

Improved Synthesis of Deoxyadenosine Triphosphate by Saccharomyces cerevisiae Using an Efficient ATP Regeneration System: Optimization of Response Surface Analysis

Abstract
:1. Introduction
2. Results and Discussion
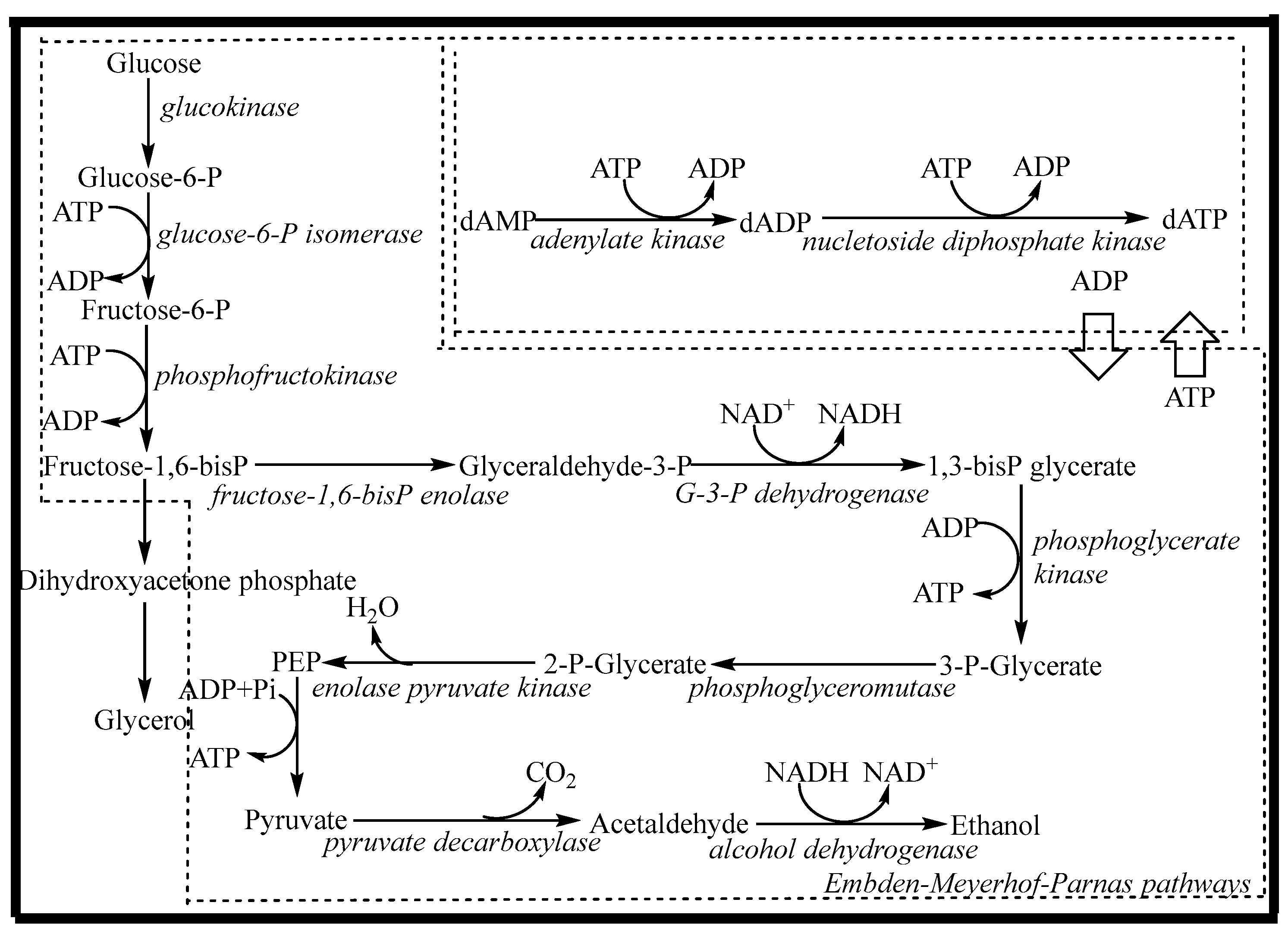
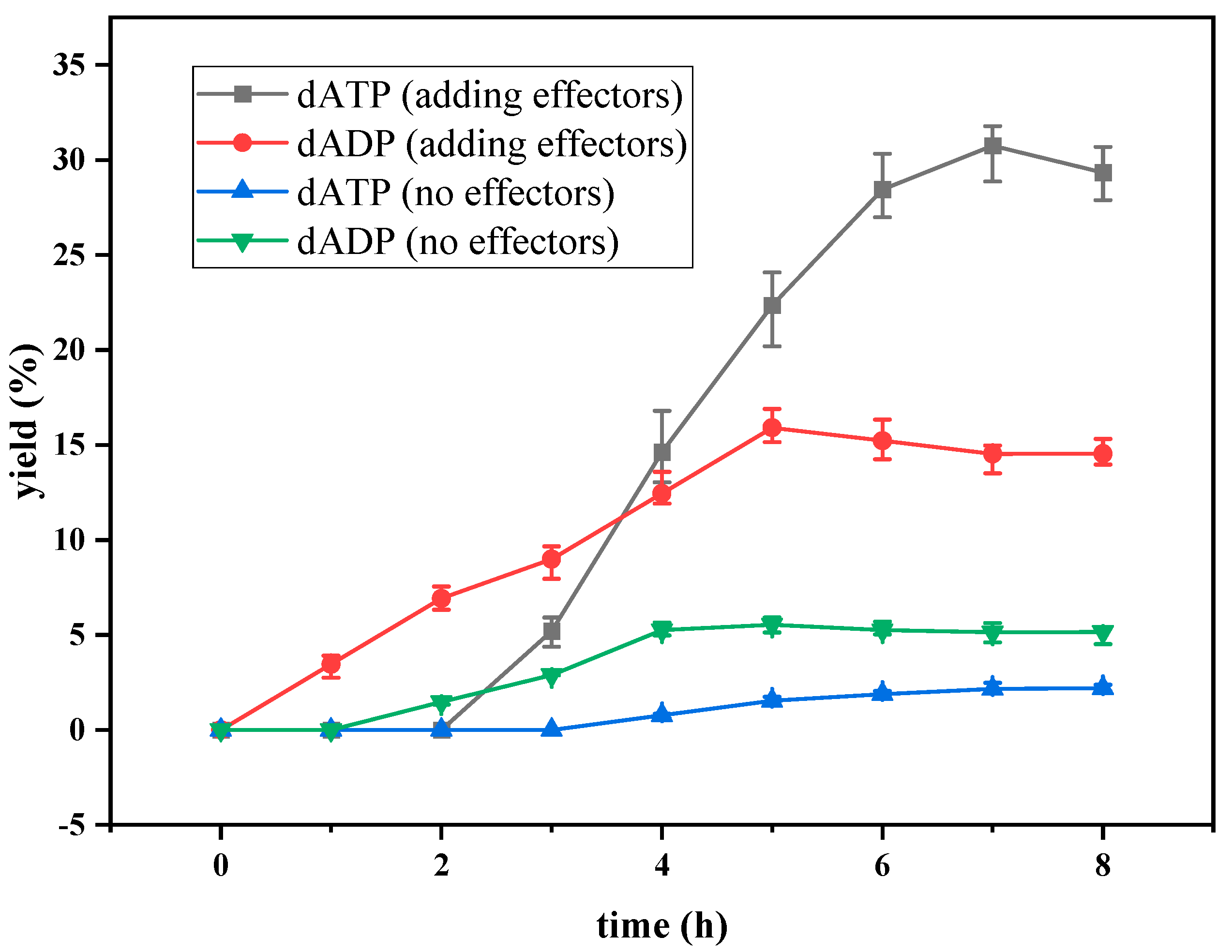
2.1. Preliminary Synthesis of dATP
2.2. Fractional Factorial Design to Identify Key Influences
2.3. Central Composite Design and Response Surface Methodology
2.4. Effect of Glucose on dATP Synthesis
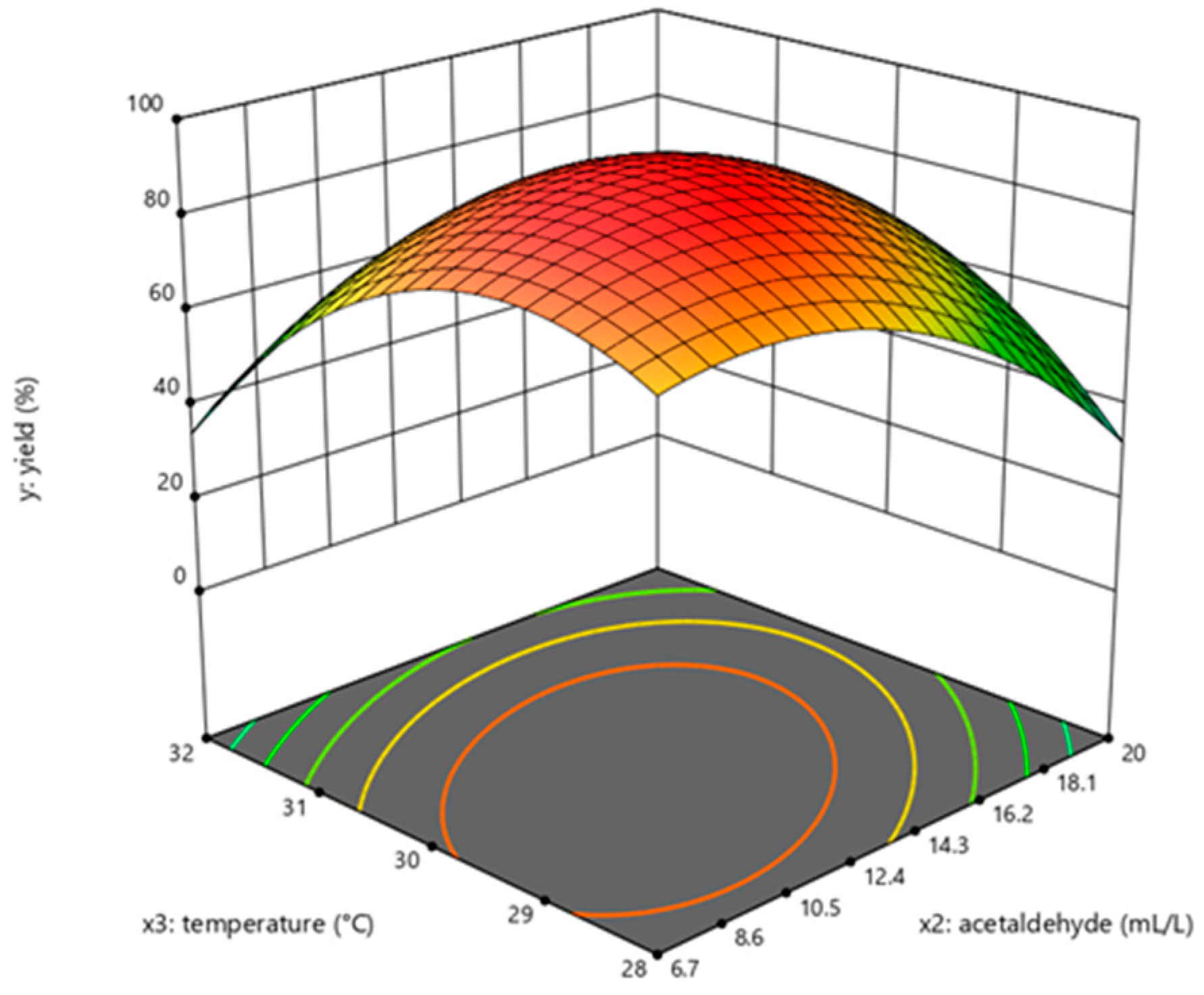
2.5. Effect of Acetaldehyde on dATP Synthesis
2.6. Effect of Temperature on dATP Synthesis
3. Materials and Methods
3.1. Strain and Medium
3.2. Preparation of Cells
3.3. dATP Synthesis Process
3.4. Analytic Method
3.5. Fractional Factorial Design (FFD)
3.6. Response Surface Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Schreier, V.N.; Loehr, M.O.; Deng, T.; Lattmann, E.; Hajnal, A.; Neuhauss, S.C.F.; Luedtke, N.W. Fluorescent dATP for DNA synthesis in vivo. ACS Chem. Biol. 2020, 15, 2996–3003. [Google Scholar] [CrossRef] [PubMed]
- Czernecki, D.; Bonhomme, F.; Kaminski, P.A.; Delarue, M. Characterization of a triad of genes in cyanophage S-2L sufficient to replace adenine by 2-aminoadenine in bacterial DNA. Nat. Commun. 2021, 12, 4710. [Google Scholar] [CrossRef]
- Zhu, J.; Chen, L.T. Highly efficient incorporation of dATP in terminal transferase polymerization forming the ploy (A)(n)-DITO-1 fluorescent probe sensing terminal transferase and T4 polynucleotide kinase activity. Anal. Chim. Acta 2022, 1221, 340080. [Google Scholar] [CrossRef]
- Cerqueira, N.M.F.S.A.; Fernandes, P.A.; Ramos, M.J. Ribonucleotide reductase: A critical enzyme for cancer chemotherapy and antiviral agents. Recent Pat. Anticancer Drug Discov. 2007, 2, 11–29. [Google Scholar] [CrossRef] [PubMed]
- Kapustina, Z.; Jaspone, A.; Dubovskaja, V.; Mackevicius, G.; Lubys, A. Enzymatic synthesis of chimeric DNA oligonucleotides by in vitro transcription with dTTP, dCTP, dATP, and 2 ‘-Fluoro modified dGTP. ACS Synth. Biol. 2021, 10, 1625–1632. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.; Khorana, H. Nucleoside polyphosphates. VI. An improved and general method for the synthesis of ribo- and deoxyribonucleoside 5′-triphosphates. J. Am. Chem. Soc. 1958, 80, 1141–1145. [Google Scholar] [CrossRef]
- Caton-Williams, J.; Smith, M.; Carrasco, N.; Huang, Z. Protection-free one-pot synthesis of 2’-deoxynucleoside 5 ‘-triphosphates and DNA polymerization. Org. Lett. 2011, 13, 4156–4159. [Google Scholar] [CrossRef]
- Fernandez-Lucas, J. Multienzymatic synthesis of nucleic acid derivatives: A general perspective. Appl. Microbiol. Biotechnol. 2015, 99, 4615–4627. [Google Scholar] [CrossRef]
- Bao, J.; Ryu, D.D.Y. Total biosynthesis of deoxynucleoside triphosphates using deoxynucleoside monophosphate kinases for PCR application. Biotechnol. Bioeng. 2007, 98, 1–11. [Google Scholar] [CrossRef]
- Robins, M.J. Ribonucleotide reductases: Radical chemistry and inhibition at the active site. Nucleosides Nucleotides Nucleic Acids 2003, 22, 519–534. [Google Scholar] [CrossRef]
- Suresh, P.; Vasu, N. Enzymatic synthesis of 2’-deoxynucleoside diphosphates and triphosphates. Biotechnol. Lett. 1998, 20, 1149–1151. [Google Scholar]
- Bochkov, D.V.; Khomov, V.V.; Tolstikova, T.G. Hydrolytic approach for production of deoxyribonucleoside-and ribonucleoside-5’-monophosphates and enzymatic synthesis of their polyphosphates. Biochemistry 2006, 71, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Yano, T.; Yasohara, Y.; Kashihara, M.; Tachiki, T.; Hidehiko, K.; Tatsurokuro, T. Production of deoxyribonucleoside triphosphates through coupled fermentation with energy transfer. Agric. Biol. Chem. 1989, 53, 1935–1940. [Google Scholar]
- Wu, W.; Bergstrom, D.E.; Davisson, V.J. A combination chemical and enzymatic approach for the preparation of azole carboxamide nucleoside triphosphate. J. Org. Chem. 2003, 68, 3860–3865. [Google Scholar] [CrossRef] [PubMed]
- Jong, A.Y.; Campbell, J.L. Characterization of Saccharomyces cerevisiae thymidylate kinase, the CDC8 gene product. General properties, kinetic analysis, and subcellular localization. J. Biol. Chem. 1984, 259, 14394–14398. [Google Scholar] [CrossRef]
- Bao, J.; Bruque, G.A.; Ryu, D.D.Y. Biosynthesis of deoxynucleoside triphosphates, dCTP and dTTP: Reaction mechanism and kinetics. Enzyme Microb. Technol. 2005, 36, 350–356. [Google Scholar] [CrossRef]
- Bao, J.; Ryu, D.D.Y. Biosynthesis reaction mechanism and kinetics of deoxynucleoside triphosphates, dATP and dGTP. Biotechnol. Bioeng. 2005, 89, 485–491. [Google Scholar] [CrossRef]
- Ding, Y.; Ou, L.; Ding, Q.B. Enzymatic Synthesis of Nucleoside Triphosphates and Deoxynucleoside Triphosphates by Surface-Displayed Kinases. Appl. Biochem. Biotechnol. 2019, 190, 1271–1288. [Google Scholar] [CrossRef]
- Alberty, R.A. Thermodynamic properties of nucleotide reductase reactions. Biochemistry 2004, 43, 9840–9845. [Google Scholar] [CrossRef]
- Antoniewicz, M.R. A guide to metabolic flux analysis in metabolic engineering: Methods, tools and applications. Metab. Eng. 2021, 63, 2–12. [Google Scholar] [CrossRef]
- Ni, C.; Dinh, C.V.; Prather, K.L.J. Dynamic Control of Metabolism. Annu. Rev. Chem. Biomol. Eng. 2021, 12, 519–541. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhang, S.; Jiang, P.X.; Yao, J.; He, Y.Z.; Chen, L.C.; Gui, X.W.; Dong, Z.Y.; Tang, S.Y. Design of an ectoine-responsive AraC mutant and its application in metabolic engineering of ectoine biosynthesis. Metab. Eng. 2015, 30, 149–155. [Google Scholar] [CrossRef]
- Fujio, T.; Furuya, A. Effects of magnesium ion and chelating agents on enzymatic production of ATP from adenine. Appl. Microbiol. Biotechnol. 1985, 21, 143–147. [Google Scholar] [CrossRef]
- Aust, A.; Yun, S.L.; Suelter, C.H. Pyruvate kinase from yeast (Saccharomyces cerevisiae). Methods Enzymol. 1975, 42, 176–182. [Google Scholar] [PubMed]
- Yoshino, M.; Murakami, K. AMP deaminase reaction as a control system of glycolysis in yeast. Role of ammonium ion in the interaction of phosphofructokinase and pyruvate kinase activity with the adenylate energy charge. J. Biol. Chem. 1985, 260, 4729–4732. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, Y.; Du, G.C.; Chen, J. Redirection of the NADH oxidation pathway in Torulopsis glabrata leads to an enhanced pyruvate production. Appl. Microbiol. Biotechnol. 2006, 72, 377–385. [Google Scholar] [CrossRef]
- Miguez, J.P.; Gama, R.S.; Bolina, I.C.A.; de Melo, C.C.; Cordeiro, M.R.; Hirata, D.B.; Mendes, A.A. Enzymatic synthesis optimization of a cosmetic ester catalyzed by a homemade biocatalyst prepared via physical adsorption of lipase on amino-functionalized rice husk silica. Chem. Eng. Res. Des. 2018, 139, 296–308. [Google Scholar] [CrossRef]
- Mendoza-Ortiz, P.A.; Gama, R.S.; Gomez, O.C.; Luiz, J.H.H.; Fernandez-Lafuente, R.; Cren, E.C.; Mendes, A.A. Sustainable Enzymatic Synthesis of a Solketal Ester-Process Optimization and Evaluation of Its Antimicrobial Activity. Catalysts 2020, 10, 218. [Google Scholar] [CrossRef]
- Tang, J.P.; Chen, Y.; Chen, X.C.; Yao, Y.L.; Ying, H.J.; Xiong, J.; Bai, J.X. Production of cytidine 5′-diphosphorylcholine with high utilization of ATP by whole cells of Saccharomyces cerevisiae. Bioresour. Technol. 2010, 101, 8807–8813. [Google Scholar] [CrossRef]
- Ernandes, J.R.; Meirsman, C.D.; Rolland, F.; Winderickx, J.; Winde, J.D.; Brandão, R.L.; Thevelein, J.M. During the initiation of fermentation overexpression of hexokinase PII in yeast transiently causes a similar deregulation of glycolysis as deletion of Tps1. Yeast 1998, 14, 255–269. [Google Scholar] [CrossRef]
- Akria, K.; Makoto, M.; Kosaku, M. Phosphorylation of CMP by the released CMP kinase system from a Yeast: Saccharomyces carlsbergensis. Agric. Biol. Chem. 1975, 39, 621–625. [Google Scholar]
- Li, Y.; Hugenholtz, J.; Chen, J.; Lun, S.Y. Enhancement of pyruvate production by Torulopsis glabrata using a two-stage oxygen supply control strategy. Appl. Microbiol. Biotechnol. 2002, 60, 101–106. [Google Scholar] [PubMed]
- Barber, A.R.; Vriesekoop, F.; Pamment, N.B. Effects of acetaldehyde on Saccharomyces cerevisiae exposed to a range of chemical and environmental stresses. Enzyme Microb. Technol. 2002, 30, 240–250. [Google Scholar] [CrossRef]
- Stanley, G.A.; Hobley, T.J.; Pamment, N.B. Effect of acetaldehyde on Saccharomyces cerevisiae and Zymomonas mobilis subjected to environmental shocks. Biotechnol. Bioeng. 1997, 53, 71–78. [Google Scholar] [CrossRef]
- Vriesekoop, F.; Barber, A.R.; Pamment, N.B. Acetaldehyde mediates growth stimulation of ethanol-stressed Saccharomyces cerevisiae: Evidence of a redox-driven mechanism. Biotechnol. Lett. 2007, 29, 1099–1103. [Google Scholar] [CrossRef]
- Tuma, D.J.; Casey, C.A. Dangerous byproducts of alcohol breakdown-Focus on adducts. Alcohol. Res. Health 2003, 27, 285–290. [Google Scholar]
- Choi, D.; Cho, K.; Jung, J. Optimization of nitrogen removal performance in a single-stage SBR based on partial nitritation and ANAMMOX. Water Res. 2019, 162, 105–114. [Google Scholar] [CrossRef]
- Li, D.; Chen, H.; Gao, X.; Zhang, J. Establishment and optimization of partial nitriffcation/anammox/partial nitriffcation/anammox (PN/A/PN/A) process based on multi-stage ammonia oxidation: Using response surface method as a tool. Bioresour. Technol. 2022, 361, 127722. [Google Scholar] [CrossRef]
- Fatah, N.A.A.; Triwahyono, S.; Jalil, A.A.; Salamun, N.; Mamat, C.R.; Majid, Z.A. n-Heptane isomerization over molybdenum supported on bicontinuous concentric lamellar silica KCC-1: Inffuence of phosphorus and optimization using response surface methodology (RSM). Chem. Eng. J. 2017, 314, 650–659. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coding Level | x1 | x2 | x3 | x4 | x5 | x6 | x7 | x8 | x9 | x10 |
|---|---|---|---|---|---|---|---|---|---|---|
| dAMP (g/L) | Glucose (g/L) | MgCl2·6H2O (g/L) | KCl (g/L) | NaH2PO4 (g/L) | Yeast (g/L) | NH4Cl (g/L) | Acetaldehyde (mL/L) | pH | T (°C) | |
| −1 | 0.80 | 30.00 | 1.00 | 1.33 | 26.00 | 250.00 | 0.33 | 6.70 | 6.5 | 28.0 |
| +1 | 2.00 | 50.00 | 7.00 | 2.67 | 36.40 | 350.00 | 1.00 | 20.00 | 7.5 | 32.0 |
| Run | Factors | Y (%) | Y′ (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| x1 | x2 | x3 | x4 | x5 | x6 | x7 | x8 | x9 | x10 | |||
| 1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | 1 | 1 | 41.68 | 41.25 |
| 2 | 1 | −1 | −1 | −1 | 1 | −1 | 1 | 1 | −1 | −1 | 9.87 | 7.54 |
| 3 | −1 | 1 | −1 | −1 | 1 | 1 | −1 | 1 | −1 | −1 | 33.71 | 32.48 |
| 4 | 1 | 1 | −1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | 70.64 | 67.50 |
| 5 | −1 | −1 | 1 | −1 | 1 | 1 | 1 | −1 | −1 | 1 | 44.93 | 57.27 |
| 6 | 1 | −1 | 1 | −1 | −1 | 1 | −1 | 1 | 1 | −1 | 5.90 | 0.98 |
| 7 | −1 | 1 | 1 | −1 | −1 | −1 | 1 | 1 | 1 | −1 | 15.22 | 23.71 |
| 8 | 1 | 1 | 1 | −1 | 1 | −1 | −1 | −1 | −1 | 1 | 79.03 | 70.25 |
| 9 | −1 | −1 | −1 | 1 | −1 | 1 | 1 | 1 | −1 | 1 | 23.53 | 14.75 |
| 10 | 1 | −1 | −1 | 1 | 1 | 1 | −1 | −1 | 1 | −1 | 27.40 | 35.89 |
| 11 | −1 | 1 | −1 | 1 | 1 | −1 | 1 | −1 | 1 | −1 | 63.54 | 58.62 |
| 12 | 1 | 1 | −1 | 1 | −1 | −1 | −1 | 1 | −1 | 1 | 15.83 | 27.72 |
| 13 | −1 | −1 | 1 | 1 | 1 | −1 | −1 | 1 | 1 | 1 | 26.59 | 23.45 |
| 14 | 1 | −1 | 1 | 1 | −1 | −1 | 1 | −1 | −1 | −1 | 22.39 | 21.16 |
| 15 | −1 | 1 | 1 | 1 | −1 | 1 | −1 | −1 | −1 | −1 | 48.42 | 46.09 |
| 16 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 50.13 | 49.70 |
| Term | Regression Analysis | ||
|---|---|---|---|
| Coefficient | F-Value | p-Value | |
| x1 | −1.05 | 0.1299 | 0.7332 |
| x2 | 10.86 | 13.7700 | 0.0138 |
| x3 | 0.43 | 0.0215 | 0.8893 |
| x4 | −1.48 | 0.2539 | 0.6357 |
| x5 | 5.75 | 3.8600 | 0.1066 |
| x6 | 1.93 | 0.4370 | 0.5378 |
| x7 | 1.38 | 0.2235 | 0.6563 |
| x8 | −13.61 | 21.6100 | 0.0056 |
| x9 | 1.49 | 0.2591 | 0.6324 |
| x10 | 7.84 | 7.1800 | 0.0439 |
| Factors | Levels of Factors | ||||
|---|---|---|---|---|---|
| −1.618 | −1 | 0 | 1 | 1.618 | |
| x1 (glucose, g/L) | 23.2 | 30 | 40 | 50 | 56.8 |
| x2 (acetaldehyde, mL/L) | 2.2 | 6.7 | 13.4 | 20 | 24.5 |
| x3 (temperature, °C) | 26.6 | 28 | 30 | 32 | 33.4 |
| Run | x1 | x2 | x3 | Y (%) | Y′ (%) |
|---|---|---|---|---|---|
| 1 | 1 | −1 | 1 | 23.24 | 24.09 |
| 2 | 0 | 0 | 0 | 90.18 | 91.19 |
| 3 | −1 | −1 | 1 | 19.55 | 29.18 |
| 4 | −1 | 1 | −1 | 11.33 | 21.00 |
| 5 | 0 | 0 | 0 | 89.64 | 91.19 |
| 6 | 0 | −1.618 | 0 | 54.65 | 51.38 |
| 7 | −1.618 | 0 | 0 | 86.46 | 68.19 |
| 8 | −1 | −1 | −1 | 56.03 | 62.52 |
| 9 | 0 | 0 | 1.618 | 24.73 | 13.17 |
| 10 | 0 | 1.618 | 0 | 42.74 | 31.13 |
| 11 | −1 | 1 | 1 | 36.19 | 49.65 |
| 12 | 1 | 1 | −1 | 27.52 | 28.40 |
| 13 | 0 | 0 | −1.618 | 33.47 | 30.16 |
| 14 | 1 | −1 | −1 | 75.90 | 72.95 |
| 15 | 1.618 | 0 | 0 | 66.73 | 70.13 |
| 16 | 1 | 1 | 1 | 37.50 | 41.53 |
| Term | Regression Analysis | ||
|---|---|---|---|
| Coefficient | F-Value | p-Value | |
| x1 | 0.97 | 0.0254 | 0.8787 |
| x2 | −6.02 | 2.7600 | 0.1476 |
| x3 | −5.05 | 1.9500 | 0.2125 |
| x12 | −22.03 | 0.0256 | 0.8781 |
| x22 | −17.65 | 0.6731 | 0.4433 |
| x32 | −24.58 | 10.7200 | 0.0169 |
| x1x2 | −1.27 | 3.1400 | 0.1270 |
| x1x3 | −6.53 | 16.1100 | 0.0070 |
| x2x3 | 15.50 | 31.2400 | 0.0014 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, J.; Xu, H.; Wang, Q.; Sun, W. Improved Synthesis of Deoxyadenosine Triphosphate by Saccharomyces cerevisiae Using an Efficient ATP Regeneration System: Optimization of Response Surface Analysis. Molecules 2023, 28, 4029. https://doi.org/10.3390/molecules28104029
Xiong J, Xu H, Wang Q, Sun W. Improved Synthesis of Deoxyadenosine Triphosphate by Saccharomyces cerevisiae Using an Efficient ATP Regeneration System: Optimization of Response Surface Analysis. Molecules. 2023; 28(10):4029. https://doi.org/10.3390/molecules28104029
Chicago/Turabian StyleXiong, Jian, Hanghang Xu, Qi Wang, and Wenyuan Sun. 2023. "Improved Synthesis of Deoxyadenosine Triphosphate by Saccharomyces cerevisiae Using an Efficient ATP Regeneration System: Optimization of Response Surface Analysis" Molecules 28, no. 10: 4029. https://doi.org/10.3390/molecules28104029