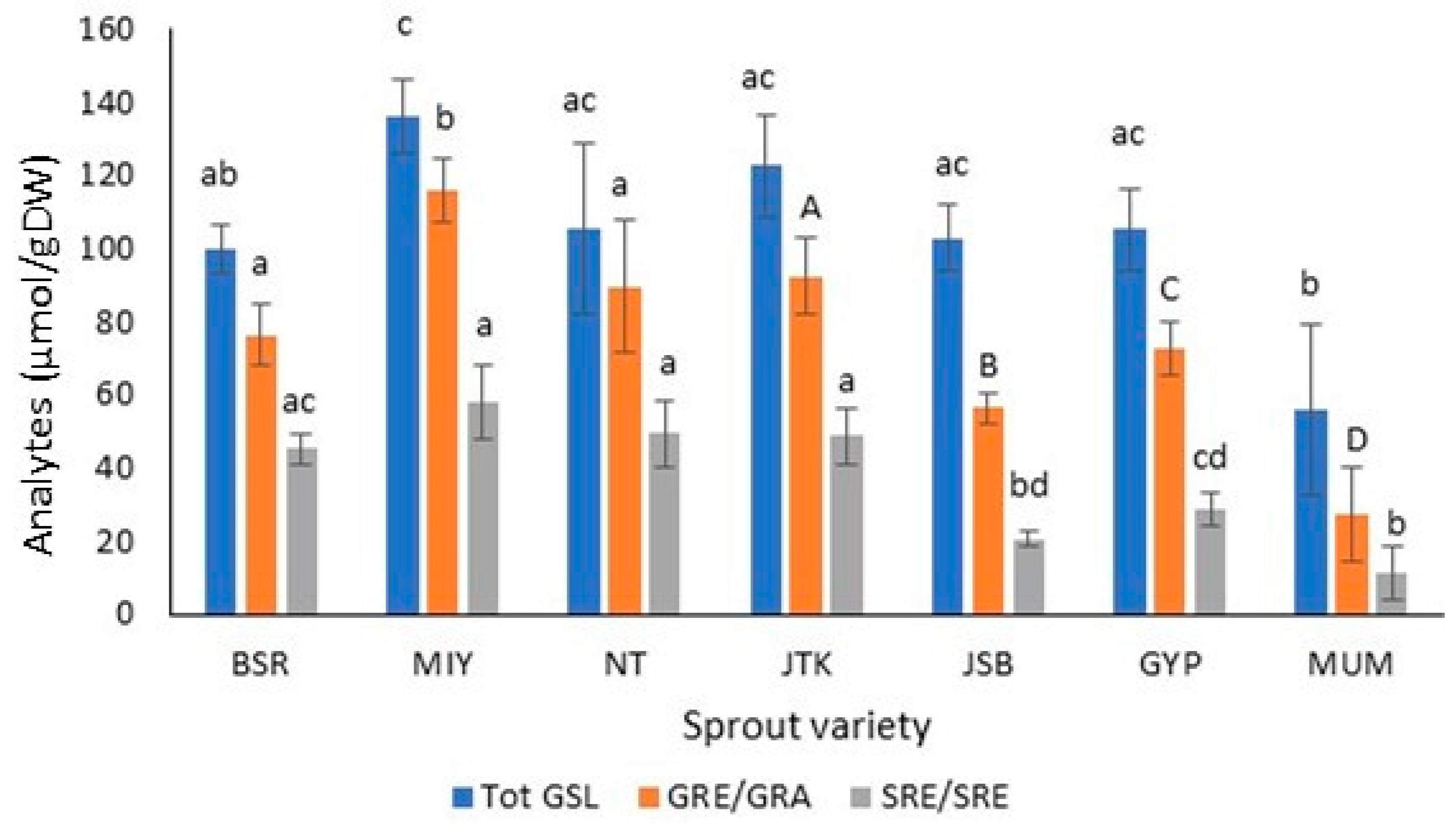
Figure 3.
Total glucosinolates (Tot GSL), glucoraphenin (GRE), glucoraphanin (GRA), sulforaphene (SFE), and sulforaphane (SFA) levels in dark-grown radish, kale, and broccoli cultivars. (For the ITC of JSB, light-grown sprouts are used.) Broccoli: Johnny’s Sprouting Broccoli (JSB), Gypsy F1 (GYP) Mumm’s Sprouting Broccoli (MUM); Kale: Johnny’s Toscano Kale (JTK); Radish: Black Spanish Round (BSR), and Miyashige (MIY), Nero Tunda (NT). GRE and SRE are in radishes only; GRA and SFA are in kale and broccoli (n = 3; ±SD). For a given analyte type, values followed by the same letter and font type are not significantly different (p ≤ 0.05). Upper-case letters are used for GRA only.
Figure 3.
Total glucosinolates (Tot GSL), glucoraphenin (GRE), glucoraphanin (GRA), sulforaphene (SFE), and sulforaphane (SFA) levels in dark-grown radish, kale, and broccoli cultivars. (For the ITC of JSB, light-grown sprouts are used.) Broccoli: Johnny’s Sprouting Broccoli (JSB), Gypsy F1 (GYP) Mumm’s Sprouting Broccoli (MUM); Kale: Johnny’s Toscano Kale (JTK); Radish: Black Spanish Round (BSR), and Miyashige (MIY), Nero Tunda (NT). GRE and SRE are in radishes only; GRA and SFA are in kale and broccoli (n = 3; ±SD). For a given analyte type, values followed by the same letter and font type are not significantly different (p ≤ 0.05). Upper-case letters are used for GRA only.
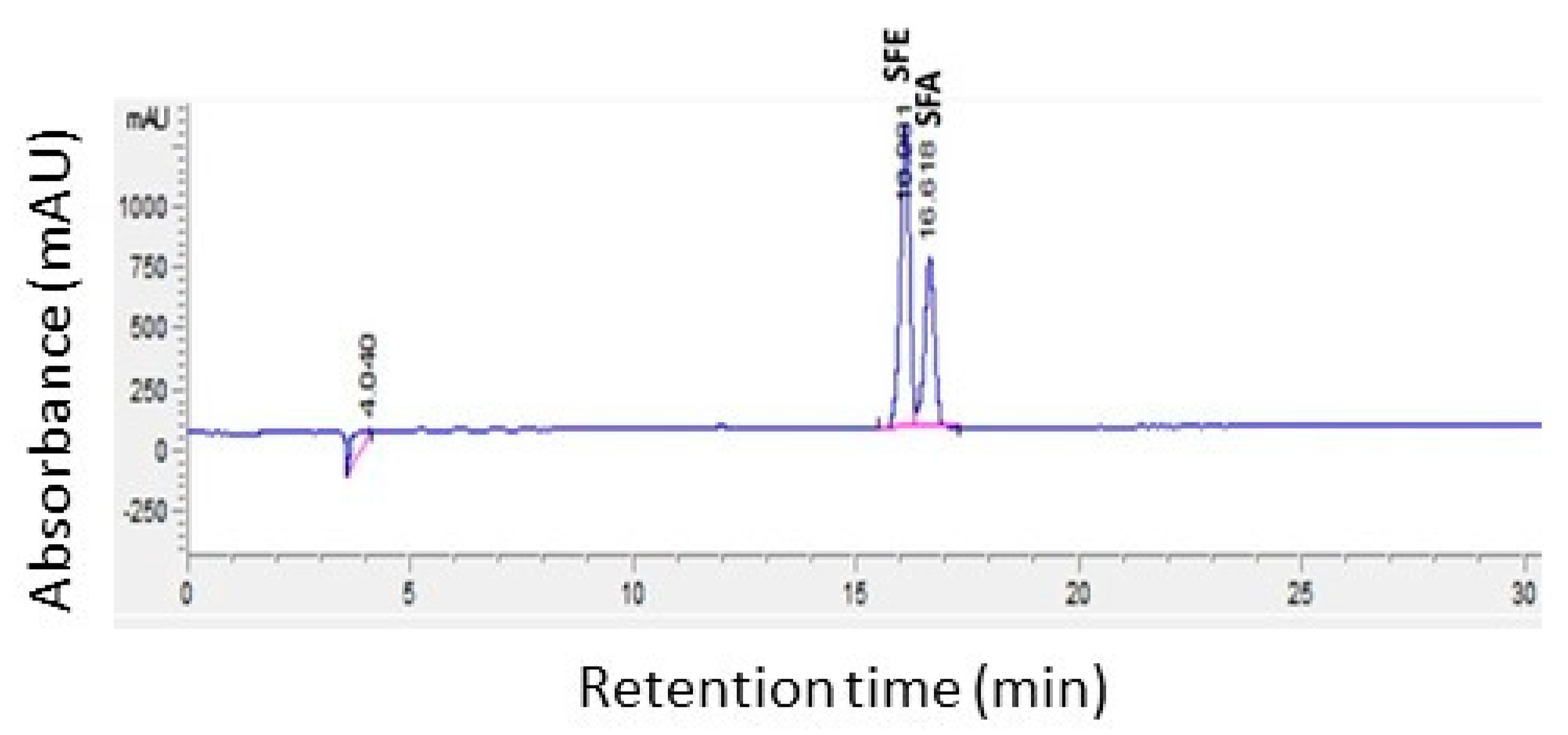
Figure 4.
HPLC Chromatogram of a mixture of sulforaphene (SFE: tR16.08 s) and sulforaphane (SFA: tR16.62 s) at 205 nm.
Figure 4.
HPLC Chromatogram of a mixture of sulforaphene (SFE: tR16.08 s) and sulforaphane (SFA: tR16.62 s) at 205 nm.
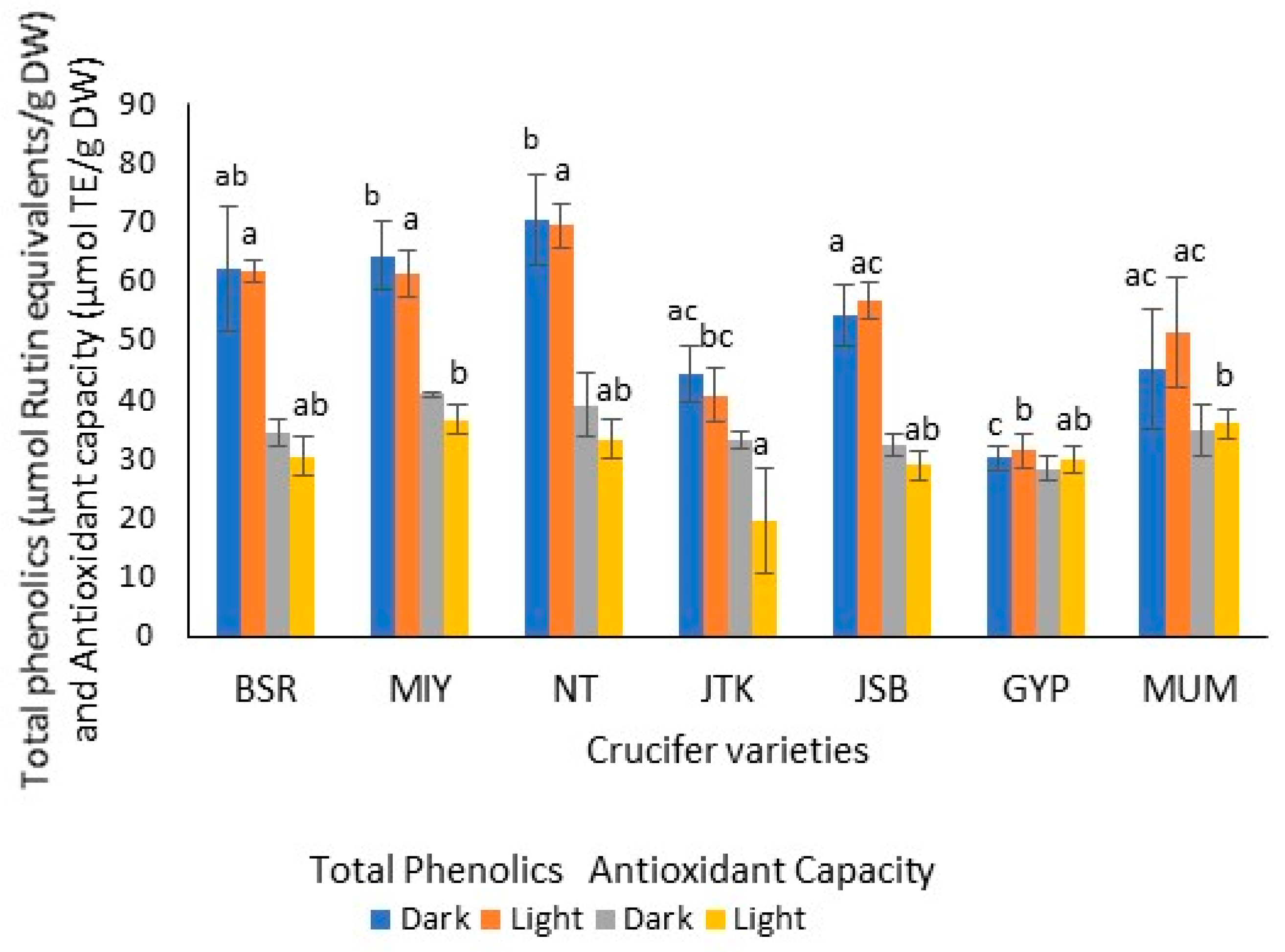
Figure 5.
Total phenolics (µmol Rutin equivalents/g DW) and antioxidant capacity (DPPH radical scavenging activity: µmol Trolox equivalents/g DW) of one-day-old sprout extracts grown under dark and light conditions. Radish: Black Spanish Round (BSR), Miyashige (MIY), and Nero Tunda (NT); kale: Johnny’s Toscano Kale (JTK); broccoli: Johnny’s Sprouting Broccoli (JSB), Gypsy F1 (GYP), and Mumm’s sprouting broccoli (MUM). (n = 3; ±SD). For a given analyte and lighting condition, values followed by the same letter are not significantly different (p ≤ 0.05). For all the sprout varieties, there was no significant difference observed in the analyte levels for different lighting conditions.
Figure 5.
Total phenolics (µmol Rutin equivalents/g DW) and antioxidant capacity (DPPH radical scavenging activity: µmol Trolox equivalents/g DW) of one-day-old sprout extracts grown under dark and light conditions. Radish: Black Spanish Round (BSR), Miyashige (MIY), and Nero Tunda (NT); kale: Johnny’s Toscano Kale (JTK); broccoli: Johnny’s Sprouting Broccoli (JSB), Gypsy F1 (GYP), and Mumm’s sprouting broccoli (MUM). (n = 3; ±SD). For a given analyte and lighting condition, values followed by the same letter are not significantly different (p ≤ 0.05). For all the sprout varieties, there was no significant difference observed in the analyte levels for different lighting conditions.
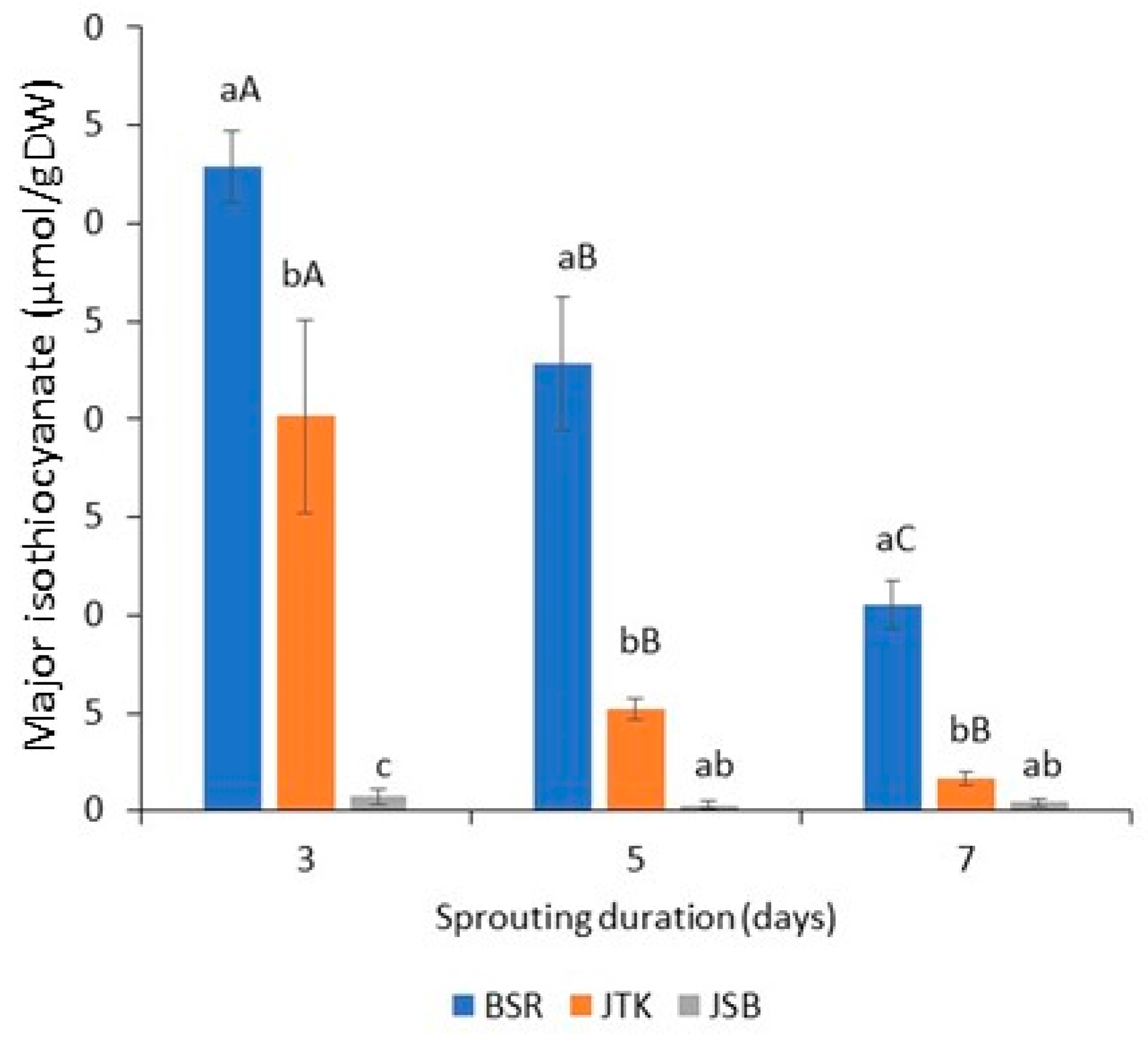
Figure 6.
Content of major isothiocyanate in three-, five-, and seven-day radish, kale, and broccoli cultivars: sulforaphene is found in Black Spanish Round (BSR) radishes, and sulforaphane is found in Johnny’s Toscano Kale (JTK) and Johnny’s Sprouting Broccoli (JSB); (n = 3; ±SD). Different lowercase and upper-case letters above bars, respectively, indicate significant differences for samples of the same sampling day and samples of the same variety at p < 0.05, according to Tukey’s Multiple Comparison Test.
Figure 6.
Content of major isothiocyanate in three-, five-, and seven-day radish, kale, and broccoli cultivars: sulforaphene is found in Black Spanish Round (BSR) radishes, and sulforaphane is found in Johnny’s Toscano Kale (JTK) and Johnny’s Sprouting Broccoli (JSB); (n = 3; ±SD). Different lowercase and upper-case letters above bars, respectively, indicate significant differences for samples of the same sampling day and samples of the same variety at p < 0.05, according to Tukey’s Multiple Comparison Test.
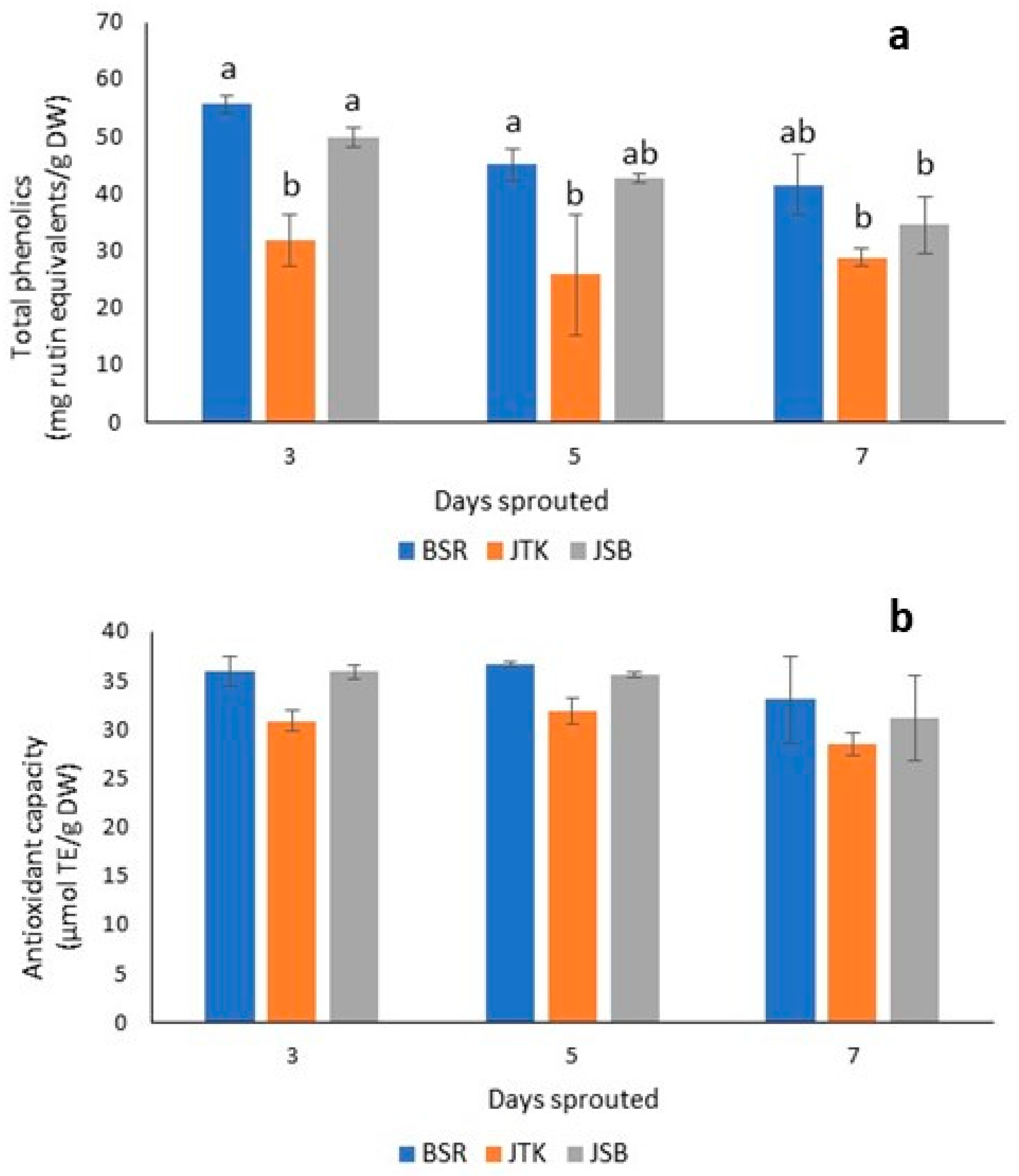
Figure 7.
Total phenolics (uM rutin equivalents/g DW) (a) and DPPH radical scavenging activity (µmol TE/g DW) (b) of three-, five-, and seven-day-old sprout extracts. Black Spanish Round radish (BSR); Johnny’s Toscano Kale (JTK); Johnny’s Sprouting Broccoli (JSB). Different letters above bars indicate significant differences at p < 0.05 according to Tukey’s Multiple Comparison Test.
Figure 7.
Total phenolics (uM rutin equivalents/g DW) (a) and DPPH radical scavenging activity (µmol TE/g DW) (b) of three-, five-, and seven-day-old sprout extracts. Black Spanish Round radish (BSR); Johnny’s Toscano Kale (JTK); Johnny’s Sprouting Broccoli (JSB). Different letters above bars indicate significant differences at p < 0.05 according to Tukey’s Multiple Comparison Test.
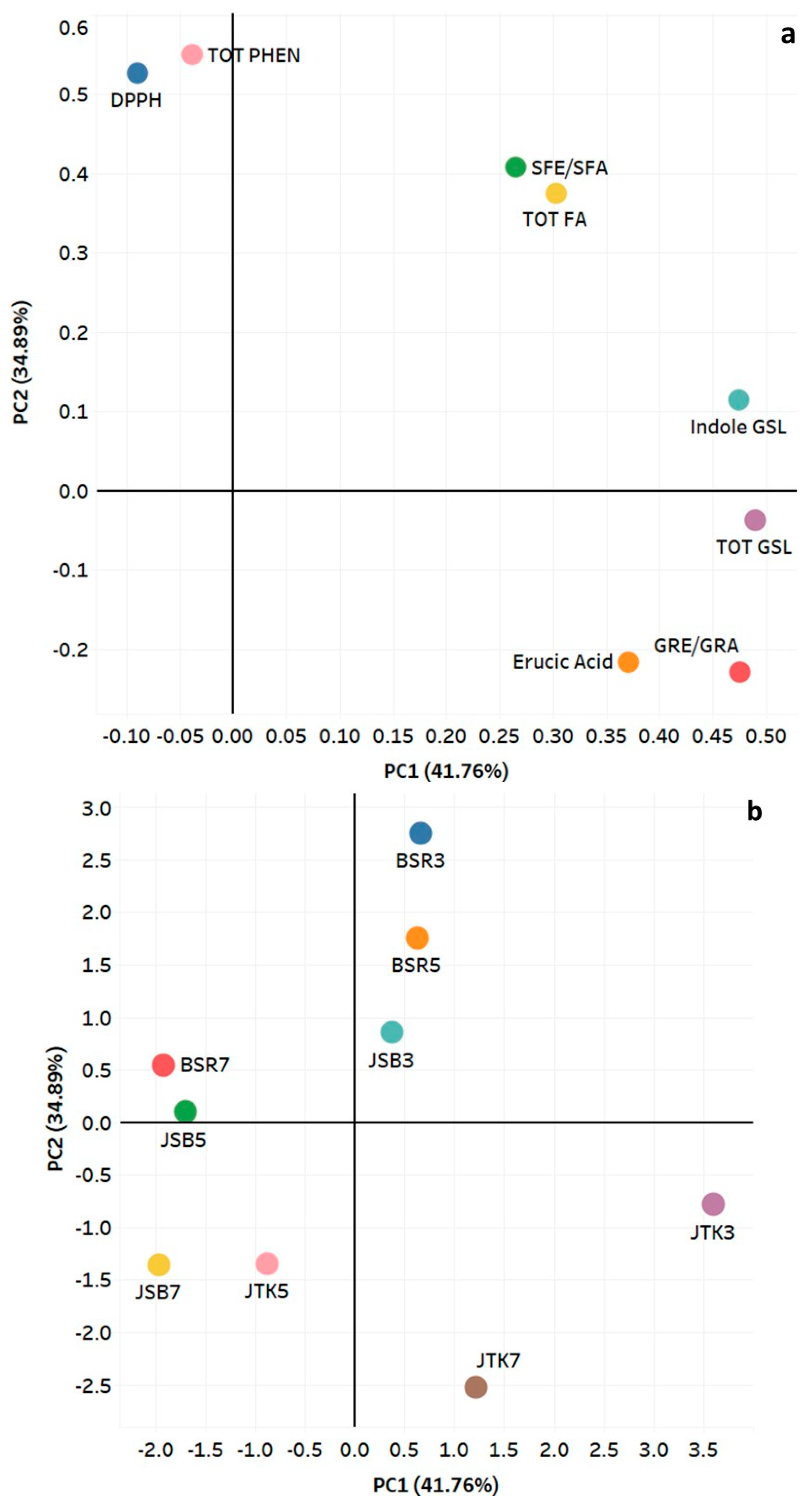
Figure 8.
Principal component analysis showing: (a) correlation of phytochemical parameters with PC1 and PC2, and (b) relationship of three cruciferous cultivars at three harvest dates. PCA was performed on the correlation matrix of average values of phytochemical parameters and antioxidant capacity (DPPH).
Figure 8.
Principal component analysis showing: (a) correlation of phytochemical parameters with PC1 and PC2, and (b) relationship of three cruciferous cultivars at three harvest dates. PCA was performed on the correlation matrix of average values of phytochemical parameters and antioxidant capacity (DPPH).
Table 1.
Glucosinolate contents in one-day-old radish, kale and broccoli sprouts grown under dark and light conditions *.
Table 1.
Glucosinolate contents in one-day-old radish, kale and broccoli sprouts grown under dark and light conditions *.
| | GIB | GRA | GRE | 4OHGBR | GIV | GER | GRPH | GBR | Tot. GSL | Total Indole GSL |
|---|
| Retention time | 4.22 | 5.65 | 5.92 | 12.26 | 13.70 | 14.49 | 14.82 | 15.77 | | |
| Dark | | | | | | | | | | |
| Black Spanish Round | ND | ND | 76.31 ± 8.38 a | 5.03 ± 0.34 | 2.73 ± 0.49 ab | ND | 5.85 ± 2.21 | ND | 89.91 ± 6.83 ab | 5.03 ± 0.34 |
| Miyashige | ND | ND | 116.15 ± 8.90 b | 5.00 ± 0.59 | 2.96 ± 0.66 ab | ND | 12.10 ± 1.61 | ND | 136.21 ± 9.97 c | 5.00 ± 0.59 |
| Nero Tunda | ND | ND | 89.70 ± 18.16 a | 4.88 ± 1.21 | 3.37 ± 0.42 a | ND | 7.62 ± 4.34 | ND | 105.57 ± 23.46 ac | 4.88 ± 1.22 |
| Toscano Kale | 3.96 ± 0.62 a | 92.57 ± 10.61 a | ND | 5.14 ± 0.81 | 1.96 ± 0.77 ab | 18.53 ± 1.9 a | ND | 0.61 ± 0.12 a | 122.77 ± 13.99 ac | 5.75 ± 0.82 |
| Johnny’s Sprouting Broccoli | 22.22 ± 2.03 b | 59.08 ± 3.46 b | ND | 5.91 ± 0.62 | 1.91 ± 0.30 ab | 18.15 ± 2.37 a | ND | 0.66 ± 0.04 a | 107.92 ± 4.51 ac | 6.57 ± 0.66 |
| Gypsy Broccoli | 3.00 ± 3.72 a | 68.88 ± 3.72 c | ND | 4.78 ± 0.81 | 1.53 ± 0.05 ab | 19.73 ± 1.867 ac | ND | 1.89 ± 0.32 b | 99.8 ± 6.17 ac | 6.67 ± 1.12 |
| Mumm’s Sprouting Broccoli | 13.03 ± 4.95 c | 27.59 ± 12.86 d | ND | 4.42 ± 1.98 | 1.97 ± 0.55 bc | 8.45 ± 3.94 bc | ND | 0.64 ± 0.16 aA | 56.11 ± 23.35 b | 5.07 ± 2.13 |
| Light | | | | | | | | | | |
| Black Spanish Round | ND | ND | 79.21 ± 2.17 a | 4.93 ± 0.39 | 1.9 ± 0.55 | ND | 3.54 ± 0.98 | ND | 89.81 ± 3.97 ac | 4.93 ± 0.39 |
| Miyashige | ND | ND | 110.34 ± 6.88 b | 4.74 ± 0.69 | 1.94 ± 046 | ND | 8.93 ± 2.80 | ND | 126.18 ± 11.08 bc | 4.74 ± 0.69 |
| Nero Tunda | ND | ND | 97.02 ± 2.58 ab | 4.57 ± 0.67 | 2.07 ± 1.04 | ND | 4.19 ± 1.48 | ND | 108.07 ± 4.00 bc | 4.57 ± 0.67 |
| Tuscan Kale | 3.69 ± 0.58 a | 91.18 ± 8.44 a | ND | 5.04 ± 0.94 | 1.61 ± 0.03 | 19.01 ± 0.78 a | ND | 0.65 ± 0.14 a | 119.87 ± 11.92 bc | 5.68 ± 0.95 |
| Johnny’s Sprouting Broccoli | 21.51 ± 1.28 b | 56.47 ± 4.36 b | ND | 5.82 ± 0.71 | 1.65 ± 0.26 | 16.86 ± 1.27 ac | ND | 0.65 ± 0.06 a | 102.97 ± 8.91 ac | 6.47 ± 0.76 |
| Gypsy Broccoli | 2.93 ± 0.26 c | 72.97 ± 7.46 c | ND | 4.98 ± 0.81 | 1.66 ± 0.08 | 20.70 ± 2.89 a | ND | 2.09 ± 0.18 b | 105.33 ± 11.29 ac | 7.07 ± 0.89 |
| Mumm’s Sprouting Broccoli | 16.48 ± 3.35 d | 35.54 ± 4.58 d | ND | 5.73 ± 1.04 | 2.18 ± 0.99 | 10.56 ± 1.73 bc | ND | 0.80 ± 0.08 aB | 71.29 ± 10.01 a | 6.53 ± 1.12 |
Table 2.
Pearson’s correlation coefficients among selected analyzed parameters in crucifer cultivars.
Table 2.
Pearson’s correlation coefficients among selected analyzed parameters in crucifer cultivars.
| | Tot GSL | GRE/GRA | Indole GSL | SFE/SFA | Phenolics | AOC a | Erucic Acid |
|---|
| Tot GSL | 1.00 | | | | | | |
| GRE/GRA | 0.94 | 1.00 | | | | | |
| Indole GSL | 0.09 | −0.19 | 1.00 | | | | |
| SFE/SFA | 0.82 | 0.95 | −0.42 | 1.00 | | | |
| Phenolics | 0.26 | 0.4 | −0.71 | 0.52 | 1.00 | | |
| AOC a | −0.2 | 0.06 | −0.72 | 0.15 | 0.61 | 1.00 | |
| Erucic acid | −0.37 | −0.56 | 0.51 | −0.68 | −0.76 | −0.43 | 1.00 |
Table 3.
Fatty acid composition of one-day-old radish, kale, and broccoli cultivars grown under dark and light conditions a.
Table 3.
Fatty acid composition of one-day-old radish, kale, and broccoli cultivars grown under dark and light conditions a.
| Fatty Acid | Sprout Variety |
|---|
| Dark | BSR | MIY | NT | JTK | JSB | GYP | MUM |
|---|
| 16:0 | 6.37 ± 0.28 | 6.15 ± 0.16 | 6.55 ± 0.20 | 5.63 ± 0.07 | 4.51 ± 0.11 | 4.54 ± 0.14 | 4.64 ± 0.41 |
| 18:0 | 1.41 ± 0.38 | 1.62 ± 0.04 | 1.48 ± 0.13 | 0.56 ± 0.04 | 0.28 ± 0.09 | 0.56 ± 0.06 | 0.30 ± 0.03 |
| 18:1c9 | 28.68 ± 0.65 | 28.39 ± 0.65 | 28.97 ± 1.11 | 11.00 ± 0.25 | 13.41 ± 0.51 | 18.80 ± 0.17 | 14.91 ± 0.06 |
| 18:1c7 | 1.05 ± 0.37 | 0.88 ± 0.09 | 1.38 ± 0.11 | 0.74 ± 0.10 | 0.90 ± 0.07 | 1.29 ± 0.13 | 0.78 ± 0.07 |
| 18:2 | 12.14 ± 0.26 | 10.78 ± 0.66 | 11.43 ± 0.29 | 16.84 ± 0.31 | 15.62 ± 0.56 | 15.40 ± 0.15 | 13.39 ± 0.55 |
| 20:0 | 0.63 ± 0.03 | 0.78 ± 0.04 | 0.62 ± 0.13 | 0.07 ± 0.03 | ND | 0.04 ± 0.05 | 0.00 |
| 20:1ω15 | 9.83 ± 0.31 | 10.15 ± 0.15 | 10.19 ± 0.13 | 3.99 ± 0.07 | 4.54 ± 0.30 | 7.96 ± 0.03 | 3.88 ± 0.04 |
| 18:3ω3 | 13.48 ± 0.51 | 9.18 ± 0.43 | 13.54 ± 0.29 | 12.30 ± 0.57 | 13.29 ± 0.68 | 11.87 ± 0.42 | 12.34 ± 0.28 |
| 20:2 | 0.12 ± 0.02 | 0.18 ± 0.04 | 0.00 | 0.32 ± 0.04 | 0.12 ± 0.09 | 0.25 ± 0.05 | ND |
| 22:0 | 0.26 ± 0.03 | 0.55 ± 0.04 | 0.26 ± 0.15 | 0.20 ± 0.05 | 0.19 ± 0.10 | ND | ND |
| 22:1 | 25.22 ± 0.42 | 30.80 ± 0.80 | 24.76 ± 1.02 | 47.41 ± 0.81 | 45.83 ± 1.01 | 39.16 ± 1.01 | 49.44 ± 1.32 |
| 22:2 | ND | ND | ND | 0.96 ± 0.04 | 0.55 ± 0.08 | 0.10 ± 0.07 | 0.31 ± 0.04 |
| 24:1 | 0.66 ± 0.34 | 0.81 ± 0.01 | 0.82 ± 0.11 | ND | ND | ND | ND |
| Saturated | 8.83 ± 0.33 | 9.10 ± 0.19 | 8.91 ± 0.31 | 6.45 ± 0.18 | 4.99 ± 0.06 | 5.16 ± 0.16 | 4.94 ± 0.44 |
| MUFA | 65.43 ± 0.40 | 70.76 ± 1.13 | 66.11 ± 0.45 | 63.13 ± 0.61 | 65.43 ± 1.15 | 67.22 ± 0.74 | 69.02 ± 1.23 |
| PUFA | 25.74 ± 0.71 | 20.14 ± 1.06 | 24.98 ± 0.15 | 30.42 ± 0.67 | 29.58 ± 1.13 | 27.62 ± 0.67 | 26.04 ± 0.79 |
| Light | | | | | | | |
| 16:00 | 6.35 ± 0.29 | 5.98 ± 0.31 | 6.36 ± 0.2 | 5.16 ± 0.21 | 4.55 ± 0.19 | 4.40 ± 0.18 | 4.41 ± 0.12 |
| 18:0 | 1.55 ± 0.13 | 1.52 ± 0.20 | 1.46 ± 0.10 | 0.51 ± 0.01 | 0.44 ± 0.11 | 0.50 ± 0.20 | 0.35 ± 0.14 |
| 18:1c9 | 28.15 ± 0.87 | 27.60 ± 0.64 | 28.97 ± 0.82 | 10.01 ± 0.69 | 15.28 ± 2.89 | 16.51 ± 3.02 | 14.27 ± 0.38 |
| 18:1c7 | 1.13 ± 0.23 | 0.68 ± 0.29 | 1.34 ± 0.20 | 0.59 ± 0.06 | 1.03 ± 0.21 | 1.12 ± 0.29 | 0.76 ± 0.04 |
| 18:2 | 11.76 ± 0.95 | 10.68 ± 0.73 | 11.58 ± 0.42 | 15.30 ± 0.96 | 15.10 ± 1.06 | 15.19 ± 0.80 | 13.26 ± 0.41 |
| 20:0 | 0.58 ± 0.14 | 0.67 ± 0.19 | 0.60 ± 0.09 | 0.11 ± 0.07 | 0.02 ± 0.01 | 0.13 ± 0.06 | ND |
| 20:1ω15 | 10.04 ± 0.54 | 10.21 ± 0.37 | 10.08 ± 0.22 | 3.85 ± 0.13 | 5.58 ± 2.00 | 6.71 ± 2.14 | 3.94 ± 0.02 |
| 18:3ω3 | 13.27 ± 0.84 | 9.35 ± 0.65 | 13.78 ± 0.66 | 11.73 ± 0.20 | 12.82 ± 0.73 | 12.68 ± 1.38 | 12.80 ± 0.37 |
| 20:2 | 0.15 ± 0.04 | 0.17 ± 0.04 | 0.12 ± 0.10 | 0.37 ± 0.08 | 0.28 ± 0.10 | 0.30 ± 0.19 | 0.09 ± 0.08 |
| 22:0 | 0.23 ± 0.16 | 0.46 ± 0.19 | 0.25 ± 0.09 | 0.28 ± 0.09 | 0.17 ± 0.10 | 0.12 ± 0.04 | 0.18 ± 0.10 |
| 22:1 | 25.96 ± 1.31 | 31.94 ± 2.20 | 24.69 ± 1.88 | 49.66 ± 1.91 | 44.22 ± 5.08 | 41.98 ± 4.42 | 49.75 ± 0.80 |
| 22:2 | ND | ND | ND | 1.03 ± 0.16 | 0.50 ± 0.36 | 0.37 ± 0.19 | 0.47 ± 0.25 |
| 24:1 | 0.84 ± 0.10 | 0.74 ± 0.13 | 0.79 ± 0.04 | 1.40 ± 0.45 | ND | ND | ND |
| Saturated | 8.70 ± 0.67 | 8.64 ± 0.87 | 8.67 ± 0.46 | 6.07 ± 0.05 | 5.18 ± 0.14 | 5.14 ± 0.21 | 4.82 ± 0.10 |
| MUFA | 66.12 ± 23.9 | 71.17 ± 2.21 | 65.86 ± 1.46 | 65.50 ± 0.97 | 66.12 ± 1.58 | 66.32 ± 2.05 | 69.25 ± 1.07 |
| PUFA | 25.17 ± 1.72 | 20.19 ± 1.37 | 25.48 ± 1.12 | 28.43 ± 0.92 | 28.69 ± 1.45 | 28.54 ± 2.00 | 25.93 ± 1.10 |
Table 4.
Concentration of erucic acid and total fatty acids in one-day-old radish, kale, and broccoli sprouts.
Table 4.
Concentration of erucic acid and total fatty acids in one-day-old radish, kale, and broccoli sprouts.
| | Concentration (mg/g DW) |
|---|
| Dark | Light |
|---|
| Sprout Variety | Erucic Acid (22:1) | Total Fatty Acids | Erucic Acid (22:1) | Total Fatty Acids |
|---|
| BSR | 16.50 ± 0.68 | 65.33 ± 1.95 | 19.28 ± 1.96 | 74.22 ± 5.82 |
| MIY | 18.89 ± 0.70 | 61.36 ± 2.29 | 21.59 ± 2.63 | 67.42 ± 3.56 |
| NT | 16.28 ± 4.70 | 65.75 ± 19.22 | 19.41 ± 3.23 | 78.28 ± 6.89 |
| JTK | 23.48 ± 1.05 | 49.55 ± 3.03 | 32.19 ± 7.06 | 63.61 ± 11.88 |
| JSB | 22.43 ± 4.05 | 48.47 ± 7.63 | 25.46 ± 4.64 | 57.29 ± 4.08 |
| GYP | 17.83 ± 1.51 | 45.59 ± 4.88 | 24.10 ± 1.95 | 58.13 ± 10.61 |
| MUM | 23.06 ± 3.68 | 45.76 ± 5.88 | 29.55 ± 8.34 | 57.92 ± 14.73 |
Table 5.
Glucosinolate composition of three-, five-, and seven-day-old radish, kale, and broccoli sprouts.
Table 5.
Glucosinolate composition of three-, five-, and seven-day-old radish, kale, and broccoli sprouts.
| | | GIB | PRO | GRA | GRE | 4-OHGBR | GIV | GER | GRPH | GBR | | |
|---|
| Sprout Variety | Age of Sprouts | 4.21 | 4.42 | 5.62 | 5.92 | 12.15 | 13.70 | 14.48 | 14.73 | 15.69 | Tot. GSL | Total Indole GSL |
|---|
| BSR | 3-day-old | ND | 2.54 ± 1.01 a | ND | 32.78 ± 3.79 a | 3.72 ± 0.43 a | 3.95 ± 0.12 | ND | 23.35 ± 0.00 a | ND | 67.01 ± 5.34 abA | 3.72 ± 0.43 a |
| 5-day-old | ND | 6.40 ± 0.39 bA | ND | 22.04 ± 0.97 b | 2.72 ± 0.18 abA | 6.49 ± 0.53 | ND | 50.30 ± 1.06 b | 0.34 ± 0.00 | 88.96 ± 3.12 aA | 3.05 ± 0.19 ab |
| 7-day-old | ND | 4.31 ± 0.94 ab | ND | 8.34 ± 1.55 c | 1.23 ± 0.14 b | 5.24 ± 0.40 | ND | 13.06 ± 1.49 c | 0.29 ± 0.04 | 33.13 ± 4.59 bA | 1.51 ± 0.18 b |
| JTK | 3-day-old | 2.87 ± 1.41 aA | 2.98 ± 1.18 | 78.83 ± 17.11 aA | ND | 4.11 ± 0.96 a | 4.04 ± 0.89 a | 15.83 ± 3.70 aA | ND | 1.20 ± 0.24 aA | 109.85 ± 24.47 aB | 5.30 ± 1.20 a |
| 5-day-old | 0.60 ± 0.85 bA | 3.16 ± 0.02 B | 18.07 ± 6.49 b | ND | 1.18 ± 0.39 bB | 5.63 ± 1.92 ab | 3.88 ± 0.99 b | ND | 0.41 ± 0.09 b | 32.92 ± 10.71 bB | 1.58 ± 0.48 b |
| 7-day-old | 2.73 ± 0.14 abA | 1.05 ± 0.06 | 77.09 ± 3.19 a | ND | 2.18 ± 0.02 bc | 8.01 ± 0.46 b | 13.35 ± 0.83 aA | ND | 1.07 ± 0.05 a | 105.47 ± 4.71 acB | 3.25 ± 0.03 ab |
| JSB | 3-day-old | 9.58 ± 0.40 aB | 4.64 ± 0.01 a | 32.44 ± 1.46 B | ND | 3.76 ± 0.08 a | 3.79 ± 0.09 a | 9.32 ± 0.36 B | ND | 0.79 ± 0.05 B | 64.32 ± 2.29 AC | 4.55 ± 0.12 a |
| 5-day-old | 5.81 ± 0.38 bB | 9.20 ± 0.28 bC | 18.67 ± 2.48 | ND | 1.88 ± 0.04 b | 9.10 ± 1.34 a | 7.94 ± 0.14 | ND | 0.61 ± 0.00 | 53.21 ± 1.43 AB | 2.49 ± 0.04 bc |
| 7-day-old | 4.72 ± 0.1 bB | 2.51 ± 0.04 ac | 13.15 ± 2.38 | ND | 1.45 ± 0.14 bc | 7.33 ± 1.22 b | 5.67 ± 0.57 B | ND | 0.57 ± 0.05 | 35.39 ± 4.41 A | 2.02 ± 0.18 c |
Table 6.
Fatty acid composition of Three, Five, and Seven-day radish, kale, and broccoli sprouts a.
Table 6.
Fatty acid composition of Three, Five, and Seven-day radish, kale, and broccoli sprouts a.
| | BSR | JTK | JSB |
|---|
| Fatty Acid | Day 3 | Day 5 | Day 7 | Day 3 | Day 5 | Day 7 | Day 3 | Day 5 | Day 7 |
|---|
| 16:00 | 7.49 ± 0.13 | 7.02 ± 0.30 | 7.40 ± 0.18 | 5.70 ± 0.57 | 5.36 ± 0.21 | 6.38 ± 0.36 | 5.09 ± 0.22 | 5.67 ± 0.34 | 5.68 ± 0.18 |
| 18:0 | 1.25 ± 0.05 | 1.27 ± 0.05 | 1.02 ± 0.06 | 0.26 ± 0.12 | ND | ND | ND | ND | ND |
| 18:1c9 | 23.43 ± 0.44 | 24.69 ± 0.68 | 24.55 ± 0.5 | 9.51 ± 0.24 | 9.82 ± 0.13 | 9.79 ± 0.35 | 12.64 ± 0.08 | 12.06 ± 0.76 | 11.37 ± 0.57 |
| 18:1c7 | 0.89 ± 0.06 | 0.83 ± 0.04 | 0.58 ± 0.05 | 0.47 ± 0.04 | 0.45 ± 0.05 | 0.65 ± 0.16 | 0.62 ± 0.18 | 1.06 ± 0.26 | 1.10 ± 0.42 |
| 18:2 | 12.74 ± 0.28 | 13.49 ± 0.36 | 14.02 ± 0.26 | 16.96 ± 0.11 | 17.14 ± 0.47 | 17.57 ± 0.54 | 17.41 ± 0.45 | 18.50 ± 0.59 | 17.53 ± 0.39 |
| 20:0 | 0.27 ± 0.04 | 0.34 ± 0.07 | ND | ND | ND | ND | ND | ND | ND |
| 20:1ω15 | 8.69 ± 0.17 | 8.45 ± 0.27 | 8.05 ± 0.23 | 3.28 ± 0.46 | 3.17 ± 0.49 | 2.73 ± 0.20 | 3.61 ± 0.12 | 2.76 ± 0.23 | 2.87 ± 0.25 |
| 18:3ω3 | 18.91 ± 0.48 | 18.62 ± 1.29 | 19.57 ± 0.55 | 15.34 ± 0.96 | 15.28 ± 0.91 | 16.42 ± 0.33 | 17.84 ± 0.42 | 21.32 ± 0.83 | 19.97 ± 0.93 |
| 20:2 | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| 22:0 | 0.07 ± 0.03 | 0.17 ± 0.1 | ND | ND | ND | ND | ND | ND | ND |
| 22:1 | 26.26 ± 0.99 | 25.11 ± 0.79 | 24.67 ± 0.44 | 47.77 ± 1.14 | 48.43 ± 0.72 | 46.19 ± 0.26 | 42.79 ± 0.73 | 38.62 ± 1.09 | 41.49 ± 1.14 |
| 22:02 | ND | ND | ND | 0.69 ± 0.18 | 0.35 ± 0.45 | 0.27 ± 0.20 | ND | ND | ND |
| Saturated | 9.09 ± 0.11 | 8.80 ± 0.17 | 8.42 ± 0.14 | 5.97 ± 0.45 | 5.36 ± 0.21 | 6.38 ± 0.36 | 5.09 ± 0.22 | 5.67 ± 0.34 | 5.68 ± 0.18 |
| MUFA | 67.67 ± 14.89 | 59.09 ± 1.71 | 57.86 ± 1.11 | 61.04 ± 1.34 | 61.87 ± 1.14 | 59.36 ± 0.35 | 59.66 ± 0.87 | 54.51 ± 1.7 | 56.82 ± 1.49 |
| PUFA | 31.65 ± 0.74 | 32.11 ± 1.64 | 33.59 ± 0.77 | 32.99 ± 0.89 | 32.76 ± 0.94 | 34.26 ± 0.06 | 35.25 ± 0.85 | 39.82 ± 1.4 | 37.50 ± 1.32 |
Table 7.
Concentration of erucic acid and total fatty acids in three-, five-, and seven-day radish, kale, and broccoli sprouts.
Table 7.
Concentration of erucic acid and total fatty acids in three-, five-, and seven-day radish, kale, and broccoli sprouts.
| | | Concentration (mg/g DW) |
|---|
| Sprout Variety | Age of Sprouts | Erucic Acid (22:1) | Total Fatty Acid |
|---|
| BSR | 3-day-old | 10.07 ± 0.15 | 38.38 ± 1.72 |
| 5-day-old | 10.44 ± 1.30 | 41.50 ± 3.89 |
| 7-day-old | 8.08 ± 0.41 | 32.75 ± 1.18 |
| JTK | 3-day-old | 18.35 ± 3.98 | 38.36 ± 7.59 |
| 5-day-old | 16.48 ± 5.89 | 32.34 ± 13.33 |
| 7-day-old | 12.23 ± 1.50 | 25.00 ± 4.64 |
| JSB | 3-day-old | 13.89 ± 1.68 | 32.43 ± 3.43 |
| 5-day-old | 9.03 ± 1.01 | 22.64 ± 3.43 |
| 7-day-old | 10.32 ± 1.32 | 23.32 ± 3.81 |
Table 8.
Pearson’s correlation coefficients among selected analyzed parameters in JSB sprouts.
Table 8.
Pearson’s correlation coefficients among selected analyzed parameters in JSB sprouts.
| | Tot GSL | GRE/GRA | Indole GSL | SFE/SFA | Phenolics | AOC a | Erucic Acid | Tot FA |
|---|
| Tot GSL | 1.00 | | | | | | | |
| GRE/GRA | 0.88 | 1.00 | | | | | | |
| Indole GSL | 0.77 | 0.72 | 1.00 | | | | | |
| SFE/SFA | 0.35 | 0.14 | 0.35 | 1.00 | | | | |
| Phenolics | −0.06 | −0.32 | 0.26 | 0.46 | 1.00 | | | |
| AOC a | −0.21 | −0.47 | 0.13 | 0.33 | 0.86 | 1.00 | | |
| Erucic acid | 0.40 | 0.58 | 0.53 | 0.03 | −0.49 | −0.30 | 1.00 | |
| Tot FA | 0.34 | 0.11 | 0.42 | 0.84 | 0.35 | 0.36 | 0.31 | 1.00 |



 and
and 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}