
Evaluation of Vetiver Volatile Compound Production under Aeroponic-Grown Conditions for the Perfume Industry

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Impact of HPA Cultivation on Root Biomass and VOCs Accumulation
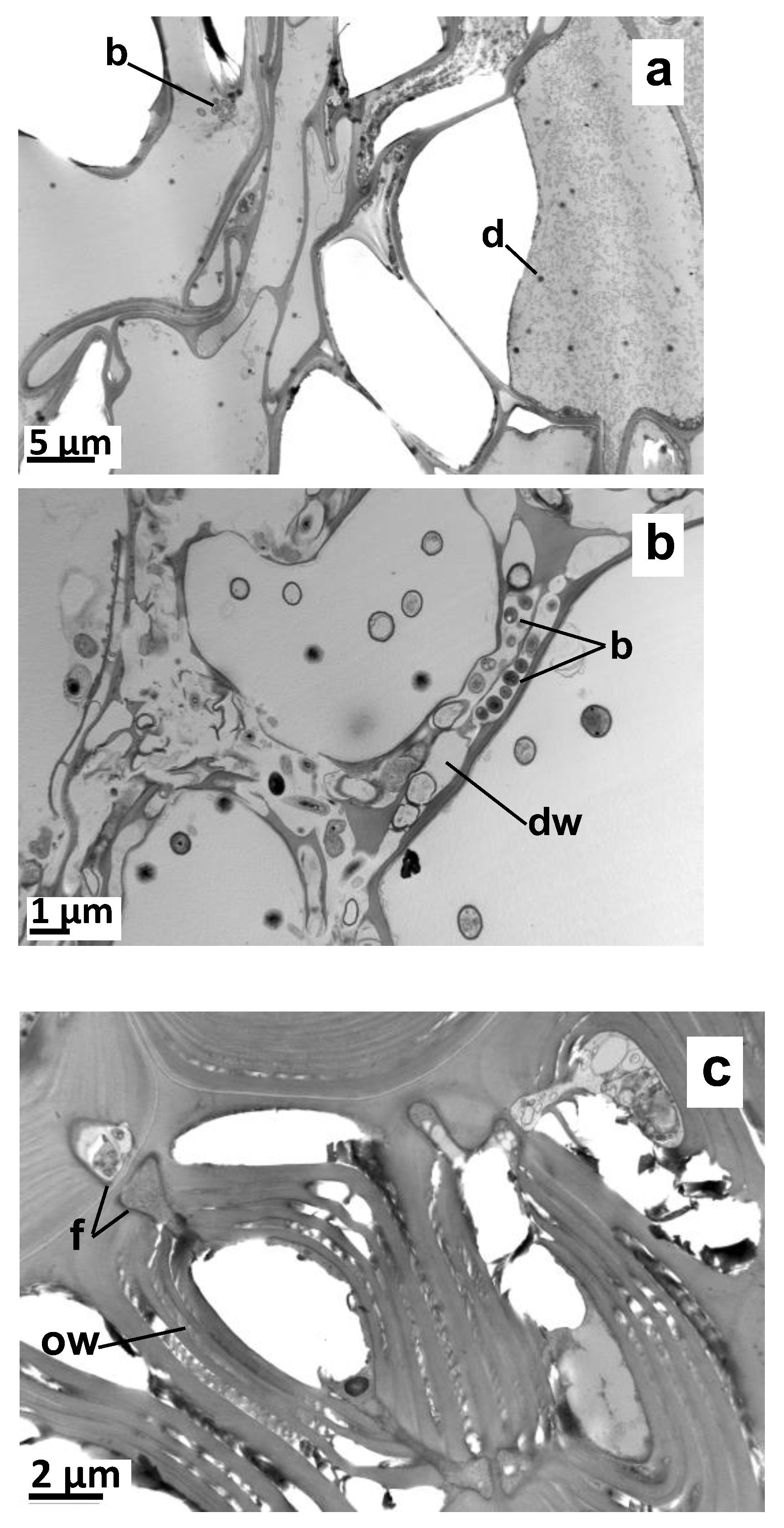
2.2. Impact of a Root-Associated Microbiome on Vetiver VOC Accumulation
2.3. Impact of Culture Conditions on Root Development
3. Materials and Methods
3.1. Plant Material and Culture Conditions
3.2. Microscopy
3.3. VOC Analyses
3.4. Hydrodistillation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Lawrence, B.M. Progress in essential oils. Perfum. Flavor. 2016, 41, 54–57. Available online: https://www.perfumerflavorist.com/fragrance/ingredients/article/21856205/progress-in-essential-oils-october-2016 (accessed on 5 February 2022).
- Maffei, M. Vetiveria, the Genus Vetiveria, 1st ed.; Taylor & Francis: London, UK, 2002; pp. 1–202. [Google Scholar]
- Grover, M.; Behl, T.; Virmani, T.; Bhatia, S.; Al-Harrasi, A.; Aleya, L. Chrysopogon zizanioides—A review on its pharmacognosy, chemical composition and pharmacological activities. Environ. Sci. Pollut. Res. 2021, 28, 44667–44692. [Google Scholar] [CrossRef]
- Lunz, K.; Stappen, I. Back to the Roots—An Overview of the Chemical Composition and Bioactivity of Selected Root-Essential Oils. Molecules 2021, 26, 3155. [Google Scholar] [CrossRef]
- Demole, E.P.; Holzner, G.W.; Youssefi, M.J. Malodor formation in alcoholic perfumes containing vetiveryl acetate and vetiver oil. Perfum. Flavor. 1995, 20, 35–40. Available online: https://www.perfumerflavorist.com/fragrance/fine-fragrance/article/21860806/malodor-formation-in-alcoholic-perfumes-containing-vetiveryl-acetate-and-vetiver-oil (accessed on 5 February 2022).
- Belhassen, E.; Filippi, J.J.; Brévard, H.; Joulain, D.; Baldovini, N. Volatile constituents of vetiver: A review. Flavour. Fragr. J. 2015, 30, 26–83. [Google Scholar] [CrossRef]
- Belhassen, E.; Baldovini, N.; Brevard, H.; Meierhenrich, U.J.; Filippi, J.J. Unravelling the scent of vetiver: Identification of character impact compounds. Chem. Biodivers. 2014, 11, 1821–1842. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, J.; Bae, H.; Jordi, S.; Dao, Q.M.; Dossenbach, S.; Dehn, S.; Lingnau, J.B.; De, C.K.; Kraft, P.; List, B. The Smelling Principle of Vetiver Oil, Unveiled by Chemical Synthesis. Angew. Chem. Int. Ed. 2021, 60, 5666. [Google Scholar] [CrossRef]
- Martinez, J.; Rosa, P.T.V.; Menut, C.; Leydet, A.; Brat, P.; Pallet, D.; Meireles, M.A.A. Valorization of Brazilian vetiver (Vetiveria zizanioides (L.) Nash ex Small) oil. J. Agric. Food Chem. 2004, 52, 6578–6584. [Google Scholar] [CrossRef]
- Champagnat, P.; Figueredo, G.; Chalchat, J.C.; Carnat, A.P.; Bessière, J.M. A study on the composition of commercial Vetiveria zizanioides oils from different geographical origins. J. Essent. Oil Res. 2006, 18, 416–422. [Google Scholar] [CrossRef]
- Pripdeevech, P.; Wongpornchai, S.; Marriott, P.J. Comprehensive two-dimensional gas chromatography–mass spectrometry analysis of volatile constituents in Thai vetiver root oils obtained by using different extraction methods. Phytochem. Anal. 2010, 21, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Despommier, D.D. The Vertical Farm: Feeding the World in the 21st Century, 1st ed.; Thomas Dunne Books, St. Martin’s Press: New York, NY, USA, 2010; pp. 1–305. [Google Scholar]
- Hussain, A.; Iqbal, K.; Aziem, S.; Mahato, P.; Negi, A.K. A review on the science of growing crops without soil (soilless culture) —A novel alternative for growing crops. Int. J. Agric. Crop Sci. 2014, 7, 833–842. [Google Scholar]
- Van Gerrewey, T.; Boon, N.; Geelen, D. Vertical Farming: The OnlyWay Is up? Agronomy 2022, 12, 2. [Google Scholar] [CrossRef]
- Lakhiar, I.A.; Gao, J.; Syed, T.N.; Chandio, F.A.; Tunio, M.H.; Ahmad, F.; Solangi, K.A. Overview of the aeroponic agriculture—An emerging technology for global food security. Int. J. Agric. Biol. Eng. 2020, 13, 1–10. [Google Scholar] [CrossRef]
- Vinci, G.; Rapa, M. Hydroponic cultivation: Life cycle assessment of substrate choice. Br. Food J. 2019, 121, 1801–1812. [Google Scholar] [CrossRef]
- Duriot, L.; Salwinski, A.; Rangoni, L. Root Extracts from Plants of the Morus Genus and Uses of Same. W.O. Patent No. WO2019/068824, 11 April 2019. [Google Scholar]
- Shoji, Y.; Farrance, C.E.; Bautista, J.; Bi, H.; Musiychuk, K.; Horsey, A.; Park, H.; Jaje, J.; Green, B.J.; Shamloul, M.; et al. A plant-based system for rapid production of influenza vaccine antigens. Influenza Other Respir. Viruses 2012, 6, 204–210. [Google Scholar] [CrossRef]
- Honary, H.; Vasundhara, M.; Nuthan, D. Hydroponics and aeroponics as alternative production systems for high-value medicinal and aromatic crops: Present scenario and future prospects. J. Med. Aromat. Plants 2011, 33, 397–403. [Google Scholar]
- Movahedi, Z.; Rostami, M. Production of Some Medicinal Plants in Aeroponic System. J. Med. Plants By-Prod. 2020, 1, 91–99. [Google Scholar] [CrossRef]
- Partap, M.; Rattan, S.; Kanika; Ashrita; Sood, A.; Kumar, P.; Warghat, A.R. Hydroponic and Aeroponic Cultivation of Economically Important Crops for Production of Quality Biomass. In Agricultural Biotechnology: Latest Research and Trends, 1st ed.; Srivastava, D.K., Thakur, A.K., Kumar, P., Eds.; Springer: Singapore, 2021; pp. 573–585. [Google Scholar] [CrossRef]
- Pripdeevech, P.; Wongpornchai, S.; Promsiri, A. Highly volatile constituents of Vetiveria zizanioides roots grown under different cultivation conditions. Molecules 2006, 11, 817–826. [Google Scholar] [CrossRef] [Green Version]
- Freeman, S. Vetiver in Southwest Haiti; Haiti Research and Policy Program; Columbia University: New York, NY, USA, 2011; Available online: http://haiti.ciesin.columbia.edu/haiti_files/documents/Freeman_UNEP_Vetiver_Report_2011_0.pdf (accessed on 5 February 2022).
- Maia, N.B.; Bovi, O.A.; Zullo, M.A.T.; Perecin, M.B.; Granja, N.P.; Carmello, Q.A.C.; Robaina, C.; Coll, F. Hydroponic Cultivation of Mint and Vetiver with Spirotane Analogues of Brassinosteroids. Acta Hortic. 2004, 644, 55–59. [Google Scholar] [CrossRef]
- Mazzoni, V.; Tomi, F.; Casanova, J. A daucane-type sesquiterpene from Daucus carota seed oil. Flavour Fragr. J. 1999, 14, 268–272. [Google Scholar] [CrossRef]
- Kovacevic, N.N.; Marcetic, M.D.; Lakusic, D.V.; Lakusic, B.S. Composition of the Essential Oils of Different Parts of Seseli annuum L. (Apiaceae). J. Essent. Oil Bear. Plants 2016, 19, 671–677. [Google Scholar] [CrossRef]
- Aladedunye, F.A.; Benn, M.H.; Okorie, D.A. Sesquiterpenes from Culcasia scandens P. Beauv. Nat. Prod. Res. 2008, 22, 879–883. [Google Scholar] [CrossRef] [PubMed]
- Bohlmann, F.; Zdero, C. Glaucolides and other constituents from south african Vernonia species. Phytochemistry 1982, 21, 2263–2267. [Google Scholar] [CrossRef]
- Cassidy, M.P.; Ghisalberti, E.L. New Terpene Hydrocarbons from the Sponge Higginsia sp. J. Nat. Prod. 1993, 56, 1190–1193. [Google Scholar] [CrossRef]
- Castro, V.; Jakupovic, J.; Bohlmann, F. A New Type of Sesquiterpene and Acorane Derivative from Calea prunifolia. J. Nat. Prod. 1984, 47, 802–808. [Google Scholar] [CrossRef]
- Marques, A.P.S.; Bonfim, F.P.G.; Dantas, W.F.C.; Puppi, R.J.; Marques, M.O.M. Chemical composition of essential oil from Varronia curassavica Jacq. accessions in different seasons of the year. Ind. Crops Prod. 2019, 140, 111656. [Google Scholar] [CrossRef]
- Cool, L.G. ent-Daucane and acorane sesquiterpenes from ×Cupressocyparis leylandii foliage. Phytochemistry 2001, 58, 969–972. [Google Scholar] [CrossRef]
- Saad, H.M.; Rahman, S.N.S.A.; Navanesan, S.; Tan, C.H.; Manickam, S.; Malek, S.N.A.M.; Sim, K.S. Evaluation of antioxidant activity and phytochemical composition of Baeckea frutescens and Leptospermum javanicum essential oils. S. Afr. J. Bot. 2021, 141, 474–479. [Google Scholar] [CrossRef]
- Hashidoko, Y.; Tahara, S.; Mizutani, J. Sesquiterpene Hydrocarbons in Glandular Trichome Exudate of Rosa rugosa Leaves. Z. Nat. C 1992, 47, 353–359. [Google Scholar] [CrossRef]
- Nogueira, J.; Mourão, S.; Dolabela, I.B.; Santos, M.G.; Mello, C.; Kelecom, A.; Mexas, R.; Feder, D.; Fernandes, C.P.; González, M.; et al. Zanthoxylum caribaeum (Rutaceae) essential oil: Chemical investigation and biological effects on Rhodnius prolixus nymph. Parasitol. Res. 2014, 113, 4271–4279. [Google Scholar] [CrossRef] [PubMed]
- Bezerra-Silva, P.C.; Dutra, K.A.; Santos, G.K.N.; Silva, R.C.S.; Iulek, J.; Milet-Pinheiro, P.; Navarro, D.M.A.F. Evaluation of the Activity of the Essential Oil from an Ornamental Flower against Aedes aegypti: Electrophysiology, Molecular Dynamics and Behavioral Assays. PLoS ONE 2016, 11, e0150008. [Google Scholar] [CrossRef] [PubMed]
- Sainz, P.; Andrés, M.F.; Martínez-Díaz, R.A.; Bailén, M.; Navarro-Rocha, J.; Díaz, C.E.; González-Coloma, A. Chemical Composition and Biological Activities of Artemisia pedemontana subsp. assoana Essential Oils and Hydrolate. Biomolecules 2019, 9, 558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; pp. 1–804. [Google Scholar]
- Farsam, H.; Amanlou, M.; Taghi-Cheetsaz, N.; Amin, G.; Salehi-Sormaghi, M.H. Essential oil constituents of Chimonanthus fragrans flowers population of Tehran. Daru 2007, 15, 129–131. [Google Scholar]
- Kahriman, N.; Tosun Çelik, G.; Terzioğlu, S.; Alpay Karaoğlu, Ş.; Yayh, N. Chemical Composition and Antimicrobial Activity of the Essential Oils from the Flower, Leaf, and Stem of Senecio pandurifolius. Rec. Nat. Prod. 2011, 52, 82–91. [Google Scholar]
- Del Giudice, L.; Massardo, D.R.; Pontieri, P.; Bertea, C.M.; Mombello, D.; Carata, E.; Tredici, S.M.; Talà, A.; Mucciarelli, M.; Groudeva, V.I.; et al. The microbial community of vetiver root and its involvement into essential oil biogenesis. Environ. Microbiol. 2008, 10, 2824–2841. [Google Scholar] [CrossRef] [PubMed]
- Tracy, S.R.; Black, C.R.; Roberts, J.A.; Mooney, S.J. Soil compaction: A review of past and present techniques for investigating effects on root growth. J. Sci. Food Agric. 2011, 91, 1528–1537. [Google Scholar] [CrossRef] [PubMed]
- Unger, P.W.; Kaspar, T.C. Soil compaction and root growth: A review. Agron. J. 1994, 86, 759–766. [Google Scholar] [CrossRef]
- Merotto, A.J.; Mundstock, C.M. Wheat root growth as affected by soil strength. Rev. Bras. Ciência Solo 1999, 23, 197–202. [Google Scholar]
- Rich, S.M.; Watt, M. Soil conditions and cereal root system architecture: Review and considerations for linking Darwin and Weaver. J. Exp. Bot. 2013, 64, 1193–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brundrett, M.C.; Kendrick, B.; Peterson, C.A. Efficient lipid staining in plant material with Sudan Red 7B or Fluoral Yellow 088 in polyethylene glycolglycerol. Biotechol. Histochem. 1991, 66, 111–116. [Google Scholar] [CrossRef]
- Danh, L.T. Supercritical CO2 Extraction of Vetiver Essential Oil and Economic Incentive for Use of Vetiver Grass in Phytoremediation. Ph.D. Thesis, The University of New South Wales, Sydney, Australia, 2010. [Google Scholar]
- Lavania, U. Towards bio-efficient and non-invasive vetiver: Lessons from genomic manipulation and chromosomal characterization. In Proceedings of the Fourth International Conference on Vetiver and Exhibition, Caracas, Venezuela, 22–26 October 2006. [Google Scholar]
- Kartusch, R.; Kartusch, B. Nachweis und lokalisierung der atherischen ole in der wurzel von Vetiveria zizanioides L. Mikroskopie 1978, 34, 195–201. [Google Scholar]
- Viano, J.; Smadja, J.; Conan, J.Y.; Gaydou, E.M. Ultrastructure of roots of Vetiveria zizanioides (L.) Stapf (Gramineae). Bull. Mus. Natl. Hist. Nat. Sect. B Adansonia 1991, 13, 61–69. [Google Scholar]
- Meyer, C.J.; Seago, J.L.; Peterson, J.C.A. Environmental effects on the maturation of the endodermis and multiseriate exodermis of Iris germanica roots. Ann. Bot. 2009, 103, 687–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayden, A.L.; Brigham, L.A.; Giacomelli, G.A. Aeroponic cultivation of ginger (Zingiber officinale) rhizomes. Acta Hortic. 2004, 659, 397–402. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- NIST. Mass Spectral Library Standard Reference Database 1, NIST 11. Standard Reference Data Program; National Institute of Standards and Technology: Gaithersburg, MD, USA, 2011.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N° a | Compound Names b | RIZB-5MS c | RIref d | In Vitro | HPA | Potting Soil |
|---|---|---|---|---|---|---|
| 1 | 117 188 e | 1348 | - | - | - | × |
| 2 | Cyclosativene | 1366 | 1369 f | - | - | × |
| 3 | 2-epi-α-Funebrene | 1379 | 1380 f | - | × | × |
| 4 | β-Elemene | 1386 | 1389 f | - | × | - |
| 5 | 91 131 188 e | 1404 | - | - | × | - |
| 6 | 2-epi-β-Funebrene | 1411 | 1411 f | - | × | × |
| 7 | β-Cedrene | 1419 | 1419 f | - | × | × |
| 8 | γ-Elemene * | 1427 | 1434 f | - | × | - |
| 9 | EpiBicyclosesqui-phellandrene | 1428 | 1430 g | - | - | × |
| 10 | 6,9-Guaiadiene | 1440 | 1442 f | - | - | × |
| 11 | Prezizaene * | 1446 | 1444 f | - | × | × |
| 12 | Khusimene | 1451 | 1453 f | - | × | × |
| 13 | α-Acoradiene | 1462 | 1464 f | - | × | - |
| 14 | 119 202 e | 1463 | - | - | - | × |
| 15 | Dehydro-aromadendrene | 1467 | 1465 h | - | - | × |
| 16 | α-Amorphene | 1477 | 1483 f | - | - | × |
| 17 | δ-Selinene | 1485 | 1492 f | - | - | × |
| 18 | γ-Amorphene | 1489 | 1495 f | - | - | × |
| 19 | Isodaucene | 1496 | 1500 f | - | × | - |
| 20 | β-Himachalene | 1501 | 1500 f | - | - | × |
| 21 | δ-Amorphene * | 1518 | 1511 f | - | - | × |
| 22 | 41 69 105 204 e | 1520 | - | - | - | × |
| 23 | 105 133 161 189 204 e | 1529 | - | - | - | × |
| 24 | 105 161 204 e | 1535 | - | - | - | × |
| 25 | β-Vetivenene | 1550 | 1554 f | - | - | × |
| 26 | Germacrene B | 1553 | 1559 f | - | × | × |
| 27 | 105 133 189 204 e | 1558 | - | - | - | × |
| 28 | 152 222 e | 1570 | - | - | - | × |
| 29 | 187 202 e | 1581 | - | - | - | × |
| 30 | 108 204 e | 1600 | - | - | - | × |
| 31 | 187 202 e | 1610 | - | - | - | × |
| 32 | 81 222 e | 1619 | - | - | - | × |
| 33 | α-Cadinol * | 1651 | 1652 f | - | - | × |
| 34 | epi-Zizanone * | 1667 | 1668 f | - | × | × |
| 35 | 131 150 220 e | 1674 | - | - | - | × |
| 36 | 41 177 222 e | 1684 | - | - | - | × |
| 37 | 119 218 e | 1689 | - | - | - | × |
| 38 | 41 67 79 93 218 e | 1691 | - | - | × | - |
| 39 | 41 91 222 e | 1694 | - | - | - | × |
| 40 | Vetiselinenol * | 1720 | 1730 f | - | - | × |
| 41 | 83 119 220 e | 1728 | - | - | - | × |
| 42 | Khusimol | 1740 | 1741 f | - | × | × |
| 43 | (E)-Isovalencenol * | 1785 | 1793 f | - | - | × |
| 44 | 93 105 120 220 e | 1789 | - | - | - | × |
| 45 | β-Vetivone * | 1810 | 1822 f | - | - | × |
| 46 | Vetivenic acid | 1816 | 1811 f | - | - | × |
| 47 | 41 91 105 218 e | 1822 | - | - | - | × |
| 48 | α-Vetivone | 1835 | 1842 f | - | - | × |
| 49 | 91 147 218 e | 1893 | - | - | - | × |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gavira, C.; Watteau, F.; Lainé, J.-M.; Bourgaud, F.; Legendre, L. Evaluation of Vetiver Volatile Compound Production under Aeroponic-Grown Conditions for the Perfume Industry. Molecules 2022, 27, 1942. https://doi.org/10.3390/molecules27061942
Gavira C, Watteau F, Lainé J-M, Bourgaud F, Legendre L. Evaluation of Vetiver Volatile Compound Production under Aeroponic-Grown Conditions for the Perfume Industry. Molecules. 2022; 27(6):1942. https://doi.org/10.3390/molecules27061942
Chicago/Turabian StyleGavira, Carole, Françoise Watteau, Jean-Marc Lainé, Frédéric Bourgaud, and Laurent Legendre. 2022. "Evaluation of Vetiver Volatile Compound Production under Aeroponic-Grown Conditions for the Perfume Industry" Molecules 27, no. 6: 1942. https://doi.org/10.3390/molecules27061942