
Extract from Black Soybean Cultivar A63 Extract Ameliorates Atopic Dermatitis-like Skin Inflammation in an Oxazolone-Induced Murine Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Analysis of Isoflavones and Flavonoids in Korean Black Soybean Cultivars
2.2. A63 Extract Attenuates Oxazolone-Induced AD-like Skin Inflammation in Mice
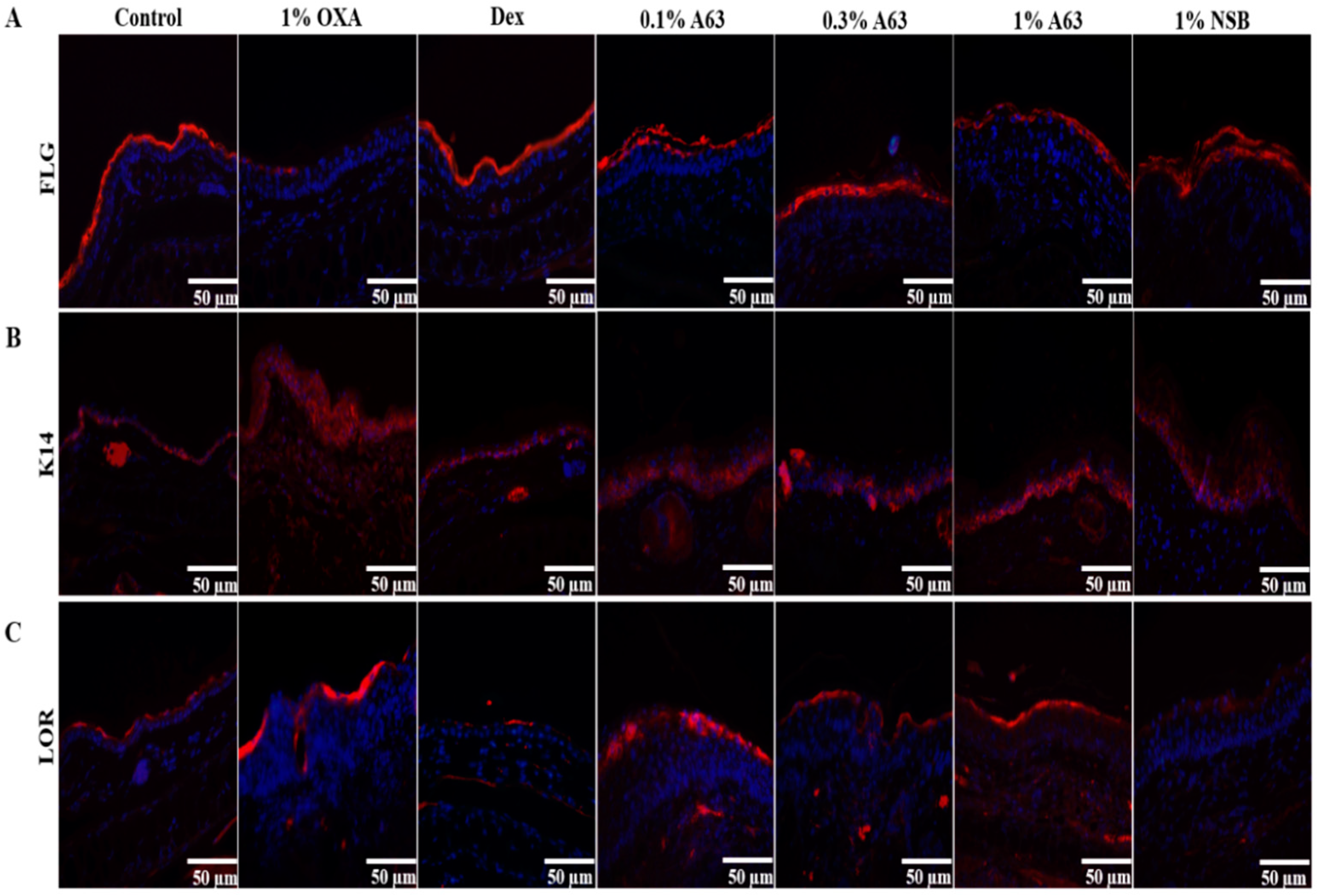
2.3. The Application of A63 Rescued the Expression of Keratinocyte Differentiation Markers
2.4. A63 Extract Suppresses the Expression of AD Hallmark Genes in Mouse Tissue
2.5. MAPK and STAT Signaling Activation was Hindered by Treatment with A63 Extract
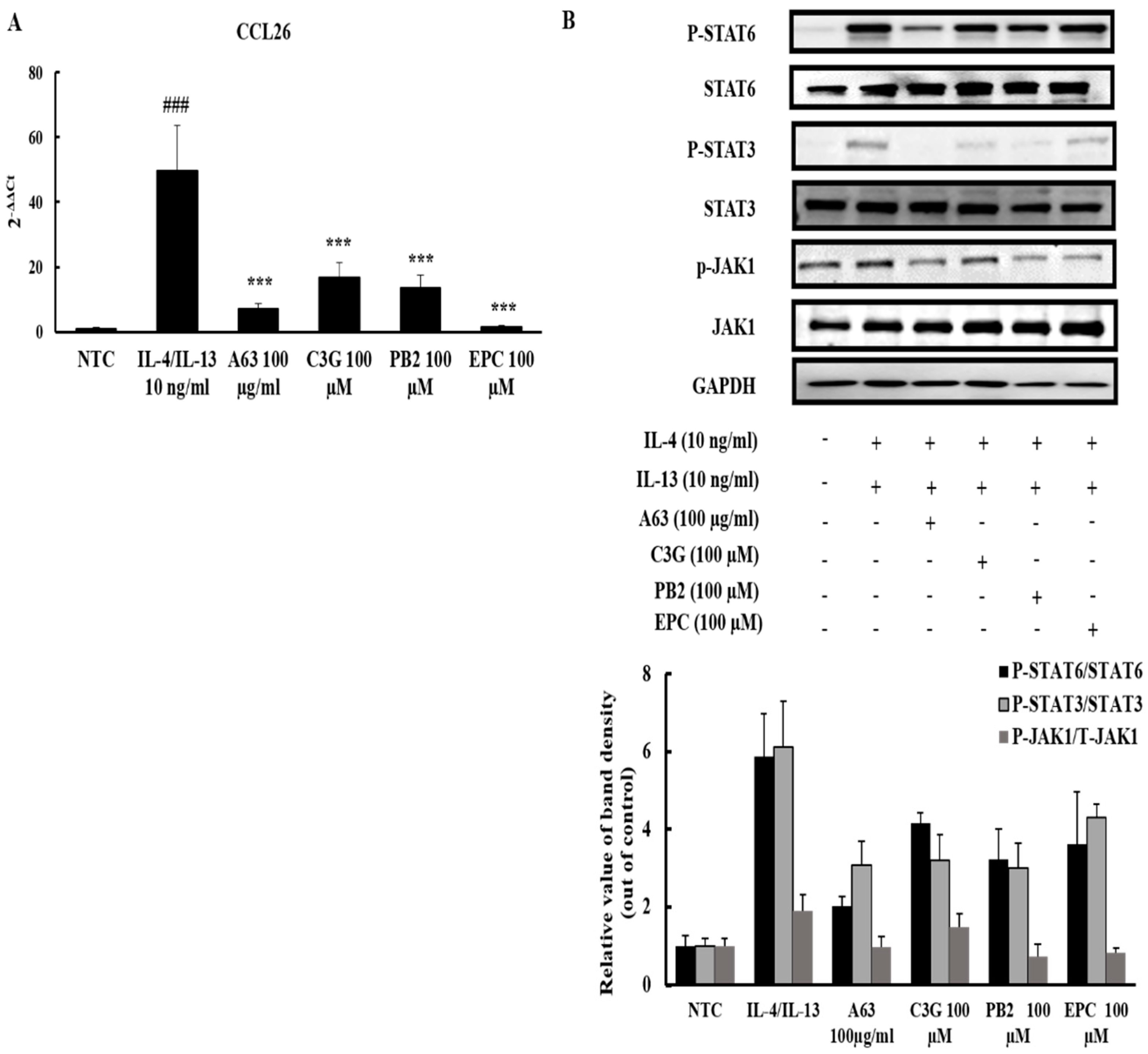
2.6. A63 Extract Prevents Eosinophil Infiltration toward the Inflammatory Site via CCL26 Expression Downregulation and the Inactivation of JAK1/STAT6 Signaling
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Analysis of Isoflavones and Other Flavonoids
4.3. Animal Study
4.4. Immunohistochemistry
4.5. RT-qPCR Analysis
4.6. Cell Culture
4.7. Western Blot Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Simpson, E.L. Atopic dermatitis: A review of topical treatment options. Curr. Med. Res. Opin. 2010, 26, 633. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; BuddenKotte, J.; Berger, T.G.; Steinhoff, M. Management of itch in atopic dermatitis. Semin. Cutan. Med. Surg. 2011, 30, 71–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, E.K.; Seo, E.-H.; Park, J.H.; Shim, H.R.; Kim, K.H.; Lee, B.R. Anti-inflammatory and anti-allergic effect of extracts from organic soybean. Korean J. Org. Agric. 2011, 19, 245–253. [Google Scholar]
- De Benedetto, A.; Rafaels, N.M.; McGirt, L.Y.; Ivanov, A.I.; Georas, S.N.; Cheadle, C.; Bergert, A.E.; Zhang, K.; Vidyasagar, S.; Yoshida, T.; et al. Tight junction defects in patients with atopic dermatitis. J. Allergy Clin. Immun. 2011, 127, 773–786. [Google Scholar] [CrossRef] [Green Version]
- Boguniewicz, M.; Leung, D.Y. Recent insights into atopic dermatitis and implications for management of infectious complications. J. Allergy Clin. Immun. 2010, 125, 4–13. [Google Scholar] [CrossRef] [Green Version]
- Hongxiu Ji, X.-K.L. Oxidative stress in Atopic dermatitis. Oxid. Med. Cell. Longev. 2016, 2016, 2721469. [Google Scholar] [CrossRef]
- Esra Birben, U.M.S. Cansin Sackesen, Serpil Erzurum, Omer Kalayci, Oxidative Stress and Antioxidant Defense. WAO J. 2012, 5, 10. [Google Scholar]
- Carretero, M.; Guerrero-Aspizua, S.; Illera, B.; Galver, V.; Navarro, M.; Garcia-Garcia, F.; Dopazo, J.; Jorcano, J.L.; Larcher, F.; del Rio, M. Differential Features between Chronic Skin Inflammatory Diseases Revealed in Skin-Humanized Psoriasis and Atopic Dermatitis Mouse Models. J. Investig. Dermatol. 2016, 136, 136–145. [Google Scholar] [CrossRef] [Green Version]
- Kammeyer, A.; Luiten, R.M. Oxidation events and skin aging. Ageing Res. Rev. 2015, 21, 16–29. [Google Scholar] [CrossRef]
- Okayama, Y. Oxidative stress in allergic and inflammatory skin diseases. Curr. Drug. Targets Inflamm. Allergy 2005, 4, 517–519. [Google Scholar] [CrossRef]
- Simonetti, O.; Bacchetti, T.; Ferretti, G.; Molinelli, E.; Rizzetto, G.; Bellachioma, L.; Offidani, A. Oxidative Stress and Alterations of Paraoxonases in Atopic Dermatitis. Antioxidants 2021, 10, 697. [Google Scholar] [CrossRef]
- Jang, S.; Ohn, J.; Kim, J.W.; Kang, S.M.; Jeon, D.; Heo, C.Y.; Lee, Y.-S.; Kwon, O.; Kim, K.H. Caffeoyl–Pro–His amide relieve. DNCB-Induced Atopic Dermatitis-Like phenotypes in BALB/c mice. Sci. Rep. 2020, 10, 8417. [Google Scholar] [CrossRef]
- Sivaranjani, N.; Rao, S.V.; Rajeev, G. Role of reactive oxygen species and antioxidants in atopic dermatitis. J. Clin. Diagn. Res. JCDR 2013, 7, 2683. [Google Scholar] [CrossRef]
- Guttman-Yassky, E.; Nograles, K.E.; Krueger, J.G. Contrasting pathogenesis of atopic dermatitis and psoriasis--part I: Clinical and pathologic concepts. J. Allergy Clin. Immun. 2011, 127, 1110–1118. [Google Scholar] [CrossRef]
- Rodenbeck, D.L.; Silverberg, J.I.; Silverberg, N.B. Phototherapy for atopic dermatitis. Clin. Dermatol. 2016, 34, 607–613. [Google Scholar] [CrossRef]
- Kumar Ganesan, B.X. A Critical review on polyphenols and health benefits of black soybeans. Nutrients 2017, 9, 455. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Wen, X.Y.; Zhang, X.Q.; Pu, H.M.; Kan, J.; Jin, C.H. Extraction, characterization and in vitro antioxidant activity of polysaccharides from black soybean. Int. J. Biol. Macromol. 2015, 72, 9. [Google Scholar] [CrossRef]
- Xu, J.L.; Shin, J.-S.; Park, S.-K.; Kang, S.; Jeong, S.-C.; Moon, J.-K.; Choi, Y.J.F.R.I. Differences in the metabolic profiles and antioxidant activities of wild and cultivated black soybeans evaluated by correlation analysis. Food Res. Int. 2017, 100, 166–174. [Google Scholar] [CrossRef]
- Hyeon, H.; Xu, J.L.; Kim, J.K.; Choi, Y.J.F.R.I. Comparative metabolic profiling of cultivated and wild black soybeans reveals distinct metabolic alterations associated with their domestication. Food Res. Int. 2020, 134, 109290. [Google Scholar] [CrossRef]
- Jung, T.; Stingl, G. Atopic dermatitis: Therapeutic concepts evolving from new pathophysiologic insights. J. Allergy Clin. Immun. 2008, 122, 1074–1081. [Google Scholar] [CrossRef]
- Weaver, C.T.; Elson, C.O.; Fouser, L.A.; Kolls, J.K. The Th17 pathway and inflammatory diseases of the intestines, lungs, and skin. Annu. Rev. Pathol.-Mech. 2013, 8, 477–512. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, Z.; Hassanshahi, G.; Khorramdelazad, H.; Zainodini, N.; Koochakzadeh, L. An Overlook to the Characteristics and Roles Played by Eotaxin Network in the Pathophysiology of Food Allergies: Allergic Asthma and Atopic Dermatitis. Inflammation 2016, 39, 1253–1267. [Google Scholar] [CrossRef]
- Spergel, J.M.; Paller, A.S. Atopic dermatitis and the atopic march. J. Allergy Clin. Immun. 2003, 112 (Suppl. S6), S118–S127. [Google Scholar] [CrossRef]
- Baek, J.; Lee, M.G. Oxidative stress and antioxidant strategies in dermatology. Redox Rep. 2016, 21, 164–169. [Google Scholar] [CrossRef]
- Choi, D.I.; Park, J.H.; Choi, J.Y.; Piao, M.; Suh, M.S.; Lee, J.B.; Yun, S.J.; Lee, S.C. Keratinocytes-Derived Reactive Oxygen Species Play an Active Role to Induce Type 2 Inflammation of the Skin: A Pathogenic Role of Reactive Oxygen Species at the Early Phase of Atopic Dermatitis. Ann. Dermatol. 2021, 33, 26–36. [Google Scholar] [CrossRef]
- Yan, C.; Ying, J.; Lu, W.; Changzhi, Y.; Qihong, Q.; Jingzhu, M.; Dongjie, S.; Tingting, Z. MiR-1294 suppresses ROS-dependent inflammatory response in atopic dermatitis via restraining STAT3/NF-kappaB pathway. Cell Immunol. 2022, 371, 104452. [Google Scholar] [CrossRef]
- Collantes, T.M.; Rho, M.C.; Kwon, H.J.; Jung, B.G.; Alfajaro, M.M.; Kim, D.S.; Kim, H.J.; Hosmillo, M.; Park, J.G.; Son, K.Y.; et al. Azuki bean (Vigna angularis) extract inhibits the development of experimentally induced atopic dermatitis-like skin lesions in NC/Nga mice. Food Chem. 2012, 132, 1269–1275. [Google Scholar] [CrossRef]
- Kasraie, S.; Werfel, T. Role of macrophages in the pathogenesis of atopic dermatitis. Mediat. Inflamm. 2013, 2013, 942375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.; Lee, C.H.; Kim, J.R.; Kwon, J.Y.; Son, M.J.; Kim, J.E.; Lee, K.W. Theobroma cacao extract attenuates the development of Dermatophagoides farinae-induced atopic dermatitis-like symptoms in NC/Nga mice. Food Chem. 2017, 216, 19–26. [Google Scholar] [CrossRef]
- Plotz, S.G.; Ring, J. What’s new in atopic eczema? Expert Opin. Emerg. Drugs 2010, 15, 249–267. [Google Scholar] [CrossRef]
- Kim, K.E.; Jung, M.J.; Houh, Y.; Kim, T.S.; Lee, W.J.; Yang, Y.; Bang, S.I.; Kim, C.H.; Kim, H.; Park, H.J.; et al. Erdr1 Attenuates Dermatophagoides farina Body Extract-Induced Atopic Dermatitis in NC/Nga Mice. J. Investig. Dermatol. 2017, 137, 1798–1802. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.-J.; Ha, T.-J.; Choi, H.-N.; Lee, J.-S.; Park, Y.-I. Inhibitory Effects of Anthocyanins Isolated from Black Soybean (Glycine max L.) Seed Coat on Degranulation and Cytokine Generation in RBL-2H3 Cells. J. Korean Soc. Food Sci. Nutr. 2011, 40, 1662–1667. [Google Scholar] [CrossRef]
- Yoon, Y.; Lee, Y.M.; Song, S.; Lee, Y.Y.; Yeum, K.J. Black soybeans protect human keratinocytes from oxidative stress-induced cell death. Food Sci. Nutr. 2018, 6, 2423–2430. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Wu, Y.; Chen, B.; Yao, Y.; Wang, Y.; Bai, H.; Li, C.; Yang, Y.; Chen, Y. Cyanidin-3-O-β-glucoside attenuates allergic airway inflammation by modulating the IL-4Rα-STAT6 signaling pathway in a murine asthma model. Int. Immunopharmacol. 2019, 69, 10. [Google Scholar] [CrossRef] [PubMed]
- Coleman, S.L.; Kruger, M.C.; Sawyer, G.M.; Hurst, R.D. Procyanidin A2 Modulates IL-4-Induced CCL26 Production in Human Alveolar Epithelial Cells. Int. J. Mol. Sci. 2016, 17, 1888. [Google Scholar] [CrossRef] [Green Version]
- Jung, A.R.; Ahn, S.H.; Park, I.S.; Park, S.Y.; Jeong, S.I.; Cheon, J.H.; Kim, K. Douchi (fermented Glycine max Merr.) alleviates atopic dermatitis-like skin lesions in NC/Nga mice by regulation of PKC and IL-4. BMC Complement. Altern. Med. 2016, 16, 416. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Ohgo, Y.; Katayanagi, Y.; Yasui, K.; Hiramoto, S.; Ikemoto, H.; Nakata, Y.; Miyoshi, N.; Isemura, M.; Ohashi, N.J.S.R. Anti-inflammatory effects of green soybean extract irradiated with visible light. Sci. Rep. 2014, 4, 4732. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dorjsembe, B.; Nho, C.W.; Choi, Y.; Kim, J.-C. Extract from Black Soybean Cultivar A63 Extract Ameliorates Atopic Dermatitis-like Skin Inflammation in an Oxazolone-Induced Murine Model. Molecules 2022, 27, 2751. https://doi.org/10.3390/molecules27092751
Dorjsembe B, Nho CW, Choi Y, Kim J-C. Extract from Black Soybean Cultivar A63 Extract Ameliorates Atopic Dermatitis-like Skin Inflammation in an Oxazolone-Induced Murine Model. Molecules. 2022; 27(9):2751. https://doi.org/10.3390/molecules27092751
Chicago/Turabian StyleDorjsembe, Banzragch, Chu Won Nho, Yongsoo Choi, and Jin-Chul Kim. 2022. "Extract from Black Soybean Cultivar A63 Extract Ameliorates Atopic Dermatitis-like Skin Inflammation in an Oxazolone-Induced Murine Model" Molecules 27, no. 9: 2751. https://doi.org/10.3390/molecules27092751