
Natural Compound Shikonin Is a Novel PAK1 Inhibitor and Enhances Efficacy of Chemotherapy against Pancreatic Cancer Cells

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Lines
2.3. Determination of Enzymatic Activity Inhibition
2.4. Molecular Docking
2.5. Assessment of Cell Proliferation and Apoptosis
2.6. Western Blot Analysis
2.7. Combination Index (CI)
2.8. Statistical Analysis
3. Results
3.1. Screening of PAK1 Inhibitors
3.2. Shikonin Inhibits PAK1 by Interacting with ATP Pocket
3.3. Shikonin Inhibited the Growth of Pancreatic Cancer Cells
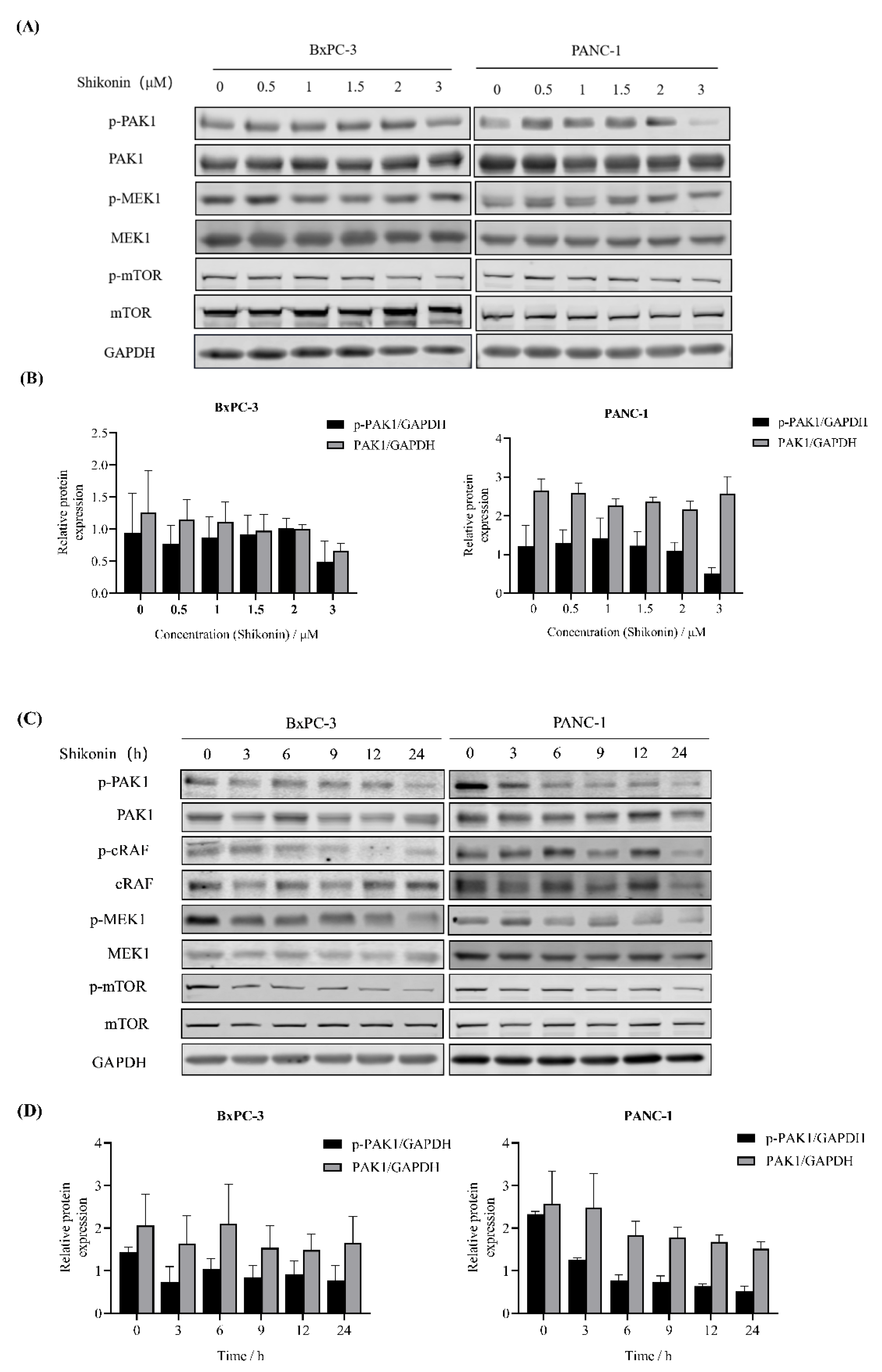
3.4. Shikonin Suppressed Downstream Pathways of PAK1 in Pancreatic Cancer Cells
3.5. Shikonin Induced Apoptosis of Pancreatic Cancer Cells
3.6. Shikonin Sensitized Pancreatic Cancer Cells to Chemotherapeutic Drugs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducreux, M.; Cuhna, A.S.; Caramella, C.; Hollebecque, A.; Burtin, P.; Goere, D.; Seufferlein, T.; Haustermans, K.; Van Laethem, J.L.; Conroy, T.; et al. Cancer of the pancreas: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2015, 26 (Suppl. 5), v56–v68. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J Clin 2021, 71, 7–33. [Google Scholar] [CrossRef]
- Motoi, F.; Kosuge, T.; Ueno, H.; Yamaue, H.; Satoi, S.; Sho, M.; Honda, G.; Matsumoto, I.; Wada, K.; Furuse, J.; et al. Randomized phase II/III trial of neoadjuvant chemotherapy with gemcitabine and S-1 versus upfront surgery for resectable pancreatic cancer (Prep-02/JSAP05). Jpn. J. Clin. Oncol. 2019, 49, 190–194. [Google Scholar] [CrossRef] [Green Version]
- Conroy, T.; Hammel, P.; Hebbar, M.; Ben Abdelghani, M.; Wei, A.C.; Raoul, J.L.; Chone, L.; Francois, E.; Artru, P.; Biagi, J.J.; et al. FOLFIRINOX or Gemcitabine as Adjuvant Therapy for Pancreatic Cancer. N. Engl. J. Med. 2018, 379, 2395–2406. [Google Scholar] [CrossRef]
- Zeng, S.; Pottler, M.; Lan, B.; Grutzmann, R.; Pilarsky, C.; Yang, H. Chemoresistance in Pancreatic Cancer. Int. J. Mol. Sci. 2019, 20, 4504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, H.; Nicholas, N.S.; Wells, C.M. Role of p-21-activated kinases in cancer progression. Int. Rev. Cell Mol. Biol. 2014, 309, 347–387. [Google Scholar]
- Ong, C.C.; Gierke, S.; Pitt, C.; Sagolla, M.; Cheng, C.K.; Zhou, W.; Jubb, A.M.; Strickland, L.; Schmidt, M.; Duron, S.G.; et al. Small molecule inhibition of group I p21-activated kinases in breast cancer induces apoptosis and potentiates the activity of microtubule stabilizing agents. Breast. Cancer Res. 2015, 17, 59. [Google Scholar] [CrossRef] [Green Version]
- Song, B.; Wang, W.; Zheng, Y.; Yang, J.; Xu, Z. P21-activated kinase 1 and 4 were associated with colorectal cancer metastasis and infiltration. J. Surg. Res. 2015, 196, 130–135. [Google Scholar] [CrossRef]
- Yeo, D.; Huynh, N.; Beutler, J.A.; Christophi, C.; Shulkes, A.; Baldwin, G.S.; Nikfarjam, M.; He, H. Glaucarubinone and gemcitabine synergistically reduce pancreatic cancer growth via down-regulation of P21-activated kinases. Cancer Lett. 2014, 346, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Liu, H.; Chen, L.; Wang, S.; Zhou, L.; Yun, X.; Sun, L.; Wen, Y.; Gu, J. Hepatitis B virus X protein confers resistance of hepatoma cells to anoikis by up-regulating and activating p21-activated kinase 1. Gastroenterology 2012, 143, 199–212.e4. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, Y.; Chen, J.; Yang, Y.; Zhu, L.; Zhao, J.; Yang, Y.; Cai, X.; Hu, C.; Rosell, R.; et al. Identification of a novel PAK1 inhibitor to treat pancreatic cancer. Acta. Pharm. Sin. B 2020, 10, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Symeonidis, N.; Lambropoulou, M.; Pavlidis, E.; Anagnostopoulos, C.; Tsaroucha, A.; Kotini, A.; Nikolaidou, C.; Kiziridou, A.; Simopoulos, C. PAK1 Expression in Pancreatic Cancer: Clinicopathological Characteristics and Prognostic Significance. Clin. Med. Insights Oncol. 2019, 13, 1179554919831990. [Google Scholar] [CrossRef] [Green Version]
- Hennig, R.; Albawardi, A.; Almarzooqi, S.; Haneefa, S.; Imbaraj, E.; Zaaba, N.E.; Nemmar, A.; Subramanya, S.; Maruta, H.; Adrian, T.E. 1,2,3-Triazolyl ester of ketorolac (15K), a potent PAK1 blocker, inhibits both growth and metastasis of orthotopic human pancreatic cancer xenografts in mice. Drug Discov. Ther. 2019, 13, 248–255. [Google Scholar] [CrossRef]
- Guo, C.; He, J.; Song, X.; Tan, L.; Wang, M.; Jiang, P.; Li, Y.; Cao, Z.; Peng, C. Pharmacological properties and derivatives of shikonin-A review in recent years. Pharmacol. Res. 2019, 149, 104463. [Google Scholar] [CrossRef]
- Zhang, N.; Peng, F.; Wang, Y.; Yang, L.; Wu, F.; Wang, X.; Ye, C.; Han, B.; He, G. Shikonin induces colorectal carcinoma cells apoptosis and autophagy by targeting galectin-1/JNK signaling axis. Int. J. Biol. Sci. 2020, 16, 147–161. [Google Scholar] [CrossRef]
- Jia, L.; Zhu, Z.; Li, H.; Li, Y. Shikonin inhibits proliferation, migration, invasion and promotes apoptosis in NCI-N87 cells via inhibition of PI3K/AKT signal pathway. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2662–2669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, W.; Cui, J.; Zhang, K.; Xi, H.; Cai, A.; Li, J.; Gao, Y.; Hu, C.; Liu, Y.; Lu, Y.; et al. Shikonin induces ROS-based mitochondria-mediated apoptosis in colon cancer. Oncotarget 2017, 8, 109094–109106. [Google Scholar] [CrossRef] [Green Version]
- Ruan, Z.; Liang, M.; Shang, L.; Lai, M.; Deng, X.; Su, X. Shikonin-mediated PD-L1 degradation suppresses immune evasion in pancreatic cancer by inhibiting NF-kappaB/STAT3 and NF-kappaB/CSN5 signaling pathways. Pancreatology 2021, 21, 630–641. [Google Scholar] [CrossRef]
- PDB Database. Available online: http://www.rcsb.org (accessed on 20 December 2021).
- Sun, X.; Wang, W.; Chen, J.; Cai, X.; Yang, J.; Yang, Y.; Yan, H.; Cheng, X.; Ye, J.; Lu, W.; et al. The Natural Diterpenoid Isoforretin A Inhibits Thioredoxin-1 and Triggers Potent ROS-Mediated Antitumor Effects. Cancer Res. 2017, 77, 926–936. [Google Scholar] [CrossRef] [Green Version]
- Chou, T.-C.; Talalay, P. Quantitative analysis of dose-effect relationships: The combined effects of multiple drugs or enzyme inhibitors. Adv. Enzym. Regul. 1984, 22, 27–55. [Google Scholar] [CrossRef]
- Chou, T.C. Theoretical basis, experimental design, and computerized simulation of synergism and antagonism in drug combination studies. Pharmacol. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef]
- Ito, M.; Codony-Servat, C.; Codony-Servat, J.; Llige, D.; Chaib, I.; Sun, X.; Miao, J.; Sun, R.; Cai, X.; Verlicchi, A.; et al. Targeting PKCiota-PAK1 signaling pathways in EGFR and KRAS mutant adenocarcinoma and lung squamous cell carcinoma. Cell Commun. Signal 2019, 17, 137. [Google Scholar] [CrossRef] [Green Version]
- Boulos, J.C.; Rahama, M.; Hegazy, M.F.; Efferth, T. Shikonin derivatives for cancer prevention and therapy. Cancer Lett. 2019, 459, 248–267. [Google Scholar] [CrossRef]
- Hilde Brems, E.B.; de Ravel, T.; Legius, E. Mechanisms in the pathogenesis of malignant tumours in neurofi bromatosis type 1. Lancet Oncol. 2009, 10, 508–515. [Google Scholar] [CrossRef]
- Maria Radu, G.S.; Kosoff1, R.; Chernoff, J. Pak Signaling in the Development and Progression of Cancer. Nat. Rev. Cancer 2014, 14, 13–25. [Google Scholar] [CrossRef]
- Xu, S.; Lei, S.L.; Liu, K.J.; Yi, S.G.; Yang, Z.L.; Yao, H.L. circSFMBT1 promotes pancreatic cancer growth and metastasis via targeting miR-330-5p/PAK1 axis. Cancer Gene. Ther. 2021, 28, 234–249. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Jubb, A.M.; Lyle, K.; Xiao, Q.; Ong, C.C.; Desai, R.; Fu, L.; Gnad, F.; Song, Q.; Haverty, P.M.; et al. PAK1 mediates pancreatic cancer cell migration and resistance to MET inhibition. J. Pathol. 2014, 234, 502–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jagadeeshan, S.; Subramanian, A.; Tentu, S.; Beesetti, S.; Singhal, M.; Raghavan, S.; Surabhi, R.P.; Mavuluri, J.; Bhoopalan, H.; Biswal, J.; et al. p21 activated kinase 1 (Pak1) signaling influences therapeutic outcome in pancreatic cancer. Ann. Oncol. 2016, 27, 1546–1556. [Google Scholar] [CrossRef]
- Chauhan, S.C.; Ebeling, M.C.; Maher, D.M.; Koch, M.D.; Watanabe, A.; Aburatani, H.; Lio, Y.; Jaggi, M. MUC13 mucin augments pancreatic tumorigenesis. Mol. Cancer Ther. 2012, 11, 24–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jagadeeshan, S.; Krishnamoorthy, Y.R.; Singhal, M.; Subramanian, A.; Mavuluri, J.; Lakshmi, A.; Roshini, A.; Baskar, G.; Ravi, M.; Joseph, L.D.; et al. Transcriptional regulation of fibronectin by p21-activated kinase-1 modulates pancreatic tumorigenesis. Oncogene 2015, 34, 455–464. [Google Scholar] [CrossRef]
- Semenova, G.; Chernoff, J. Targeting PAK1. Biochem. Soc. Trans. 2017, 45, 79–88. [Google Scholar] [CrossRef]
- Mo, L.; Xu, L.; Jia, M.; Su, B.; Hu, Y.; Hu, Z.; Li, H.; Zhao, C.; Zhao, Z.; Li, J. Shikonin suppresses the epithelial-to-mesenchymal transition by downregulating NHE1 in bladder cancer cells. J. Cancer 2021, 12, 6814–6824. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Z.Y.; Chen, L.; Zhang, J.Y.; Fu, L.Y.; Tao, L.; Zhang, Y.; Hu, X.X.; Shen, X.C. Shikonin inhibits triple-negative breast cancer-cell metastasis by reversing the epithelial-to-mesenchymal transition via glycogen synthase kinase 3beta-regulated suppression of beta-catenin signaling. Biochem. Pharmacol. 2019, 166, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fan, X.X.; Jiang, Z.B.; Loo, W.T.; Yao, X.J.; Leung, E.L.; Chow, L.W.; Liu, L. Shikonin inhibits gefitinib-resistant non-small cell lung cancer by inhibiting TrxR and activating the EGFR proteasomal degradation pathway. Pharmacol. Res. 2017, 115, 45–55. [Google Scholar] [CrossRef]
- Sun, Q.; Gong, T.; Liu, M.; Ren, S.; Yang, H.; Zeng, S.; Zhao, H.; Chen, L.; Ming, T.; Meng, X.; et al. Shikonin, a naphthalene ingredient: Therapeutic actions, pharmacokinetics, toxicology, clinical trials and pharmaceutical researches. Phytomedicine 2022, 94, 153805. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, W.; Sun, X.; Gao, Y.; Lu, M.; Zhu, L.; Wang, D.; Hu, C.; Chen, J.; Cao, P. Natural Compound Shikonin Is a Novel PAK1 Inhibitor and Enhances Efficacy of Chemotherapy against Pancreatic Cancer Cells. Molecules 2022, 27, 2747. https://doi.org/10.3390/molecules27092747
Ji W, Sun X, Gao Y, Lu M, Zhu L, Wang D, Hu C, Chen J, Cao P. Natural Compound Shikonin Is a Novel PAK1 Inhibitor and Enhances Efficacy of Chemotherapy against Pancreatic Cancer Cells. Molecules. 2022; 27(9):2747. https://doi.org/10.3390/molecules27092747
Chicago/Turabian StyleJi, Wenjing, Xiaoyan Sun, Yang Gao, Man Lu, Lingxia Zhu, Dawei Wang, Chunping Hu, Jiao Chen, and Peng Cao. 2022. "Natural Compound Shikonin Is a Novel PAK1 Inhibitor and Enhances Efficacy of Chemotherapy against Pancreatic Cancer Cells" Molecules 27, no. 9: 2747. https://doi.org/10.3390/molecules27092747