
Identification and Functional Characterization of Tyrosine Decarboxylase from Rehmannia glutinosa

Abstract
:1. Introduction

2. Results
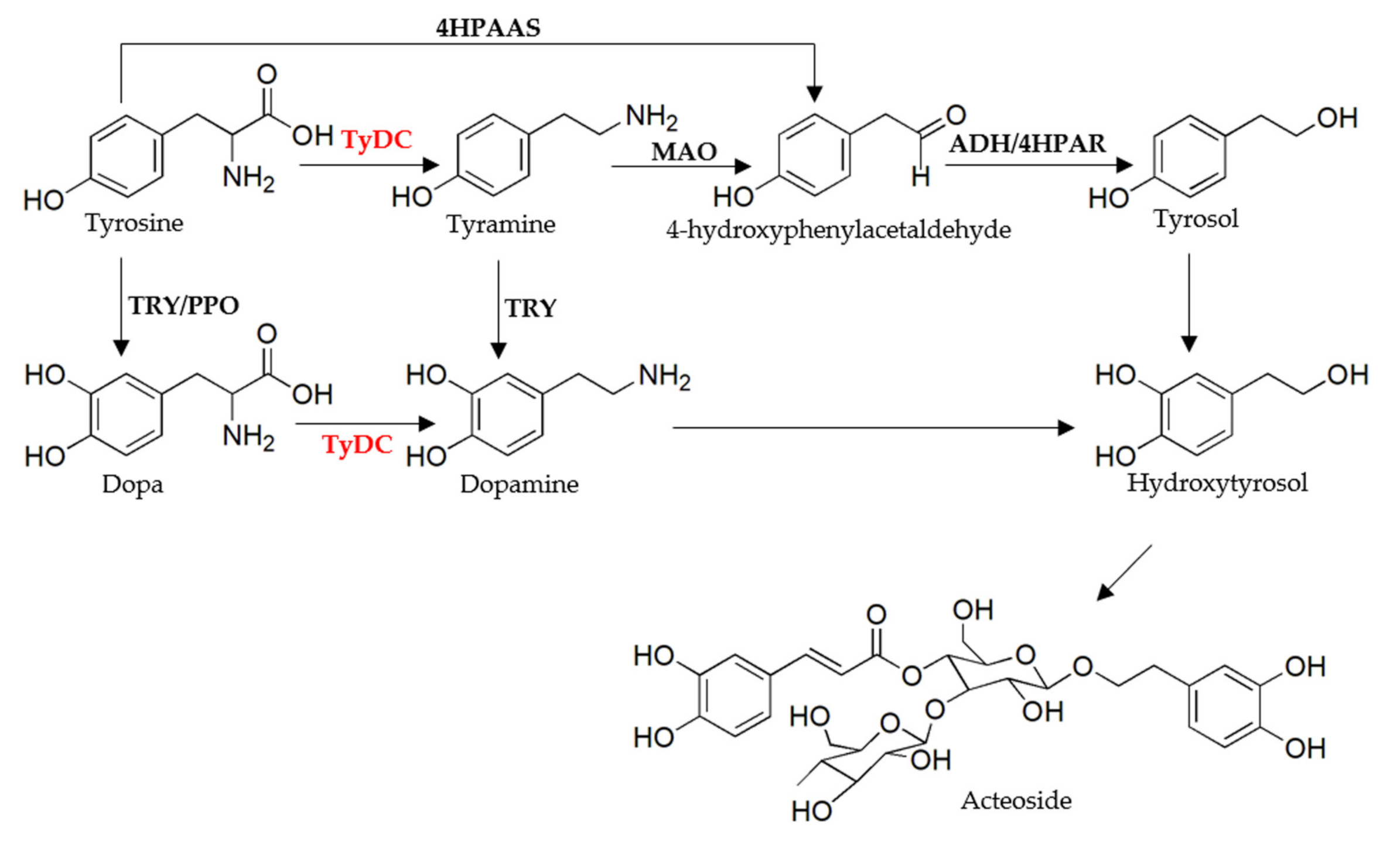
2.1. The Identification of Candidate Genes in the Biosynthesis of Acteoside
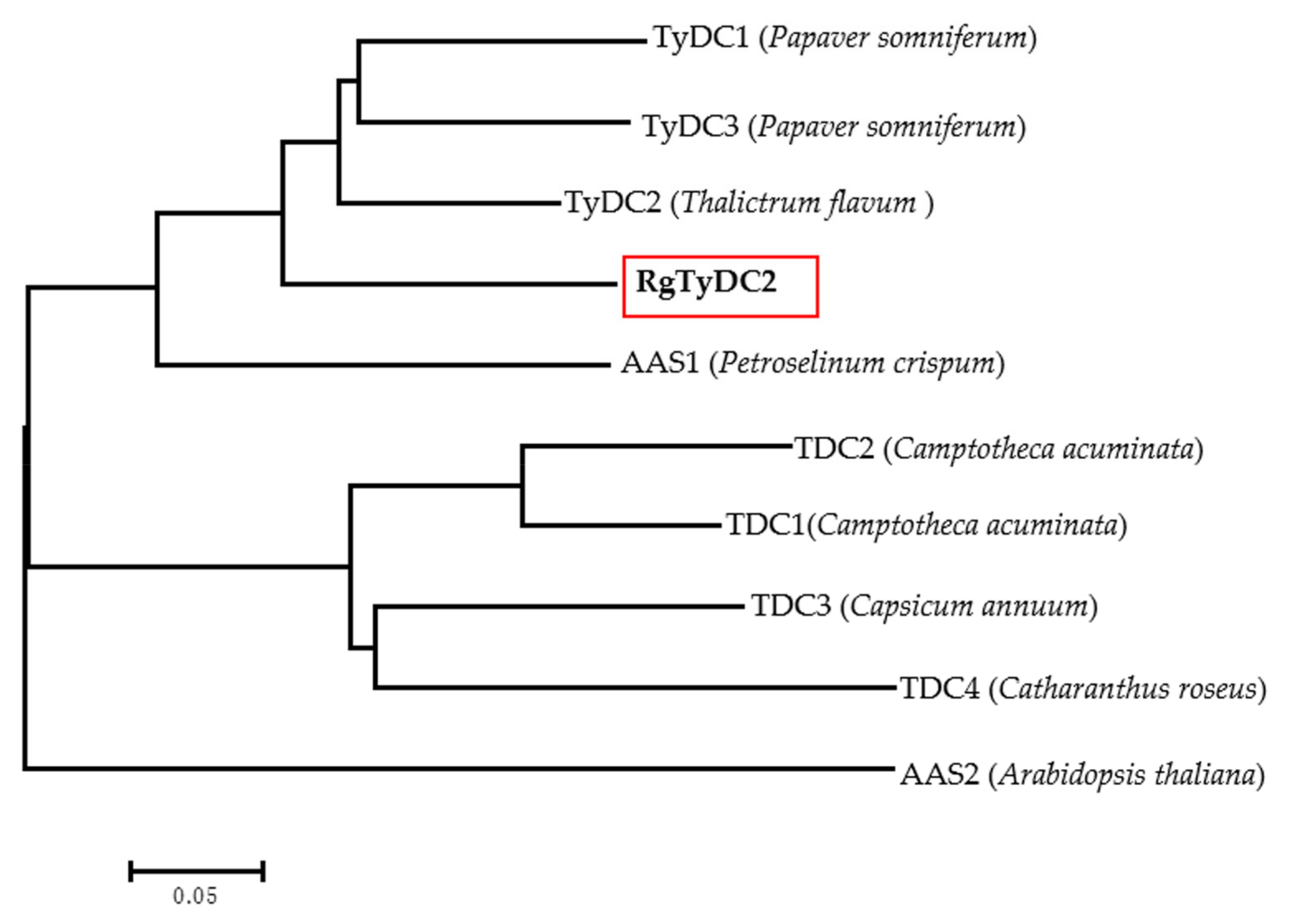
2.2. Identification and Sequence Analysis of R. glutinosa TyDC Genes
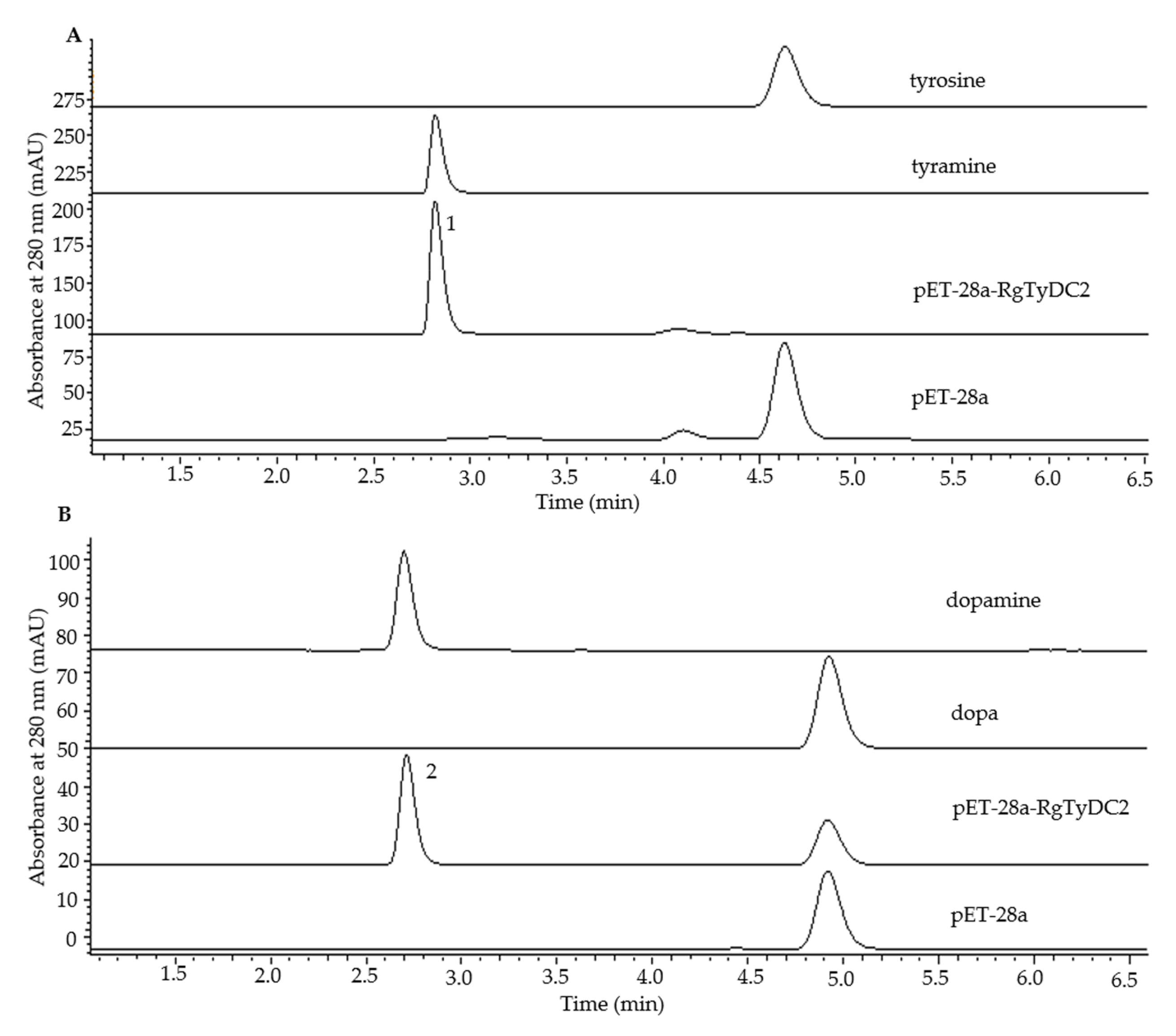
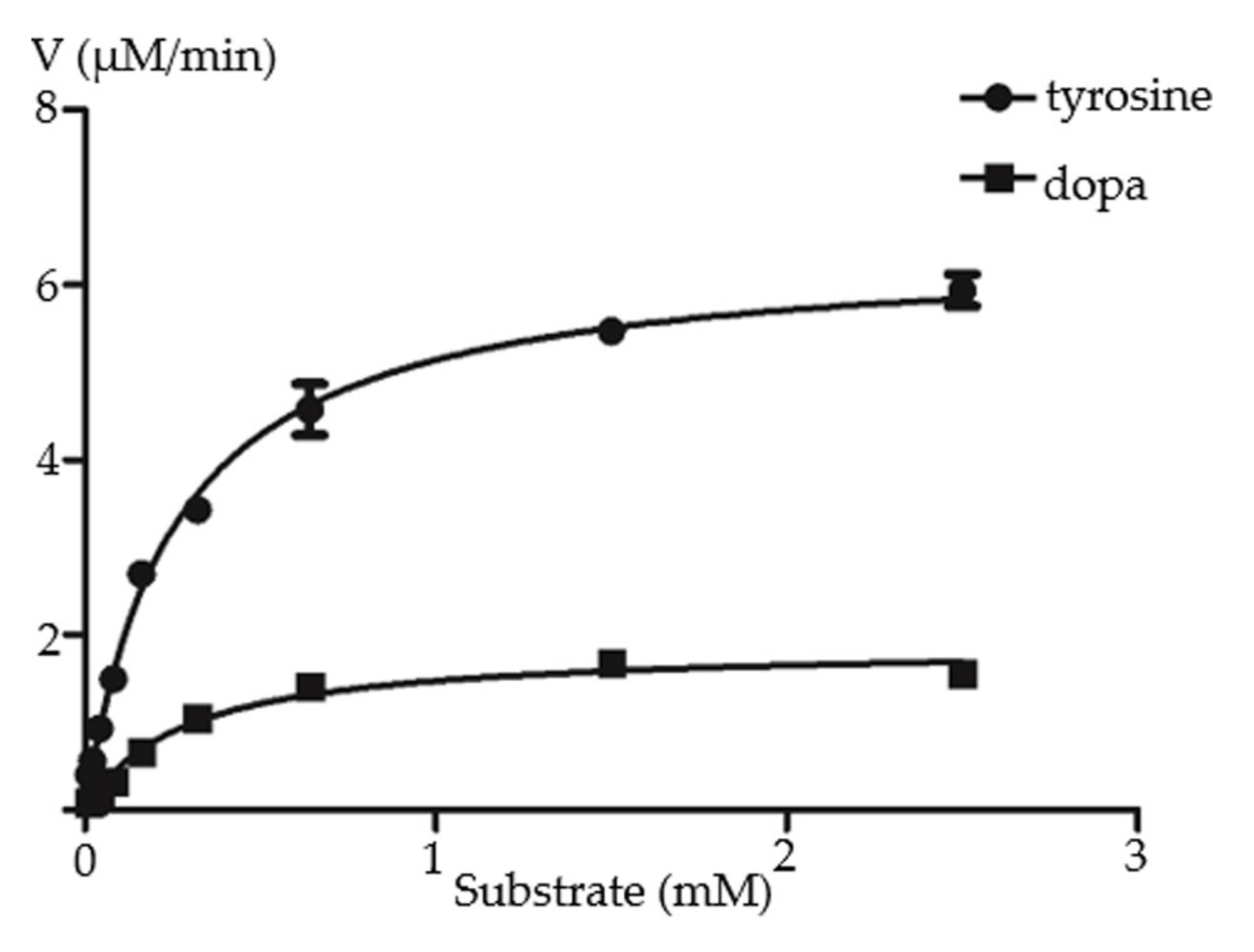
2.3. Functional Characterization of RgTyDC2


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | kcat (s−1) | Km (μM) | kcat/Km (s−1M−1) |
|---|---|---|---|
| tyrosine | 0.507 ± 0.0104 | 249.7 ± 17.11 | 2.032 × 103 |
| dopa | 0.126 ± 0.0052 | 273.8 ± 37.18 | 4.602 × 102 |

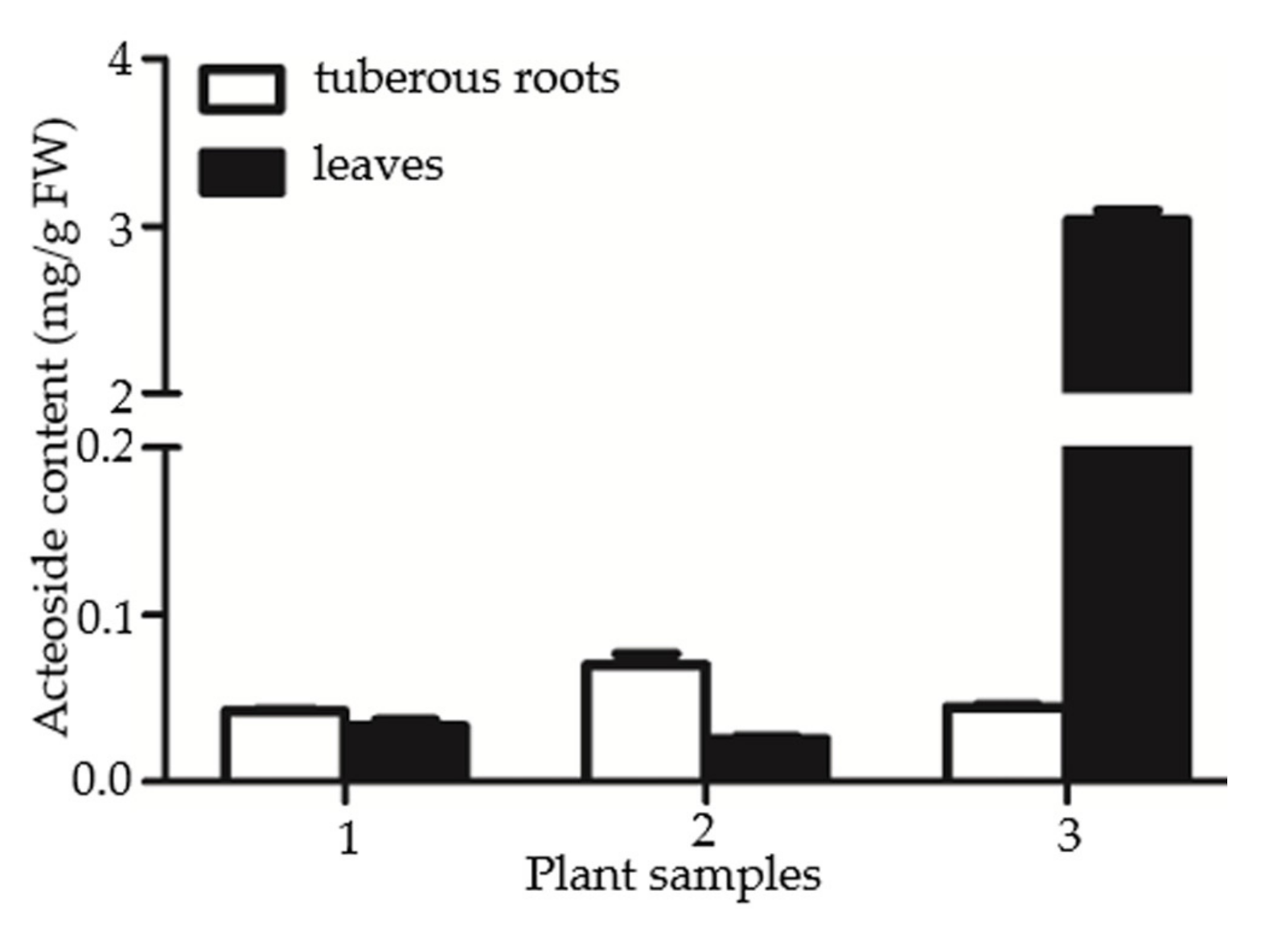
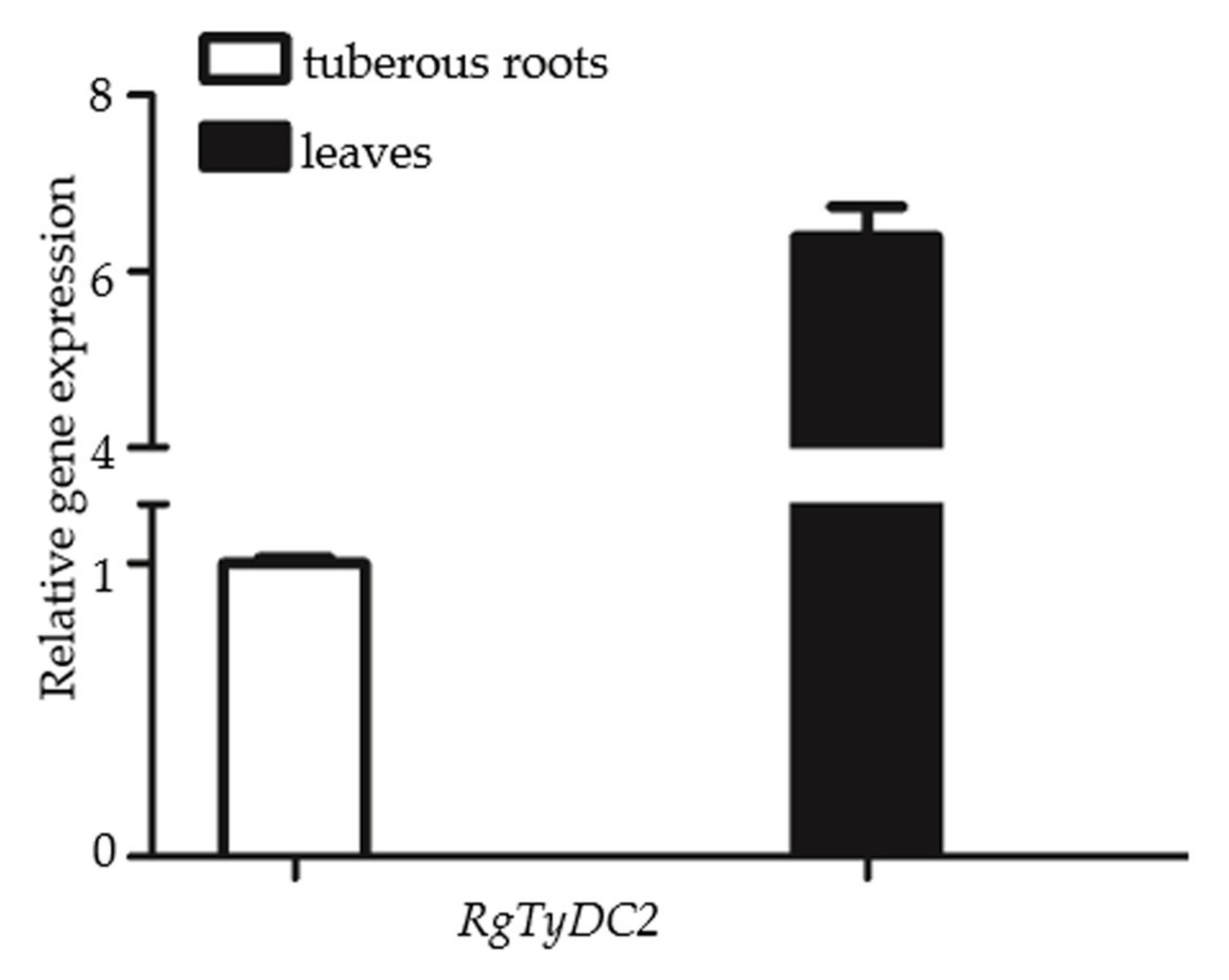
2.4. Gene Expression Data of RgTyDC2 Was Consistent with the Accumulation Pattern of Acteoside in R. glutinosa
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Chemicals
4.2. Identification and Cloning of the Tyrosine Decarboxylase from R. glutinosa
4.3. Heterologous Expression of RgTyDC2
4.4. Enzyme Assays
4.5. Determination of Acteoside from Plant Materials
4.6. HPLC Analysis
4.7. Real-Time PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, R.X.; Li, M.X.; Jia, Z.P. Rehmannia glutinosa: Review of botany, chemistry and pharmacology. J. Ethnopharmacol. 2008, 117, 199–214. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.L.; Cao, Y.G.; Kan, Y.X.; Ren, Y.J.; Wang, M.N.; Fan, X.L.; Zheng, X.K.; Feng, W.S. Renoprotective activity of a new amide and a new hydroxycinnamic acid derivative from the fresh roots of Rehmannia glutinosa. J. Asian Nat. Prod. Res. 2022, 24, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.Y.; Fu, P.; Fan, W.X.; Zuo, C.; Feng, P.; Shi, P.; Cao, L.N.; Liu, F.; Zhou, L.; Chen, F.; et al. Treatment of primary chronic glomerulonephritis with Rehmannia glutinosa acteosides in combination with the angiotensin receptor blocker irbesartan: A randomized controlled trial. Phytother. Res. 2014, 28, 132–136. [Google Scholar] [CrossRef]
- Qin, Z.X.; Wang, W.; Liao, D.Q.; Wu, X.Y.; Li, X.E. UPLC-Q/TOF-MS-Based serum metabolomics reveals hypoglycemic effects of Rehmannia glutinosa, Coptis chinensis and their combination on high-fat-diet-induced diabetes in KK-Ay Mice. Int. J. Mol. Sci. 2018, 19, 3984. [Google Scholar] [CrossRef] [Green Version]
- Bai, L.; Shi, G.Y.; Yang, Y.J.; Chen, W.; Zhang, L.F.; Qin, C. Rehmannia glutinosa exhibits anti-aging effect through maintaining the quiescence and decreasing the senescence of hematopoietic stem cells. Anim. Model. Exp. Med. 2018, 1, 194–202. [Google Scholar] [CrossRef]
- Xu, L.; Kwak, M.; Zhang, W.; Zeng, L.; Whan Lee, P.C.; Jin, J.O. Rehmannia glutinosa polysaccharide induces toll-like receptor 4 dependent spleen dendritic cell maturation and anti-cancer immunity. Oncoimmunology 2017, 6, e1325981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.M.; Pei, L.X.; Zhang, Y.Y.; Cheng, Y.X.; Niu, C.L.; Cui, Y.; Feng, W.S.; Wang, G.F. Ethanol extract of Rehmannia glutinosa exerts antidepressant-like effects on a rat chronic unpredictable mild stress model by involving monoamines and BDNF. Metab. Brain Dis. 2018, 33, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Zhang, H.; Bi, J.; Zhang, X.L. Neuroprotective activities of catalpol on MPP+/MPTP-induced neurotoxicity. Neurol. Res. 2008, 30, 639–644. [Google Scholar] [CrossRef]
- He, J.; Hu, X.P.; Zeng, Y.; Li, Y.; Wu, H.Q.; Qiu, R.Z.; Ma, W.J.; Li, T.; Li, C.Y.; He, Z.D. Advanced research on acteoside for chemistry and bioactivities. J. Asian Nat. Prod. Res. 2011, 13, 449–464. [Google Scholar] [CrossRef]
- Hausmann, M.; Obermeier, F.; Paper, D.H.; Balan, K.; Dunger, N.; Menzel, K.; Falk, W.; Schoelmerich, J.; Herfarth, H.; Rogler, G. In vivo treatment with the herbal phenylethanoid acteoside ameliorates intestinal inflammation in dextran sulphate sodium-induced colitis. Clin. Exp. Immunol. 2007, 148, 373–381. [Google Scholar] [CrossRef]
- Wang, H.Q.; Xu, Y.X.; Yan, J.; Zhao, X.Y.; Sun, X.B.; Zhang, Y.P.; Guo, J.C.; Zhu, C.Q. Acteoside protects human neuroblastoma SH-SY5Y cells against β-amyloid-induced cell injury. Brain Res. 2009, 1283, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ma, C.; Wang, S. Effects of acteoside on lipopolysaccharide-induced inflammation in acute lung injury via regulation of NF-κB pathway in vivo and in vitro. Toxicol. Appl. Pharmacol. 2015, 285, 128–135. [Google Scholar]
- Wu, Y.Y.; Zeng, M.N.; Xu, R.Q.; Zhang, B.B.; Wang, S.C.; Li, B.K.; Kan, Y.X.; Cao, B.; Zheng, X.K.; Feng, W.S. Inhibitory activity of acteoside in melanoma via regulation of the ERβ-Ras/Raf1-STAT3 pathway. Arch. Biochem. Biophys. 2021, 710, 108978. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Wang, J.; Liu, L.; Chen, M.Q.; Wang, Z.Y. Acteoside as a potential therapeutic option for primary hepatocellular carcinoma: A preclinical study. BMC Cancer 2020, 20, 936. [Google Scholar] [CrossRef]
- Alipieva, K.; Korkina, L.; Orhan, I.E.; Georgiev, M.I. Verbascoside-A review of its occurrence, (bio)synthesis and pharmacological significance. Biotechnol. Adv. 2014, 32, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Ellis, B.E. Production of hydroxyphenylethanol glycosides in suspension cultures of Syringa vulgaris. Phytochemistry 1983, 22, 1941–1943. [Google Scholar] [CrossRef]
- Saimaru, H.; Orihara, Y. Biosynthesis of acteoside in cultured cells of Olea europaea. J. Nat. Med. 2010, 64, 139–145. [Google Scholar] [CrossRef]
- Torrens-Spence, M.P.; Pluskal, T.; Li, F.S.; Carballo, V.; Weng, J.K. Complete pathway elucidation and heterologous reconstitution of Rhodiola salidroside biosynthesis. Mol. Plant 2017, 11, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.Q.; Zhi, J.Y.; Zhang, Z.Y.; Wang, L.N.; Suo, Y.F.; Xie, C.X.; Li, M.J.; Zhang, B.; Du, J.F.; Gu, L.; et al. Transcriptome analysis of salicylic acid treatment in Rehmannia glutinosa hairy roots using RNA-seq technique for identification of genes involved in acteoside biosynthesis. Front. Plant Sci. 2017, 8, 787. [Google Scholar] [CrossRef]
- Facchini, P.J.; Huber-Allanach, K.L.; Tari, L.W. Plant aromatic L-amino acid decarboxylases: Evolution, biochemistry, regulation, and metabolic engineering applications. Phytochemistry 2000, 54, 121–138. [Google Scholar] [CrossRef]
- Wang, F.Q.; Zhi, J.Y.; Xie, C.X.; Du, J.F.; Suo, Y.F.; Wang, H.Y.; Zhang, Z.Y. Cloning and expression analysis of a tyrosine decarboxylase gene from Rehmannia glutinosa. Zhongguo Zhong Yao Za Zhi 2016, 41, 2981–2986. [Google Scholar] [PubMed]
- Torrens-Spence, M.P.; Liu, P.Y.; Ding, H.Z.; Harich, K.; Gillaspy, G.; Li, J.Y. Biochemical evaluation of the decarboxylation and decarboxylation-deamination activities of plant aromatic amino acid decarboxylases. J. Biol. Chem. 2013, 288, 2376–2387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torrens-Spence, M.P.; Lazear, M.; von Guggenberg, R.; Ding, H.Z.; Li, J.Y. Investigation of a substrate-specifying residue within Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases. Phytochemistry 2014, 106, 37–43. [Google Scholar] [CrossRef] [PubMed]
- De Masi, L.; Castaldo, D.; Pignone, D.; Servillo, L.; Facchiano, A. Experimental evidence and in silico identification of tryptophan decarboxylase in Citrus genus. Molecules 2017, 22, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Facchiano, A.; Pignone, D.; Servillo, L.; Castaldo, D.; De Masi, L. Structure and ligands interactions of Citrus tryptophan decarboxylase by molecular modeling and docking simulations. Biomolecules 2019, 9, 117. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.Q.; Wang, X.N.; Wang, W.S.; Duan, H.Y. De novo transcriptome sequencing-based discovery and expression analyses of verbascoside biosynthesis-associated genes in Rehmannia glutinosa tuberous roots. Mol. Breed. 2016, 36, 139. [Google Scholar] [CrossRef]
- Hu, J.W.; Li, W.; Liu, Z.; Zhang, G.L.; Luo, Y.G. Molecular cloning and functional characterization of tyrosine decarboxylases from galanthamine-producing Lycoris radiata. Acta Physiol. Plant. 2021, 43, 84. [Google Scholar] [CrossRef]
- Zhang, J.X.; Ma, L.Q.; Yu, H.S.; Zhang, H.; Wang, H.T.; Qin, Y.F.; Shi, G.L.; Wang, Y.N. A tyrosine decarboxylase catalyzes the initial reaction of the salidroside biosynthesis pathway in Rhodiola sachalinensis. Plant Cell Rep. 2011, 30, 1443–1453. [Google Scholar] [CrossRef]
- Sun, P.; Chen, Q.J.; Li, X.E. Selection of the reference genes for gene expression studies in Rehmannia glutinosa by real-time quantitative PCR. Chin. Agric. Sci. Bull. 2011, 27, 76–82. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Wang, Y.; Huang, L.; Chen, C.; An, N.; Zheng, X. Identification and Functional Characterization of Tyrosine Decarboxylase from Rehmannia glutinosa. Molecules 2022, 27, 1634. https://doi.org/10.3390/molecules27051634
Li Y, Wang Y, Huang L, Chen C, An N, Zheng X. Identification and Functional Characterization of Tyrosine Decarboxylase from Rehmannia glutinosa. Molecules. 2022; 27(5):1634. https://doi.org/10.3390/molecules27051634
Chicago/Turabian StyleLi, Yuanjun, Yanan Wang, Longyu Huang, Chunrong Chen, Na An, and Xiaoke Zheng. 2022. "Identification and Functional Characterization of Tyrosine Decarboxylase from Rehmannia glutinosa" Molecules 27, no. 5: 1634. https://doi.org/10.3390/molecules27051634