1. Introduction
Iodine is an essential element necessary for the proper development and functioning of the human and animal organism. Its major role is related to the function it plays in the proper functioning of the thyroid gland [
1]. Iodine is a substrate necessary for the synthesis of thyroid hormones, thyroxine-T4 and triiodothyronine-T3 [
1,
2,
3]. Pregnant women are particularly at risk of deficiency of this element, and at the same time the most needy of it. Iodine deficiency during pregnancy is associated with health consequences for the baby, such as impaired fetal brain development, leading to wider and irreversible changes such as cretinism disease. In some cases, insufficient daily dose of iodine in pregnant women can lead to infertility or miscarriage [
4,
5]. Another health consequence of iodine deficiency is the so-called hypertrophy of the thyroid gland, endemic goiter and increased probability of developing thyroid and stomach cancer [
6,
7]. The daily dose of iodine is 200–250 µg for pregnant women, 150 µg for adults, 90–120 µg for children from 5–12 years of age, under 5 years old 90 µg, respectively [
1,
6,
7,
8].
Around 2 billion people in the world suffer from iodine deficiency in the diet (hidden hunger of this microelement), and about 50 million are diagnosed with an affliction caused by this microelement deficiency [
5,
9]. The problem with insufficient iodine intake in the diet and the consequences associated with it occur in economically developed countries such as England, Germany, Australia and Italy [
10,
11]. Even though iodization has shown a great effect, iodine deficiency often persists. Moreover, WHO restrictions on limiting the consumption of table salt (iodization of table salt as one of the main programs to challenge the deficiency of this element in the diet), justified by the possibility of increasing hypertension (especially in pregnant women), initiated many studies and programs on alternative methods of implementation iodine in the human diet; one of them is the biofortification of crops [
12,
13]. Agrotechnical methods, soil fertilization, foliar application or genetic methods (plant breeding) also constitute cost-effective biofortification strategies [
9,
14,
15,
16]. A balanced diet is the basis for supplementing all the macro-/microelements and vitamins needed for proper functioning and the core of this balanced diet are vegetables, fruits and grain products [
17,
18].
Researches on the methods of effective enrichment of plants with iodine takes into account the type of application (foliar application, soil application, fertigation in field and soilless crops, hydroponics), the type of compound applied—inorganic compounds (such as KI, KIO
3) and organic compounds (2IBeA, 5ISA, 3,5-di-ISA, 4IBeA)—and plant type in terms of iodine enrichment into edible parts (fruit, grain, leaf, root) [
19,
20,
21,
22]. The research objectives and research hypotheses as well as the selection of these factors are different in individual research works. The highest and most effective degree of iodine enrichment is achieved by leafy vegetables [
23,
24,
25]. Corn, wheat and rice can also be effectively biofortified with iodine, the achievement of which was shown by Cakmak et al. 2017 [
22].
Information and current knowledge about the largest reservoir of iodine on Earth, which is seawater and the organisms that live in it, was a base for further study on the possibility of implementing that mechanism on terrestrial plants and combining it with sweet corn biofortification. Iodine volatilization from both the marine and terrestrial environments is a major component of the iodine biogeochemical cycle. There are two pathways of iodine volatilization in the marine environment—photochemical and the dominant biological one from micro- and macro-algae [
26,
27,
28]. The capture and accumulation of iodine by algae is mediated by the enzyme vanadium-dependent haloperoxidase (vHPO) [
29]. Vanadium-dependent haloperoxidase oxidizes halides and participates in the synthesis of organohalogen [
28,
30]. Vanadium-dependent haloperoxidase, whose prosthetic group is occupied by vanadium (vHIPO), improves iodine binding by catalyzing the oxidation of I
- to more lipophilic compounds (iodine(I) acid) HIO and subsequently molecular I
2. These molecules easily diffuse across cell membranes into the cytosol. The process of further reduction of HIO or I
2 to I
− in the apoplast is not yet well known, same as the presence of vanadium-dependent iodoperoxidase in higher plants [
29,
31,
32]. Smoleń et al. 2020 conducted research on vanadium-dependent iodine peroxidase activity in lettuce plants [
20]. The level of vHPO activity in corn plants at an early stage of their development after soil application of ammonium metavanadate together with organic (5ISA, 2IBeA) and inorganic iodine compounds (KI, KIO
3) was also tested by Grzanka et al. [
33].
Sweet corn grain is rich in nutritional value, it contains large amounts of protein and most vitamins and microelements. The valuable components of sweet corn grains also include such microelements as selenium, chromium, zinc, copper, nickel and iron. It is reasonable to add such an important micronutrient as iodine to this list, since maize for dry grain is one of the three most important cereals worldwide for more than 200 million people [
34].
The aim of the research was to obtain an effective level of iodine in corn grain and to evaluate a better method of iodine foliar biofortification with the combined use of inorganic (KI, KIO3) and organic iodine compounds (5ISA, 2IBeA) with vanadium. Moreover, the aim was to determine the effectiveness of iodine enrichment of sweet corn grains by foliar fertilization at various stages of plant development and expand knowledge about the content of iodosalicylates and 3,5-diiodosalicylic acid, iodobenzoates, 2,3,5-triiodobenzoic acid and iodide and iodate after foliar application of organic and inorganic iodine compounds to create a basis for iodine synthesis/transformation in maize plants to answer the question of whether a land plant like corn will be able to create an iodine uptake mechanism stimulated by the application of exogenous vanadium.
3. Discussion
Corn is a very important element in the diet of people all over the world, regardless of the level of economic development of the countries, and it is an important energy crop [
35]. Corn and food products derived from it are of great importance for people intolerant to gluten and with celiac disease. Around 1% of the world’s population suffers from this type of disorder [
36,
37]. The implementation of iodine enrichment of crops (corn, cereals, vegetables) in agricultural practice would reduce problems with iodine deficiency occurring all over the world [
5,
7,
10]. The last two decades have provided a lot of research on agrotechnical methods of enriching crops with iodine [
6,
11]. Many of them proved the effectiveness of the iodine compounds used, which were mainly KI and KIO
3 [
22,
38]. There had been available results of studies on the biofortification of crops into iodine with the use of organic compounds of this element, including 5-iodosalicylic acid, 3,5-diiodosalicylic acid, 2-iodobenzoic acid, 4-iodobenzoic acid or iodoacetic acid in tomato cultivation [
19], lettuce [
39,
40] and spinach [
41]. It is also known that applying iodine in the fertilization of plants in combination with organic stabilizers. In the research of Rangel et al. 2020, the effect of enriching lettuce with iodine was obtained after using the chitosan-I complex (Cs-KIO
3, Ch-KI) [
42].
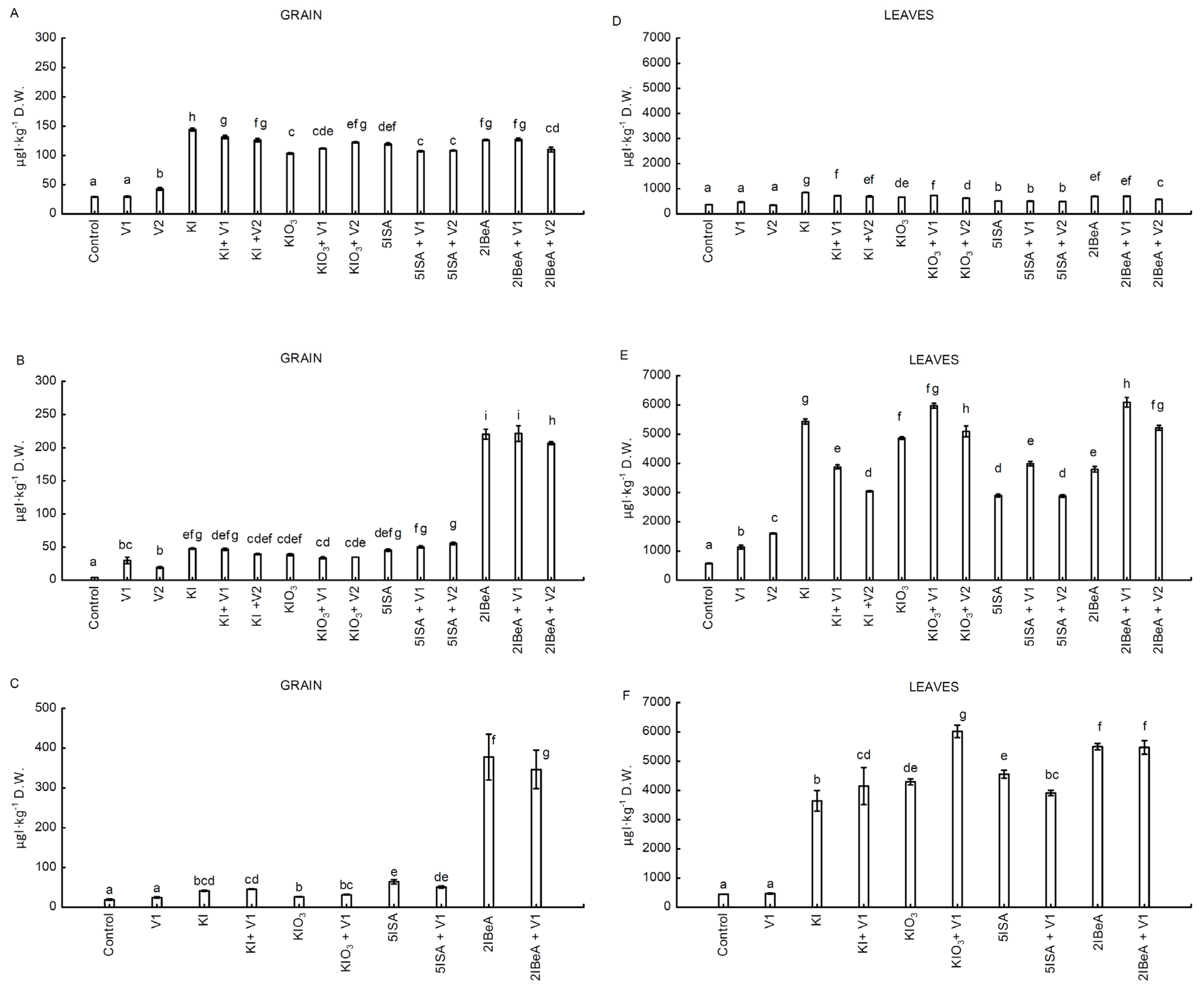
The studies determined the interaction of two trace elements, iodine and vanadium, in corn plants. This interaction—the participation of vanadium in iodine uptake—is described for several species of sea algae [
29,
31,
43]. All of the tested combinations of iodine and iodine with vanadium in the three carried out experiments significantly influenced the degree of iodine accumulation in leaves and grains of sweet corn. Effective enrichment of sweet corn grains with iodine was demonstrated after foliar application of both organic (5ISA, 2IBeA) and inorganic (KI, KIO
3) iodine compounds. The highest effect of enriching sweet corn grain with iodine was obtained after the application of the organic iodine compound 2IBeA in two experiments, No. 2 and No. 3 (analyzing only the effectiveness of iodine compounds applied individually, without vanadium). On the other hand, in experiment No. 1, the inorganic KI compound turned out to be the most effective, and the second in line was the organic 2IBeA. The conducted experiments allowed three aspects determining the effectiveness of enrichment of grains with iodine to be documented, without leading to any phytotoxic symptoms of iodine and vanadium on plants. These include the issue of increasing the dose of iodine from 10 µM I (experiment No. 1) to 100 µM I (experiment No. 2 and 3), compressing the application period between subsequent foliar applications treatments, and carrying out foliar application I and V in various phases of plant development. This is the performance of foliar application I and V at the beginning of flowering (experiment No. 3) doubled the level of iodine accumulation in grains compared to the application of these elements in the earlier stage of plant development (experiment No. 2). The grains of sweet corn treated only with 2IBeA in experiment No. 3 contained about 2.6 times more iodine than in the combination of KI from experiment No. 1 and 1.8 times more than after the application of 2IBeA in experiment No. 2. Satisfactory effects of enriching tomato fruits (generative organs) with iodine after 2IBeA application were obtained by Halka et al. 2019 [
44]. In the experiment with tomato in the seedling phase after soil application of iodine compounds, both the roots, leaves and shoots after applying KI had the highest content of iodine, followed by plants treated with 2IBeA and 4-IBeA (4-iodobenzoic acid) [
19].
From the point of view of an effective implementation program of foliar iodine biofortification, the 100 µM iodine dose used in experiment No. 3 was safe for plants and did not cause phytotoxic symptoms on sweet corn plants. Phytotoxic symptoms were demonstrated in studies with the application of inorganic forms of iodine in rice plants in doses of 10 and 100 µM KI and 100 µM KIO
3 [
45] or while fertigation 2.34 mM KIO
3 and 3.01 mM KI of corn, barley, potato and tomato [
46]. In the experiment with the use of organo-iodine compounds 3,5-diISA (3,5-diiodosalicic acid) and 4-IBeA at a dose of 25 μM I in tomato plants due to fertigation in the seedling phase, after the applied organo-iodine compounds a negative effect on the growth and development of above-ground young tomato plants parts was observed [
19]. 5ISA in a dose of 40 μM I led to a significant decrease in the biomass of roots and leaves of lettuce grown in the NFT hydroponic system [
39].
The used compounds in all experiments with both a lower dose (No. 1) and a higher dose of iodine (No. 2 and No. 3) did not have a statistically significant effect on the sweet corn yield; similar results after soil and foliar application of KI and KIO
3 were obtained Cakmak et al. [
22]. In the pot tests on sweet corn, no toxic effect was found and no statistically significant effect on the yield and development of young corn plants at a lower iodine dose of 10 µM I and analogously doses of vanadium 0.1 µM and 1 µM V [
43]. The iodine content in the leaves in all experiments was on average 14 times higher than in the grains. The higher dose of iodine compounds used significantly increased the content of iodine in the leaves by about 6 times in experiment No. 2 and No. 3 compared to No. 1. Vegetative parts of plants (leaves, shoots) accumulate higher amounts of this element than the generative parts. Similar results were obtained in studies on rice [
22,
45], sweet corn [
43], green beans [
47] and plum trees [
21].
The transport of iodine from the leaves to the corn grains after foliar application would have to be via the phloem. The phloem transport of iodine has been confirmed in many studies on the effectiveness of the accumulation of this element in the generative parts [
21,
38,
48]. Nevertheless, xylem transport of iodine is much more efficient. Hurtevent et al. (2013) pointed to the fact of the relative mobility of iodine in the phloem which was proven in the research by Zou et al. 2019 [
49,
50]. Foliar iodine application (inorganic KIO
3) in the wheat grain filling stage created an available pool of iodine in the leaf tissues, causing phloem transport to the grain. In this phase, there is an intense transfer of photo assimilates to the seeds, and the activation of the phloem transport [
50]. The presented research in experiment No. 3 also confirmed the described effects of effective phloem transport as a consequence of more effective accumulation of iodine in grains.
The combined application of iodine with other micronutrients such as Zn, Fe, Se has been described in studies on wheat and rice [
9,
50]. Foliar application of I, Zn, Se and Fe combined (in most locations where it was performed) resulted in less iodine accumulation than the same dose of iodine applied individually [
50]. In the case of vanadium, the interaction of vanadium with iodine, which could be the stimulation or antagonism of this element, is not known in higher plants yet. In a group, brown algae, enzymes as vanadium-dependent haloperoxidases (vHPO) play a key role in the uptake and accumulation of iodine (as well halides metabolism of Cl and Br) [
29,
31]. They are mainly responsible for the capture of iodine from the water, the synthesis of hydrogen halides in marine algae and antioxidant defense [
32,
51]. The studies on the interaction of iodine and vanadium in higher plants was conducted, among others, on lettuce by Smoleń et al. [
20,
40] and in sweet corn by Grzanka at al. [
33,
43].
In all experiments, in sweet corn leaves the stimulating effect of vanadium on iodine accumulation was preeminent. In experiments No. 1, No. 2 and No. 3, the application of KIO3 + V1 was statistically significantly more effective in terms of iodine accumulation than the application individually of KIO3. For 2IBeA in experiment No. 1 and No. 3, iodine accumulation in 2IBeA + V1 was at a similar level of significance with 2IBeA; in experiment No. 2 both doses (0.1 µM and 1 µM) of vanadium significantly increased I accumulation compared to the individually application of 2IBeA. In experiment No. 1, combined application 5ISA with vanadium did not show any impact on the accumulation of iodine. In experiment No. 2, after using the combination of 5ISA + V1, the iodine content in the leaves was significantly higher than after application of only 5ISA. In turn, in experiment No. 3, only the combination of 5-iodosalicylic acid with vanadium resulted in a decrease in the accumulation of iodine in the leaves (and in grain) compared to the individually applied of 5ISA.
A similar effect was obtained in a pot experiment with soil application of 5ISA with two doses of vanadium (0.1 µM and 1 µM) in sweet corn cultivation [
43]. The combined application of iodine and vanadium in a hydroponic system with organic and inorganic iodine compounds gave a variable effect. Lettuce grown in the hydroponic system in the combination of 5ISA + V and for 3,5-diISA + V resulted in a significantly higher level of iodine accumulation in the roots than the individual application of organic compounds 5ISA and 3,5-diISA. In the lettuce leaves the stimulating effect for both compounds used in hydroponics and as well in peat substrate was not observed. On the other hand, in the mineral soil, lettuce leaves had a statistically significantly higher iodine content in/after treatment with 3,5-diISA + V vs. 3,5-diISA [
40].
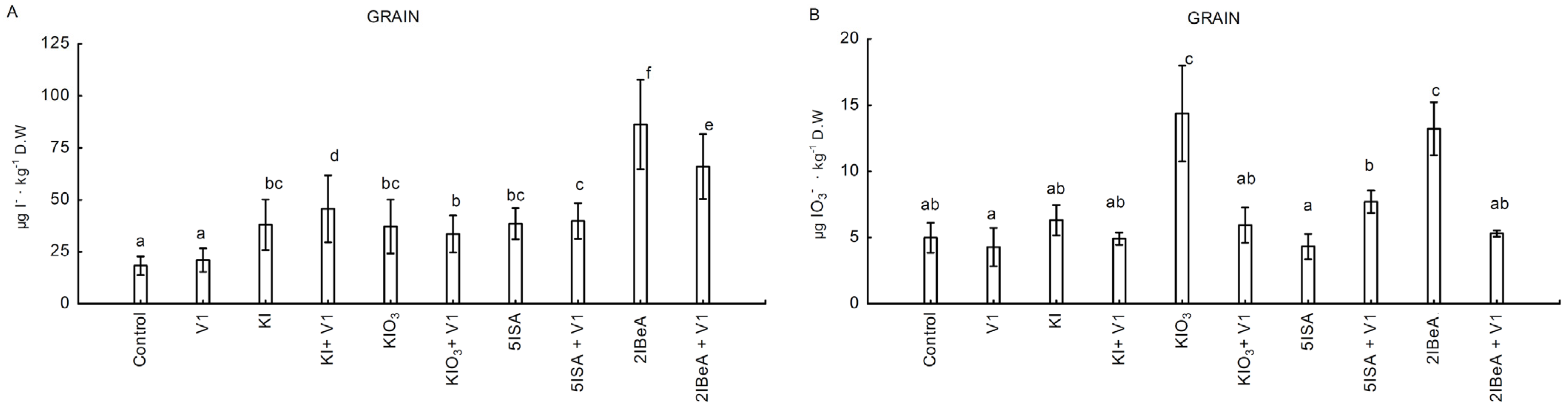
The combined application of 2IBeA with vanadium resulted in a statistically significant decrease in the accumulation of iodides and iodates compared to the individual application of 2IBeA. On the other hand, the combination of KI + V1 increased the accumulation of iodides in the grain of sweet corn compared to the solo application of KI. The combined application of KIO
3, 2IBeA, 5ISA with vanadium resulted in a significant increase in the synthesis/accumulation of 5ISA and 3,5-diISA iodosalicylates in grain. In the leaves, this relationship is consistent with the combined application of the vanadium from KI, KIO
3 and 5ISA. The combined application of vanadium with all iodine compounds, both organic and inorganic, significantly reduced the accumulation of 2,3,5-triIBeA in leaves and grains of sweet corn. The literature indicated that this organo-iodine compound is an auxin inhibitor [
52]. The results of the research based on a one-year experiment with a lower dose of vanadium (No. 3 year 2020) confirm the stimulating effect of vanadium (in a dose of 0.1 µM) on the accumulation of iodosalicylates (5ISA, 3,5-diISA) in the leaves and grains of sweet corn in combination with organic and inorganic iodine compounds. Research on tomato [
44] and lettuce [
20] also confirm the occurrence of iodosalicylates and iodobenzoates in control plants. After foliar application of iodosalicylates and iodobenzoates, their accumulation in sweet corn leaves and grain increased. Similar results were found in studies on tomato and lettuce [
20,
44].
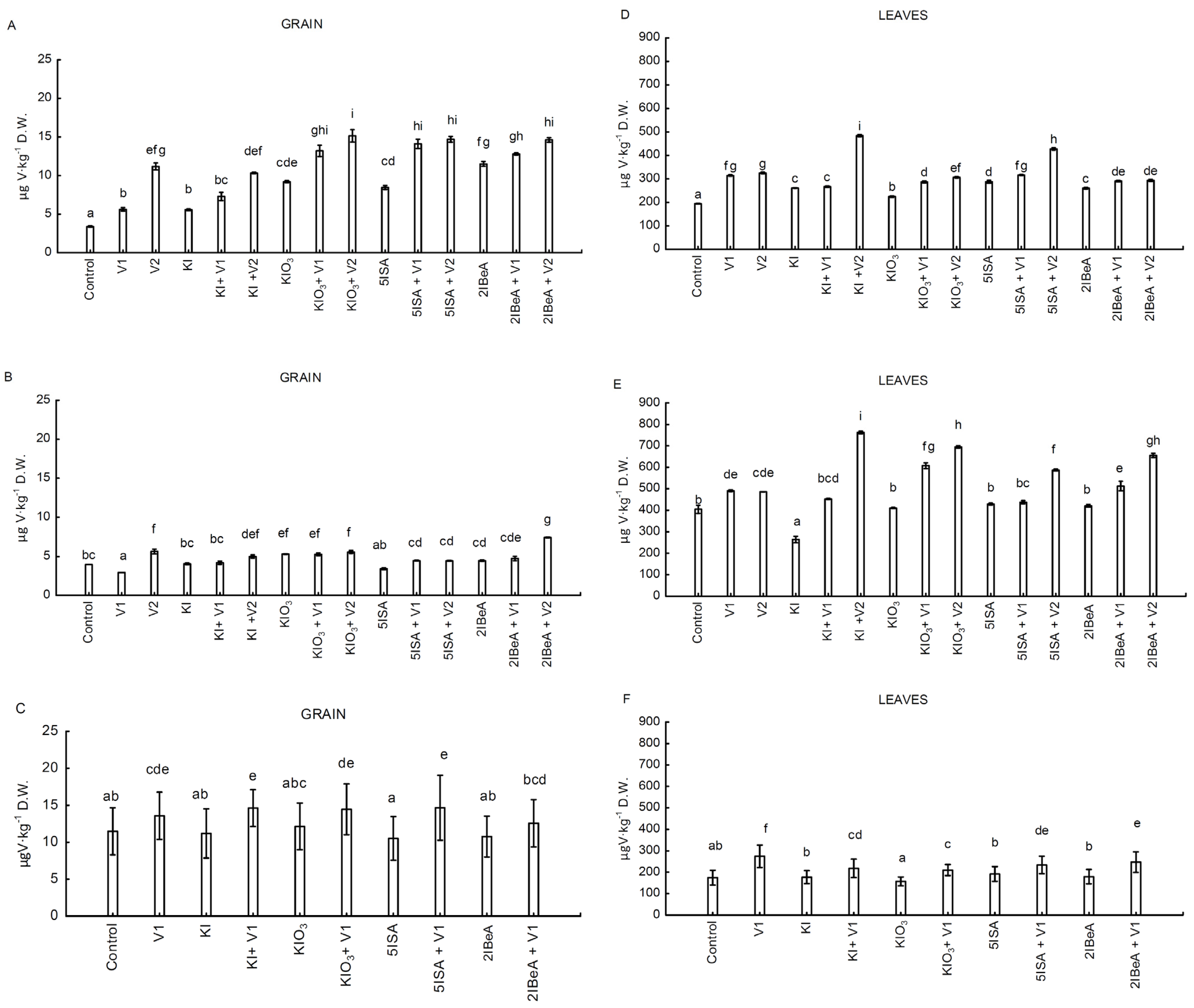
Transport of vanadium from the roots to the higher parts of plants, mainly to the generative organs, is limited. The highest degree of accumulation of this element is in the roots [
53,
54]. It is related to the process of vanadium biotransformation during the uptake of vanadium by the roots. This biotransformation consists in the reduction of pentavalent vanadium, which easily oxidize ketones, aldehydes, catechols, sulfhydryls and olefins placed in the cell wall even at pH 7. As a result, vanadium is retained by root tissues and vanadium (V) is reduced to the quadrivalent form of vanadium (IV) [
55].
In the conducted experiments No. 1 and No. 2 (with two applied doses of vanadium 0.1 and 1 µM), an increase in the applied dose of vanadium increased the accumulation of this element in leaves and corn grains. The effect of increased accumulation of vanadium with the increasing dose of this element was confirmed in the cultivation of pepper [
56], chickpea [
57] and beans [
54]. In experiments No. 1 and No. 2, the application of KI and 5ISA stimulated the accumulation of vanadium by increasing its content in the leaves compared to the application of only V2 (vanadium in a higher dose) without iodine. At the same time, V2 significantly reduced iodine accumulation in combination with 5ISA and KI. A similar effect was obtained in studies on sweet corn with the application of iodine and vanadium compounds to the soil [
43]. In the early development stages of sweet corn, the accumulation of iodine in the roots was highest in combination with the application of solo ammonium metavanadate at a dose of 0.1 µM [
33]. Vanadium is determined as a beneficial element to higher plants. The stimulating effect for plants ranges from 1–10 µg L
−1; higher doses have a phytotoxic effect on plants [
55,
58,
59]. Vanadium furtherance the nitrogen fixation process in soils with deficient in molybdenum, low doses stimulate the synthesis of chlorophyll [
59,
60]. In the carried-out experiments, vanadium doses of 0.1 µM and 1 µM did not cause any phytotoxic effect on plants. Vanadium did not affect the sweet corn yield in a statistically significant way.
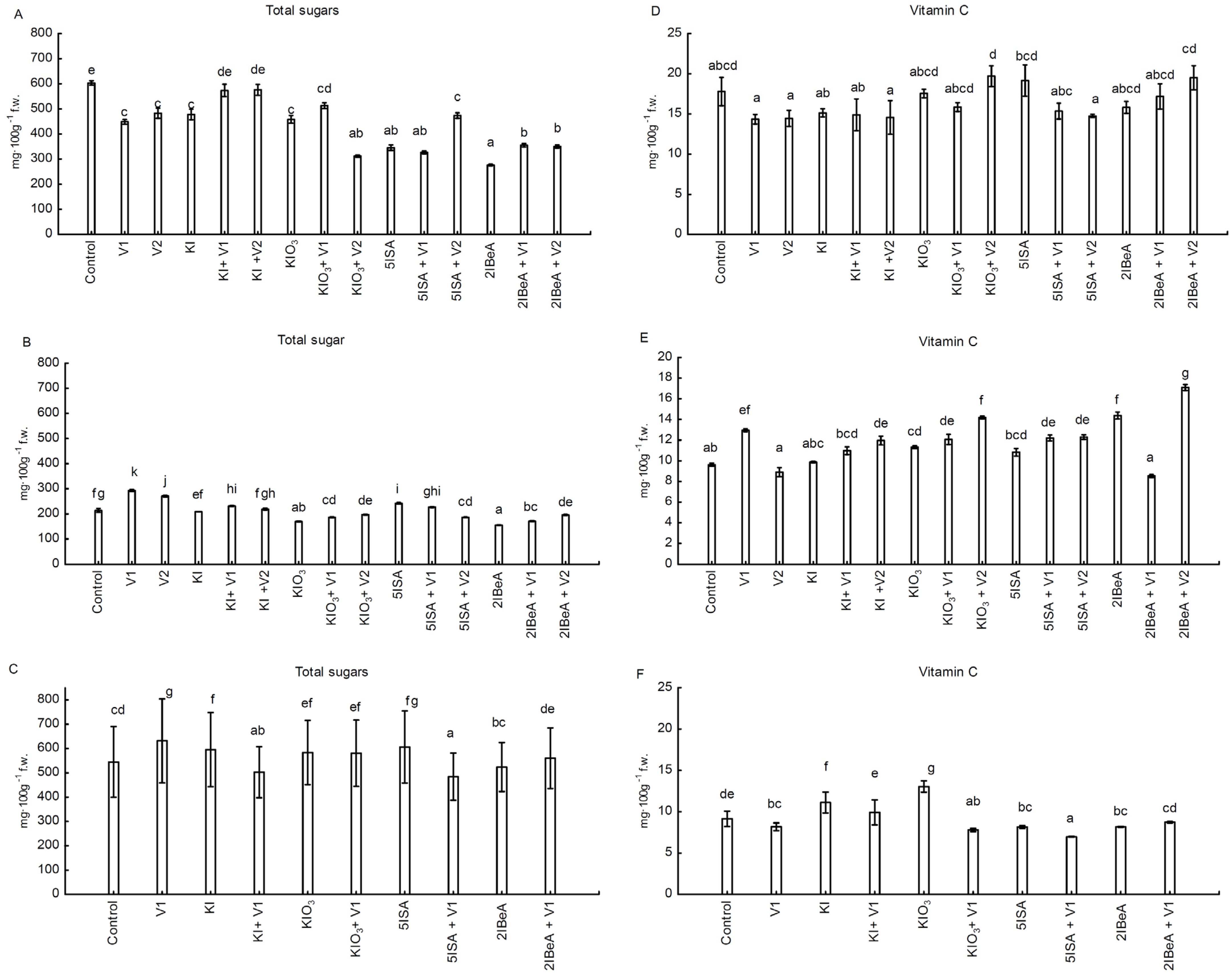
The content of total sugars and vitamin C was mainly formed by the weather conditions in the year of the experiment, and it was characterized by a huge diverseness between the studied combinations. In experiment No. 1 and No. 2, the grains had the lowest total sugar content after the foliar application of 2IBeA. The highest total sugars content was in the control for experiment No. 1, and for No. 2 and No. 3 with the application V1 (0.1 µM). In the experiments No. 1 and No. 2, the addition of vanadium to KIO
3 increased the content of vitamin C in the grains. In No. 3, the exclusive application of KIO
3 significantly increased the content of ascorbic acid. In the experience of lettuce the total sugar content and vitamin C was mainly determined by type of applied iodine compounds (organic or inorganic iodine), as well largely the dose determined the variability in sugar and vitamin C content. After the application of 5ISA in a dose of 8.0 μM was obtained the lowest level of vitamin C. The highest total sugar content was recorded in plants grown in a nutrient solution containing KIO
3 + SA. 5ISA in the dose of 40 μM resulted in a higher total sugar content in the lettuce compared to the lower doses (1.6 μM and 8 μM) [
39]. The application of KI and KIO
3 increased the content of vitamin C in Opuntia ficus-indica var. Copena V1 almost two times [
61] as well in water spinach [
41].
An important issue for the implementation of biofortified plants is regular monitoring of iodine intake status to detect excessive intakes. Some data emphasize that healthy adults who are iodine sufficient are curiously tolerant to iodine intake even 1000 µg·day
−1. Appropriate iodine intake is the most important for people in chronic iodine deficiency because swift increase of iodine may cause thyroiditis or hyperthyroidism [
10]. Upper levels of biofortification of the edible parts plants has to be an I of HQ < 1.0, which indicates a safe level for the consumer. In the presented study, this indicator HQ was not exceeded value 1.0 in all the conducted experiments. In the third experiment after the application of organic iodine compound 2IBeA and 2IBeA + V1, the I-RDA is the highest, so if the average daily intake per adult is 200 g, we can provide approximately 14% of the recommended daily iodine intake.








{kind=link}
{kind=link}
{kind=link}
{kind=link}