
Bioprospecting the Antibiofilm and Antimicrobial Activity of Soil and Insect Gut Bacteria

 , ,
, ,
Abstract
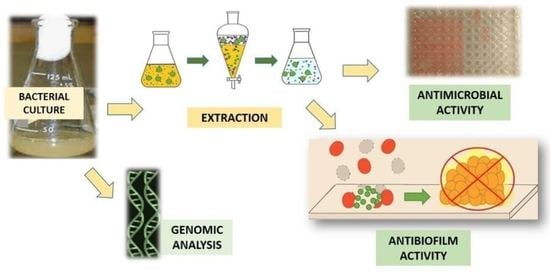
:
1. Introduction
2. Results
2.1. Antimicrobial Activity
2.2. Antibiofilm Activity
2.2.1. Biofilm Formation
2.2.2. Biofilm Inhibition Ability
2.3. Genomic Analysis of Bacillus Cereus ILB55
2.4. Thin Layer Chromatography (TLC)
3. Discussion
4. Materials and Methods
4.1. Organisms and Culture
4.2. Preparation of Extracts
4.3. Antimicrobial Assays
Determination of Minimal Inhibitory Concentration (MIC)
4.4. Antibiofilm Activity
4.4.1. Biofilm Formation Assay
4.4.2. Determination of Minimal Biofilm Inhibitory Concentration (MBIC)
4.4.3. Statistical Analysis
4.5. Genomic Analysis
4.6. Thin Layer Chromatography (TLC)
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- WHO. Global Health Estimates 2016: Deaths by Cause, Age, Sex, by Country and by Region, 2000–2016; WHO: Geneva, Switzerland, 2018.
- WHO. Causes of child mortality. In Global Health Observatory Data; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Pandey, R.; Mishra, S.K.; Shrestha, A. Characterisation of eskape pathogens with special reference to multidrug resistance and biofilm production in a nepalese hospital. Infect. Drug Resist. 2021, 14, 2201–2212. [Google Scholar] [CrossRef] [PubMed]
- Simpkin, V.L.; Renwick, M.J.; Kelly, R.; Mossialos, E. Incentivising innovation in antibiotic drug discovery and development: Progress, challenges and next steps. J. Antibiot. 2017, 70, 1087–1096. [Google Scholar] [CrossRef] [Green Version]
- Peralta, M.A.; Ortega, M.G.; Cabrera, J.L.; Paraje, M.G. The antioxidant activity of a prenyl flavonoid alters its antifungal toxicity on Candida albicans biofilms. Food Chem. Toxicol. 2018, 114, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Paraje, M.G. Antimicrobial resistance in biofilms. In Science against Microbial Pathogens: Communicating Current Research and Technological Advances; Formatex: Badajoz, Spain, 2011; Volume 2, pp. 736–744. [Google Scholar]
- Del Pozo, J.L. Biofilm-related disease. Expert Rev. Anti-Infect. Ther. 2018, 16, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Asif, M.; Hussain, T.; Ali, M.; Rafiq, M.; Atif, M. Bacterial biofilm and associated infections. J. Chin. Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, F.M.; Teixeira-Santos, R.; Mergulhão, F.J.M.; Gomes, L.C. The use of probiotics to fight biofilms in medical devices: A systematic review and meta-analysis. Microorganisms 2021, 9, 27. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.W.; Mah, T.F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, R.; Santhakumari, S.; Poonguzhali, P.; Geetha, M.; Dyavaiah, M.; Xiangmin, L. Bacterial Biofilm Inhibition: A Focused Review on Recent Therapeutic Strategies for Combating the Biofilm Mediated Infections. Front. Microbiol. 2021, 12, 676458. [Google Scholar] [CrossRef]
- Penesyan, A.; Gillings, M.; Paulsen, I.T. Antibiotic discovery: Combatting bacterial resistance in cells and in biofilm communities. Molecules 2015, 20, 5286–5298. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Geng, M.; Bai, L. Targeting biofilms therapy: Current research strategies and development hurdles. Microorganisms 2020, 8, 1222. [Google Scholar] [CrossRef]
- Divakar, S.; Lama, M.; Asad, U.K. Antibiotics versus biofilm: An emerging battleground in microbial communities. Antimicrob. Resist. Infect. Control 2019, 3, 76. [Google Scholar]
- Estevez, M.B.; Raffaelli, S.; Mitchell, S.G.; Faccio, R.; Alborés, S. Biofilm eradication using biogenic silver nanoparticles. Molecules 2020, 25, 2023. [Google Scholar] [CrossRef] [PubMed]
- Alam, K.; Farraj, D.A.A.; Mah-e-Fatima, S.; Yameen, M.A.; Elshikh, M.S.; Alkufeidy, R.M.; Mustafa, A.E.-Z.M.A.; Bhasme, P.; Alshammari, M.K.; Alkubaisi, N.A.; et al. Anti-biofilm activity of plant derived extracts against infectious pathogen-Pseudomonas aeruginosa PAO1. J. Infect. Public Health 2020, 13, 1734–1741. [Google Scholar] [CrossRef]
- Beoletto, V.G.; de las Mercedes Oliva, M.; Marioli, J.M.; Carezzano, M.E.; Demo, M.S. Antimicrobial Natural Products against Bacterial Biofilms. In Antibiotic Resistance: Mechanisms and New Antimicrobial Approaches; Academic Press: Cambridge, MA, USA, 2016; pp. 291–307. [Google Scholar]
- Song, X.; Xia, Y.; He, Z.; Zhang, H. A Review of Natural Products with Anti-Biofilm Activity. Curr. Org. Chem. 2018, 21, 789–817. [Google Scholar] [CrossRef]
- Ćirić, A.D.; Petrović, J.D.; Glamočlija, J.M.; Smiljković, M.S.; Nikolić, M.M.; Stojković, D.S.; Soković, M.D. Natural products as biofilm formation antagonists and regulators of quorum sensing functions: A comprehensive review update and future trends. S. Afr. J. Bot. 2019, 120, 65–80. [Google Scholar] [CrossRef]
- Shin, J.M.; Ateia, I.; Paulus, J.R.; Liu, H.; Fenno, J.C.; Rickard, A.H.; Kapila, Y.L. Antimicrobial nisin acts against saliva derived multi-species biofilms without cytotoxicity to human oral cells. Front. Microbiol. 2015, 6, 617. [Google Scholar] [CrossRef] [Green Version]
- Saising, J.; Dube, L.; Ziebandt, A.K.; Voravuthikunchai, S.P.; Nega, M.; Götz, F. Activity of gallidermin on Staphylococcus aureus and Staphylococcus epidermidis biofilms. Antimicrob. Agents Chemother. 2012, 56, 5804–5810. [Google Scholar] [CrossRef] [Green Version]
- Sansinenea, E.; Ortiz, A. Secondary metabolites of soil Bacillus spp. Biotechnol. Lett. 2011, 33, 1523–1538. [Google Scholar] [CrossRef]
- Clements, T.; Ndlovu, T.; Khan, W. Broad-spectrum antimicrobial activity of secondary metabolites produced by Serratia marcescens strains. Microbiol. Res. 2019, 229, 126329. [Google Scholar] [CrossRef]
- Saxena, A.K.; Kumar, M.; Chakdar, H.; Anuroopa, N.; Bagyaraj, D.J. Bacillus species in soil as a natural resource for plant health and nutrition. J. Appl. Microbiol. 2020, 128, 1583–1594. [Google Scholar] [CrossRef] [Green Version]
- Abreo, E.; Valle, D.; González, A.; Altier, N. Control of damping-off in tomato seedlings exerted by Serratia spp. strains and identification of inhibitory bacterial volatiles in vitro. Syst. Appl. Microbiol. 2021, 44, 126177. [Google Scholar] [PubMed]
- Belmadani, A.; Semlali, A.; Rouabhia, M. Dermaseptin-S1 decreases Candida albicans growth, biofilm formation and the expression of hyphal wall protein 1 and aspartic protease genes. J. Appl. Microbiol. 2018, 125, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Oshima, N.; Saito, M.; Niino, M.; Hiraishi, Y.; Ueki, K.; Okoshi, K.; Hakamatsuka, T.; Hada, N. Using Quantitative Thin-Layer Chromatography Analysis. Molecules 2022, 27, 593. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Li, Q.; Xu, Z.; Zhang, N.; Shen, Q.; Zhang, R. Responses of beneficial Bacillus amyloliquefaciens SQR9 to different soilborne fungal pathogens through the alteration of antifungal compounds production. Front. Microbiol. 2014, 5, 636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, M.; Jia, H.; Zhang, G.; Zeng, L.; Zhang, T.; Long, Y.; Lan, J.; Hu, Z.; Zeng, Z.; Wang, B.; et al. Siderophore Production by Rhizosphere Biological Control Bacteria Brevibacillus brevis GZDF3 of Pinellia ternata and Its Antifungal Effects on Candida albicans. J. Microbiol. Biotechnol. 2020, 30, 689–699. [Google Scholar] [CrossRef] [PubMed]
- Fourie, R.; Kuloyo, O.O.; Mochochoko, B.M.; Albertyn, J.; Pohl, C.H. Iron at the centre of Candida albicans interactions. Front. Cell. Infect. Microbiol. 2018, 8, 185. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.Y.; Abuduaini, X.; Mamat, N.; Yang, Q.L.; Wu, M.J.; Lin, X.R.; Wang, R.; Lin, R.R.; Zeng, W.J.; Ning, H.C.; et al. Genome sequencing and functional annotation of Bacillus sp. strain BS-Z15 isolated from cotton rhizosphere soil having antagonistic activity against Verticillium dahliae. Arch. Microbiol. 2021, 203, 1565–1575. [Google Scholar] [CrossRef]
- Tsadila, C.; Nikolaidis, M.; Dimitriou, T.G.; Kafantaris, I.; Amoutzias, G.D.; Pournaras, S.; Mossialos, D. Antibacterial activity and characterization of bacteria isolated from diverse types of greek honey against nosocomial and foodborne pathogens. Appl. Sci. 2021, 11, 5801. [Google Scholar] [CrossRef]
- Chu, J.; Wang, Y.; Zhao, B.; Zhang, X.M.; Liu, K.; Mao, L.; Kalamiyets, E. Isolation and identification of new antibacterial compounds from Bacillus pumilus. Appl. Microbiol. Biotechnol. 2019, 103, 8375–8381. [Google Scholar] [CrossRef]
- Suvega, T.; Arunkumar, K. Antimicrobial Activity of Bacteria Associated with Seaweeds against Plant Pathogens on Par with Bacteria Found in Seawater and Sediments. Br. Microbiol. Res. J. 2014, 4, 841–855. [Google Scholar] [CrossRef]
- Singla, R.K.; Dubey, A.K. Molecules and Metabolites from Natural Products as Inhibitors of Biofilm in Candida spp. pathogens. Curr. Top. Med. Chem. 2019, 19, 2567–2578. [Google Scholar] [CrossRef] [PubMed]
- Viju, N.; Punitha, S.M.J.; Satheesh, S. Antibiofilm activity of symbiotic Bacillus species associated with marine gastropods. Ann. Microbiol. 2020, 70, 11. [Google Scholar] [CrossRef]
- Ranieri, M.R.; Whitchurch, C.B.; Burrows, L.L. Mechanisms of biofilm stimulation by subinhibitory concentrations of antimicrobials. Curr. Opin. Microbiol. 2018, 45, 164–169. [Google Scholar] [CrossRef]
- CLSI. Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically. In Approved Standard, 10th ed.; CLSI: Wayne, PA, USA, 2015; p. 35. [Google Scholar]
- Marioni, J.; da Silva, M.A.; Cabrera, J.L.; Montoya, S.C.N.; Paraje, M.G. The anthraquinones rubiadin and its 1-methyl ether isolated from Heterophyllaea pustulata reduces Candida tropicalis biofilms formation. Phytomedicine 2016, 23, 1321–1328. [Google Scholar] [CrossRef] [PubMed]
- Chebbi, A.; Elshikh, M.; Haque, F.; Ahmed, S.; Dobbin, S.; Marchant, R.; Sayadi, S.; Chamkha, M.; Banat, I.M. Rhamnolipids from Pseudomonas aeruginosa strain W10; as antibiofilm/antibiofouling products for metal protection. J. Basic Microbiol. 2017, 57, 364–375. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.S.; Carvalho, J.W.P.; Aires, C.P.; Nitschke, M. Disruption of Staphylococcus aureus biofilms using rhamnolipid biosurfactants. J. Dairy Sci. 2017, 100, 7864–7873. [Google Scholar] [CrossRef] [Green Version]
- Teanpaisan, R.; Kawsud, P.; Pahumunto, N.; Puripattanavong, J. Screening for antibacterial and antibiofilm activity in Thai medicinal plant extracts against oral microorganisms. J. Tradit. Complement. Med. 2016, 7, 172–177. [Google Scholar] [CrossRef] [Green Version]
- Jork, H.; Funk, W.; Fisher, W.; Wimmer, H. Thin Layer Chromatography Reagents and Detection Methods, Vol 1a: VCH, Weinheim, 1990 (ISBN 3-527-27834-6). xv + 464 pp. Price DM 148.00. Anal. Chim. Acta 1990, 237, 51–512. [Google Scholar]
- Galand, N.; Pothier, J.; Viel, C. Plant drug analysis by planar chromatography. J. Chromatogr. Sci. 2002, 40, 585–597. [Google Scholar] [CrossRef] [Green Version]
- Pascual, M.E.; Carretero, M.E.; Slowing, K.V.; Villar, A. Simplified screening by TLC of plant drugs. Pharm. Biol. 2002, 40, 139–143. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ILBB55 | Bacillus cereus |
| ILBB44 | Bacillus pumilus |
| ILBB19 | Bacillus thuringiensis |
| ILBB139 | Bacillus wiedmannii |
| ILBB7 | Lysinibacillus fusiformis |
| ILBB63 | Lysinibacillus xylanilyticus |
| ILBB68 | Paenibacillus durus |
| ILBB15 | Peribacillus butanolivorans |
| ILB172 | Priestia megaterium |
| ILBB95 | Priestia megaterium |
| ILBB 162 | Serratia bockelmannii |
| ILBB 219 | Serratia nevei |
| ILBB 145 | Serratia ureilytica |
| CB Extract | Minimal Inhibitory Concentration (µg mL−1) | ||||||
|---|---|---|---|---|---|---|---|
| S. aureus | P. aeruginosa | E. cloacae | E. faecalis | K. pneumoniae | A. baumannii | C. albicans | |
| ILBB55 | 1250 | 312 | 2500 | NI | NI | NI | 125 |
| ILBB44 | 312 | 156 | 2500 | NI | NI | NI | NI |
| ILBB19 | 625 | 2500 | 2500 | NI | NI | 2500 | NI |
| ILBB139 | 1250 | NI | 2500 | 2500 | 2500 | NI | NI |
| ILBB7 | 625 | 625 | NI | ND | NI | NI | NI |
| ILBB63 | 625 | NI | 2500 | 625 | 2500 | NI | NI |
| ILBB68 | 1250 | NI | 2500 | 2500 | 2500 | NI | NI |
| ILBB15 | 312 | 76 | 2500 | ND | NI | NI | NI |
| ILBB172 | 625 | 312 | 2500 | 1250 | 2500 | 2500 | NI |
| ILBB95 | 1250 | 1250 | NI | 2500 | NI | 2500 | 2500 |
| ILBB162 | 156 | 63 | 76 | NI | NI | 1250 | NI |
| ILBB219 | 1250 | 2500 | 2500 | NI | NI | NI | NI |
| ILBB145 | 1250 | 250 | 1250 | NI | NI | 1250 | NI |
| C. albicans | P. aeruginosa | S. aureus | ||||
|---|---|---|---|---|---|---|
| Concentration (µg mL−1) | Inhibition (%) | Concentration (µg mL−1) | Inhibition (%) | Concentration (µg mL−1) | Inhibition (%) | |
| ILBB55 | 16 | 71 | 125 | 62 | 16 | 99 |
| ILBB44 | 16 | 60 | NI | NI | ||
| ILBB19 | 125 | 55 | NI | 32 | 42 | |
| ILBB139 | 250 | 40 | 63 | 9 | NI | |
| ILBB7 | 16 | 79 | NI | NI | ||
| ILBB63 | 16 | 64 | 500 | 68 | NI | |
| ILBB68 | 250 | 68 | 63 | 13 | NI | |
| ILBB15 | 16 | 30 | NI | NI | ||
| ILBB172 | 125 | 71 | NI | NI | ||
| ILBB95 | 500 | 93 | NI | NI | ||
| ILBB162 | 63 | 54 | NI | 250 | 77 | |
| ILBB219 | 32 | 49 | NI | 500 | 78 | |
| ILBB145 | 32 | 46 | NI | 125 | 94 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raffaelli, S.; Abreo, E.; Altier, N.; Vázquez, Á.; Alborés, S. Bioprospecting the Antibiofilm and Antimicrobial Activity of Soil and Insect Gut Bacteria. Molecules 2022, 27, 2002. https://doi.org/10.3390/molecules27062002
Raffaelli S, Abreo E, Altier N, Vázquez Á, Alborés S. Bioprospecting the Antibiofilm and Antimicrobial Activity of Soil and Insect Gut Bacteria. Molecules. 2022; 27(6):2002. https://doi.org/10.3390/molecules27062002
Chicago/Turabian StyleRaffaelli, Sofía, Eduardo Abreo, Nora Altier, Álvaro Vázquez, and Silvana Alborés. 2022. "Bioprospecting the Antibiofilm and Antimicrobial Activity of Soil and Insect Gut Bacteria" Molecules 27, no. 6: 2002. https://doi.org/10.3390/molecules27062002