
Immunomodulatory Properties of Polysaccharide-Rich Young Green Barley (Hordeum vulgare) Extract and Its Structural Characterization

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
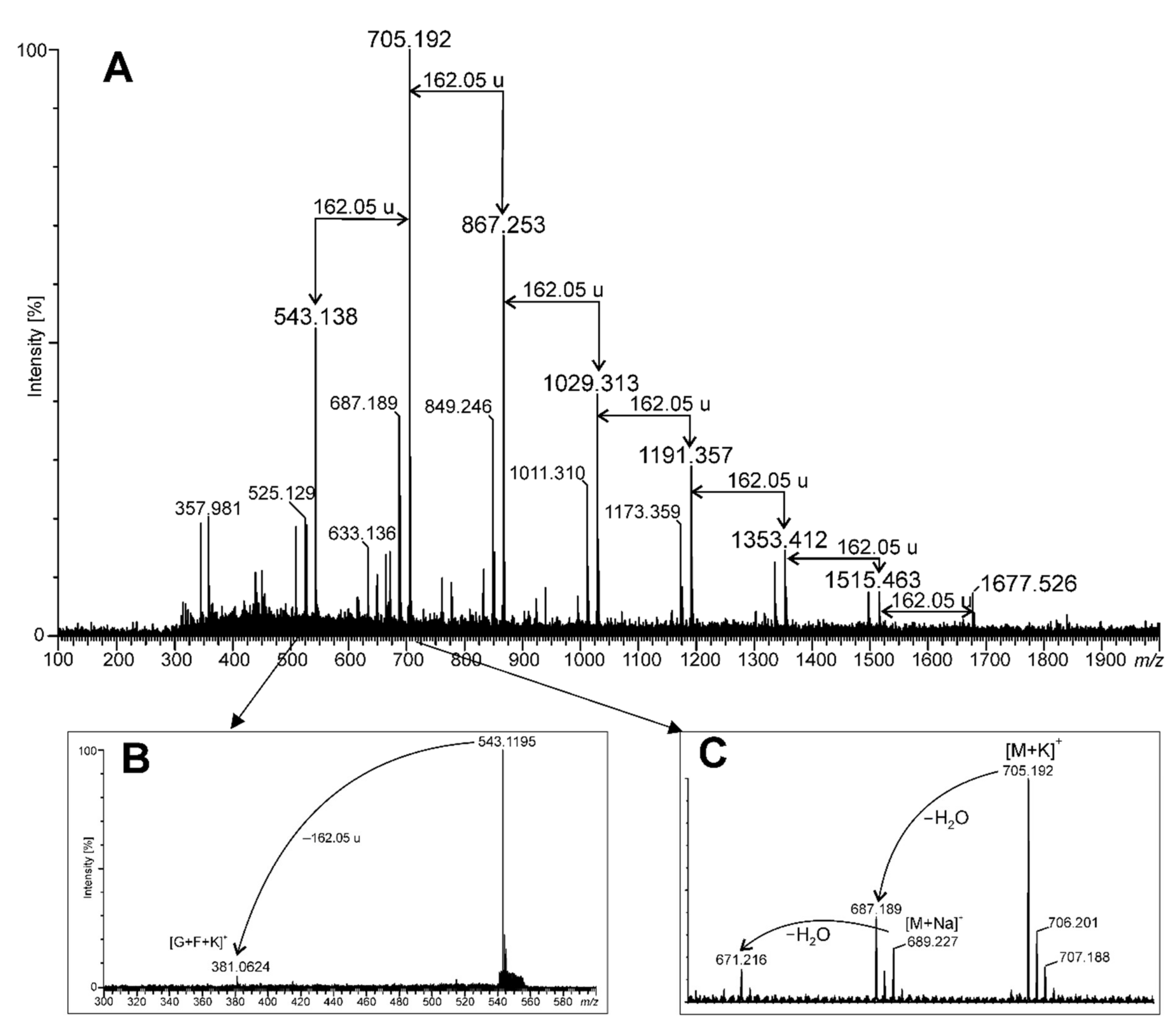
2.1. Young Green Barley Extract Composition
2.2. Young Green Barley Extract Impacts NK-92 Cell Viability and Proliferation
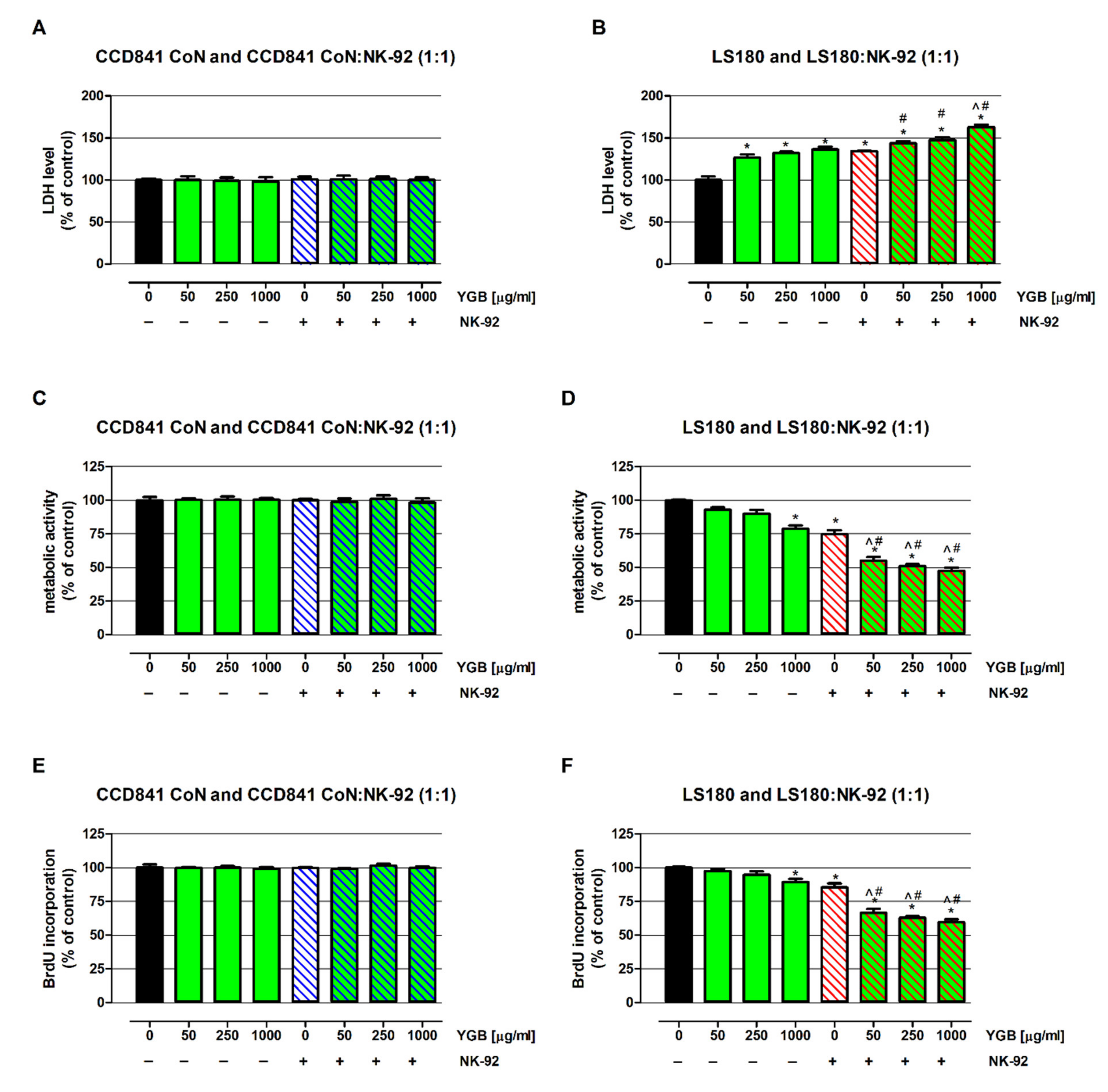
2.3. Young Green Barley Extract Enhances NK-92 Cells’ Cytotoxicity in Human Colon Cancer LS180 Cells without any Side Effects on Human Colon Epithelial CCD841 CoN Cells
2.4. Young Green Barley Extract Enhances Colon Cancer Cell Death in Response to NK-92 Cells
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Preparation of Young Green Barley Extract
4.3. Sugar Content Determination
4.4. Protein Content Determination
4.5. Proteins Separation—SDS-PAGE
4.6. Chemical Analyses of Sugars
4.7. Fructan Content Determination
4.8. FTIR Spectroscopy
4.9. GLC-MS Analysis
4.10. Mass Spectrometry
4.11. Cell Lines
4.12. Cell Treatment
4.13. Cytotoxicity Assessment—LDH Assay
4.14. Assessment of Metabolic Activity—MTT Assay
4.15. Assessment of DNA Synthesis—BrdU Assay
4.16. Cell Death Detection—ELISA
4.17. Cell Death Detection—Nuclear Double Staining
4.18. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Miller, J.S.; Lanier, L.L. Natural killer cells in cancer immunotherapy. Annu. Rev. Cancer Biol. 2019, 3, 77–103. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.S. Biology of natural killer cells in cancer and infection. Cancer Investig. 2002, 20, 405–419. [Google Scholar] [CrossRef] [PubMed]
- Kubota, K.; Matsuoka, Y.; Seki, H. Isolation of potent anti-inflammatory protein from barley leaves. Jpn. J. Inflamm. 1983, 3, 4. [Google Scholar]
- Ohtake, H.; Nonaka, S.; Sawada, Y.; Hagiwara, Y.; Hagiwara, H.; Kubota, K. Studies on the constituents of green juice from young barley leaves. Effect on dietarily induced hypercholesterolemia in rats. J. Pharm. Soc. Jpn. 1985, 105, 1052–1057. [Google Scholar] [CrossRef] [Green Version]
- Ohtake, H.; Yuasa, H.; Komura, C.; Miyauchi, T.; Hagiwara, Y.; Kubota, K. Studies on the constituents of green juice from young barley leaves. Antiulcer activity of fractions from barley juice. J. Pharm. Soc. Jpn. 1985, 105, 1046–1051. [Google Scholar] [CrossRef] [Green Version]
- Lahouar, L.; El-Bok, S.; Achour, L. Therapeutic potential of young green barley leaves in prevention and treatment of chronic diseases: An overview. Am. J. Chin. Med. 2015, 43, 1311–1329. [Google Scholar] [CrossRef]
- Zeng, Y.; Pu, X.; Yang, J.; Du, J.; Yang, X.; Li, X.; Li, L.; Zhou, Y.; Yang, T. Preventive and therapeutic role of functional ingredients of barley grass for chronic diseases in human beings. Oxid. Med. Cell. Longev. 2018, 2018, 3232080. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Pu, X.; Du, J.; Yang, X.; Li, X.; Mandal, M.S.N.; Yang, T.; Yang, J. Molecular mechanism of functional ingredients in barley to combat human chronic diseases. Oxid. Med. Cell. Longev. 2020, 2020, 3836172. [Google Scholar] [CrossRef]
- Kim, H.; Yu, K.W.; Hong, H.D.; Shin, K.S. Effect of arabinoxylan- and rhamnogalacturonan I-rich polysaccharides isolated from young barley leaf on intestinal immunostimulatory activity. J. Funct. Foods 2017, 35, 384–390. [Google Scholar] [CrossRef]
- Han, H.S.; Shin, J.S.; Song, Y.R.; Rhee, Y.K.; Cho, C.W.; Ryu, J.H.; Inn, K.S.; Hong, H.D.; Lee, K.T. Immunostimulatory effects of polysaccharides isolated from young barley leaves (Hordeum vulgare L.) with dual activation of Th1 and Th2 in splenic T cells and cyclophosphamide-induced immunosuppressed mice. Int. J. Biol. Macromol. 2020, 147, 954–964. [Google Scholar] [CrossRef]
- Han, L.; Meng, M.; Guo, M.; Cheng, D.; Shi, L.; Wang, X.; Wang, C. Immunomodulatory activity of a water-soluble polysaccharide obtained from highland barley on immunosuppressive mice models. Food Funct. 2019, 10, 304–314. [Google Scholar] [CrossRef] [PubMed]
- Ryu, D.S.; Kim, S.H.; Oh, S.M.; Lee, S.M.; Jeong, S.M.; Kim, S.H.; Lee, D.S. Immunosuppressive activities of water-soluble barley β-glucan on alloantigen reactive cell proliferation and cytotoxicity. Food Sci. Biotechnol. 2011, 20, 267–271. [Google Scholar] [CrossRef]
- Misra, C.K.; Das, B.K.; Mukherjee, S.C.; Pattnaik, P. Effect of multiple injections of β-glucan on non-specific immune response and disease resistance in Labeo rohita fingerlings. Fish Shellfish Immunol. 2006, 20, 305–319. [Google Scholar] [CrossRef] [PubMed]
- Aoe, S.; Yamanaka, C.; Fuwa, M.; Tamiya, T.; Nakayama, Y.; Miyoshi, T.; Kitazono, E. Effects of BARLEYmax and high-β-glucan barley line on short-chain fatty acids production and microbiota from the cecum to the distal colon in rats. PLoS ONE 2019, 14, e0218118. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, A.; Kitazono, E.; Imose, K.; Urita, S.; Matsui, T. Effect of functional barley BARLEYmax (Tantangara) on intestinal regulation: A double-blind, randomized, placebo-controlled parallel group comparison clinical study. Jpn. Pharmacol. Ther. 2017, 45, 1047–1055. [Google Scholar]
- Sasaki, D.; Sasaki, K.; Kadowaki, Y.; Aotsuka, Y.; Kondo, A. Bifidogenic and butyrogenic effects of young barley leaf extract in an in vitro human colonic microbiota model. AMB Express 2019, 9, 182. [Google Scholar] [CrossRef]
- Liu, C.; Li, X.; Li, Y.; Feng, Y.; Zhou, S.; Wang, F. Structural characterization and antimutagenic activity of a novel polysaccharide isolated from Sepiella maindroni ink. Food Chem. 2008, 110, 807–813. [Google Scholar] [CrossRef]
- Grube, M.; Bekers, M.; Upite, D.; Kaminska, E. Infrared spectra of some fructans. Spectroscopy 2002, 16, 289–296. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Pei, F.; Shi, Y.; Zhao, L.; Fang, Y.; Hu, Q. Purification, characterization and anti-proliferation activity of polysaccharides from Flammulina velutipes. Carbohydr. Polym. 2012, 88, 474–480. [Google Scholar] [CrossRef]
- Gieroba, B.; Krysa, M.; Wojtowicz, K.; Wiater, A.; Pleszczyńska, M.; Tomczyk, M.; Sroka-Bartnicka, A. The FT-IR and Raman spectroscopies as tools for biofilm characterization created by cariogenic Streptococci. Int. J. Mol. Sci. 2020, 21, 3811. [Google Scholar] [CrossRef]
- Galinari, É.; Sabry, D.A.; Sassaki, G.L.; Macedo, G.R.; Passos, F.; Mantovani, H.C.; Rocha, H. Chemical structure, antiproliferative and antioxidant activities of a cell wall α-d-mannan from yeast Kluyveromyces marxianus. Carbohydr. Polym. 2017, 157, 1298–1305. [Google Scholar] [CrossRef] [PubMed]
- Seymour, F.R.; Julian, R.L.; Jeanes, A.; Lamberts, B.L. Structural analysis of insoluble d-glucans by Fourier-transform, infrared difference-spectrometry: Correlation between structures of dextrans from strains of leuconostoc mesenteroides and of d-glucans from strains of streptococcus mutans. Carbohydr. Res. 1980, 86, 227–246. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Z.; Zhou, L.; Bao, J.; Xu, J. The modifications of a fructan from Anemarrhena asphodeloides Bunge and their antioxidant activities. Int. J. Biol. Macromol. 2020, 164, 4435–4443. [Google Scholar] [CrossRef] [PubMed]
- Guerrant, G.O.; Moss, C.W. Determination of monosaccharides as aldononitrile, O-methyloxime, alditol, and cyclitol acetate derivatives by gas chromatography. Anal. Chem. 1984, 56, 633–638. [Google Scholar] [CrossRef]
- Wahjudi, P.N.; Patterson, M.E.; Lim, S.; Yee, J.K.; Mao, C.S.; Lee, W.N.P. Measurement of glucose and fructose in clinical samples using gas chromatography/mass spectrometry. Clin. Biochem. 2010, 43, 198–207. [Google Scholar] [CrossRef] [Green Version]
- Müller, J.; Aeschbacher, R.A.; Sprenger, N.; Boller, T.; Wiemken, A. Disaccharide-mediated regulation of sucrose: Fructan-6-fructosyltransferase, a key enzyme of fructan synthesis in barley leaves. Plant Physiol. 2000, 123, 265–274. [Google Scholar] [CrossRef] [Green Version]
- Carpita, N.C.; Kanabus, J.; Housley, T.L. Linkage structure of fructans and fructan oligomers from Triticum aestivum and Festuca arundinacea leaves. J. Plant. Physiol. 1989, 134, 162–168. [Google Scholar] [CrossRef]
- Vijn, I.; Smeekens, S. Fructan: More than a reserve carbohydrate? Plant Physiol. 1999, 120, 351–360. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, C.; Andersson, A.A.M.; Andersson, R.; Mangelsen, E.; Sun, C.; Åman, P. Relationship of grain fructan content to degree of polymerisation in different barleys. Food Nutr. Sci. 2014, 5, 581–589. [Google Scholar] [CrossRef] [Green Version]
- Peukert, M.; Thiel, J.; Peshev, D.; Weschke, W.; van den Ende, W.; Mock, H.P.; Matros, A. Spatio-temporal dynamics of fructan metabolism in developing barley grains. Plant Cell. 2014, 26, 3728–3744. [Google Scholar] [CrossRef] [Green Version]
- Franco-Robles, E.; López, M.G. Implication of fructans in health: Immunomodulatory and antioxidant mechanisms. Sci. World J. 2015, 2015, 289267. [Google Scholar] [CrossRef] [Green Version]
- Young, I.D.; Latousakis, D.; Juge, N. The immunomodulatory properties of β-2,6 fructans: A comprehensive review. Nutrients 2021, 13, 1309. [Google Scholar] [CrossRef]
- Meng, X.; Liang, H.; Luo, L. Antitumor polysaccharides from mushrooms: A review on the structural characteristics, antitumor mechanisms and immunomodulating activities. Carbohydr. Res. 2016, 424, 30–41. [Google Scholar] [CrossRef]
- Arai, S.; Meagher, R.; Swearingen, M.; Myint, H.; Rich, E.; Martinson, J.; Klingemann, H. Infusion of the allogeneic cell line NK-92 in patients with advanced renal cell cancer or melanoma: A phase I trial. Cytotherapy 2008, 10, 625–632. [Google Scholar] [CrossRef]
- Cheng, M.; Zhang, J.; Jiang, W.; Chen, Y.; Tian, Z. Natural killer cell lines in tumor immunotherapy. Front. Med. 2012, 6, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.H.; Maki, G.; Klingemann, H.G. Characterization of a human cell line (NK-92) with phenotypical and functional characteristics of activated natural killer cells. Leukemia 1994, 8, 652–658. [Google Scholar] [PubMed]
- Kawka, K.; Lemieszek, M.K.; Rzeski, W. Chemopreventive properties of young green barley extracts in in vitro model of colon cancer. Ann. Agric. Environ. Med. 2019, 26, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Lemieszek, M.K.; Rzeski, W. Enhancement of chemopreventive properties of young green barley and chlorella extracts used together against colon cancer cells. Ann. Agric. Environ. Med. 2020, 27, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Sawardeker, J.S.; Sloneker, J.H.; Jeanes, A. Quantitative determination of monosaccharides as their alditol acetates by gas liquid chromatography. Anal. Chem. 1965, 37, 1602–1604. [Google Scholar] [CrossRef]
- Ciucanu, J.; Kerek, F. A simple and rapid method for the permethylation of carbohydrates. Carbohydr. Res. 1984, 131, 209–217. [Google Scholar] [CrossRef]
- Choma, A.; Komaniecka, I. Characterization of cyclic β-glucans of Bradyrhizobium by MALDI-TOF mass spectrometry. Carbohydr. Res. 2011, 346, 1945–1950. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Amount (%) |
|---|---|
| t-Fruf-(2→ | 22.3 |
| t-Manp-(1→ | 1.2 |
| t-Glcp-(1→ | 47.2 |
| t-Galp-(1→ | 5 |
| →6)-Fruf-(2→ | 9.7 |
| →3)-Glcp-(1→ | 0.9 |
| →4)-Glcp-(1→ | 3.1 |
| →3)-Galp-(1→ | 0.5 |
| →6)-Glcp-(1→ | 3 |
| →6)-Galp-(1→ | 2 |
| →1,6)-Fruf-(2→ | 3.5 |
| →3,6)-Glcp-(1→ | 1.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lemieszek, M.K.; Komaniecka, I.; Chojnacki, M.; Choma, A.; Rzeski, W. Immunomodulatory Properties of Polysaccharide-Rich Young Green Barley (Hordeum vulgare) Extract and Its Structural Characterization. Molecules 2022, 27, 1742. https://doi.org/10.3390/molecules27051742
Lemieszek MK, Komaniecka I, Chojnacki M, Choma A, Rzeski W. Immunomodulatory Properties of Polysaccharide-Rich Young Green Barley (Hordeum vulgare) Extract and Its Structural Characterization. Molecules. 2022; 27(5):1742. https://doi.org/10.3390/molecules27051742
Chicago/Turabian StyleLemieszek, Marta Kinga, Iwona Komaniecka, Michał Chojnacki, Adam Choma, and Wojciech Rzeski. 2022. "Immunomodulatory Properties of Polysaccharide-Rich Young Green Barley (Hordeum vulgare) Extract and Its Structural Characterization" Molecules 27, no. 5: 1742. https://doi.org/10.3390/molecules27051742