
Natural Dietary Compound Xanthohumol Regulates the Gut Microbiota and Its Metabolic Profile in a Mouse Model of Alzheimer’s Disease

 , and
, and
Abstract
:1. Introduction
2. Results
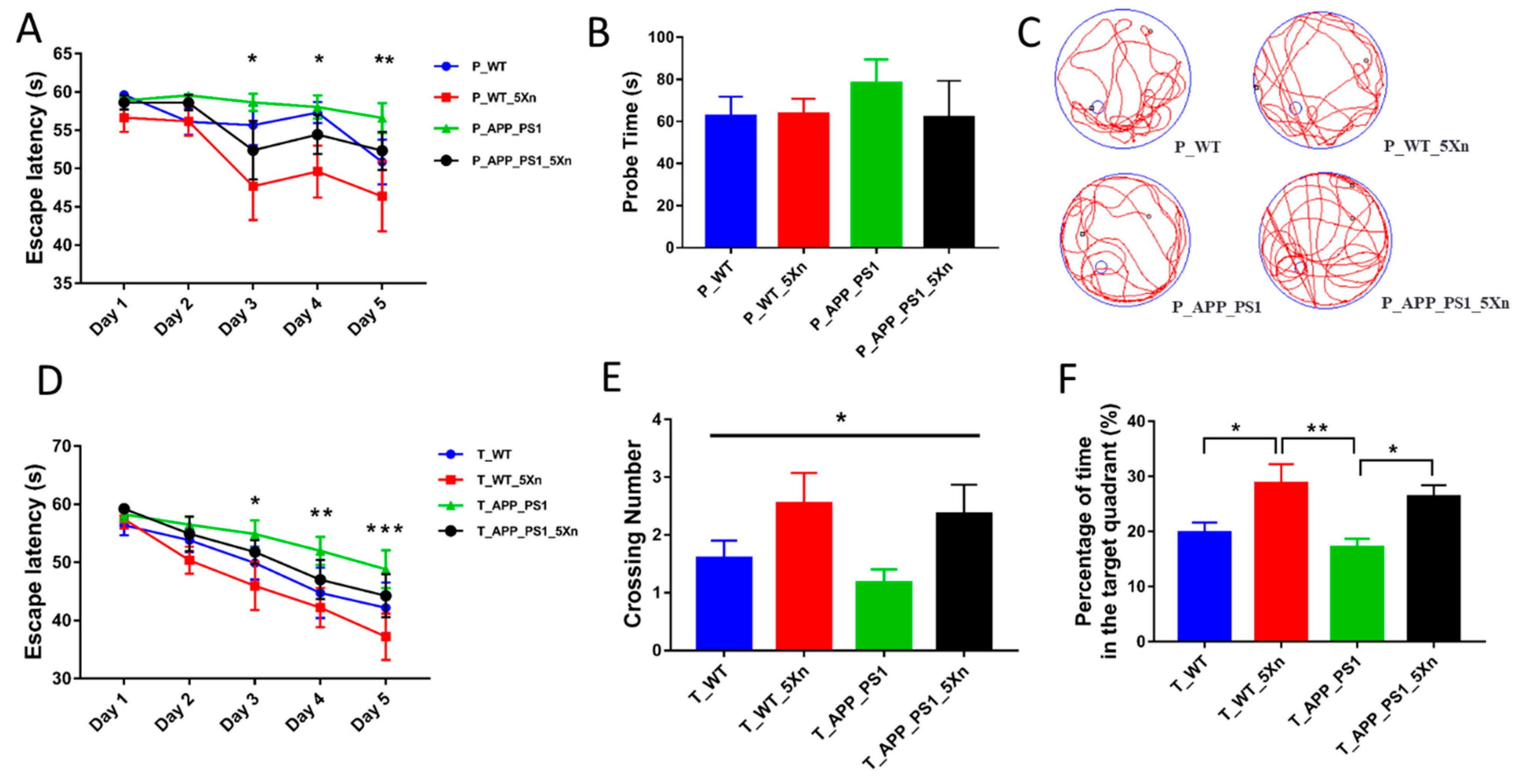
2.1. Xn Improves Cognitive Function in APP/PS1 Mice
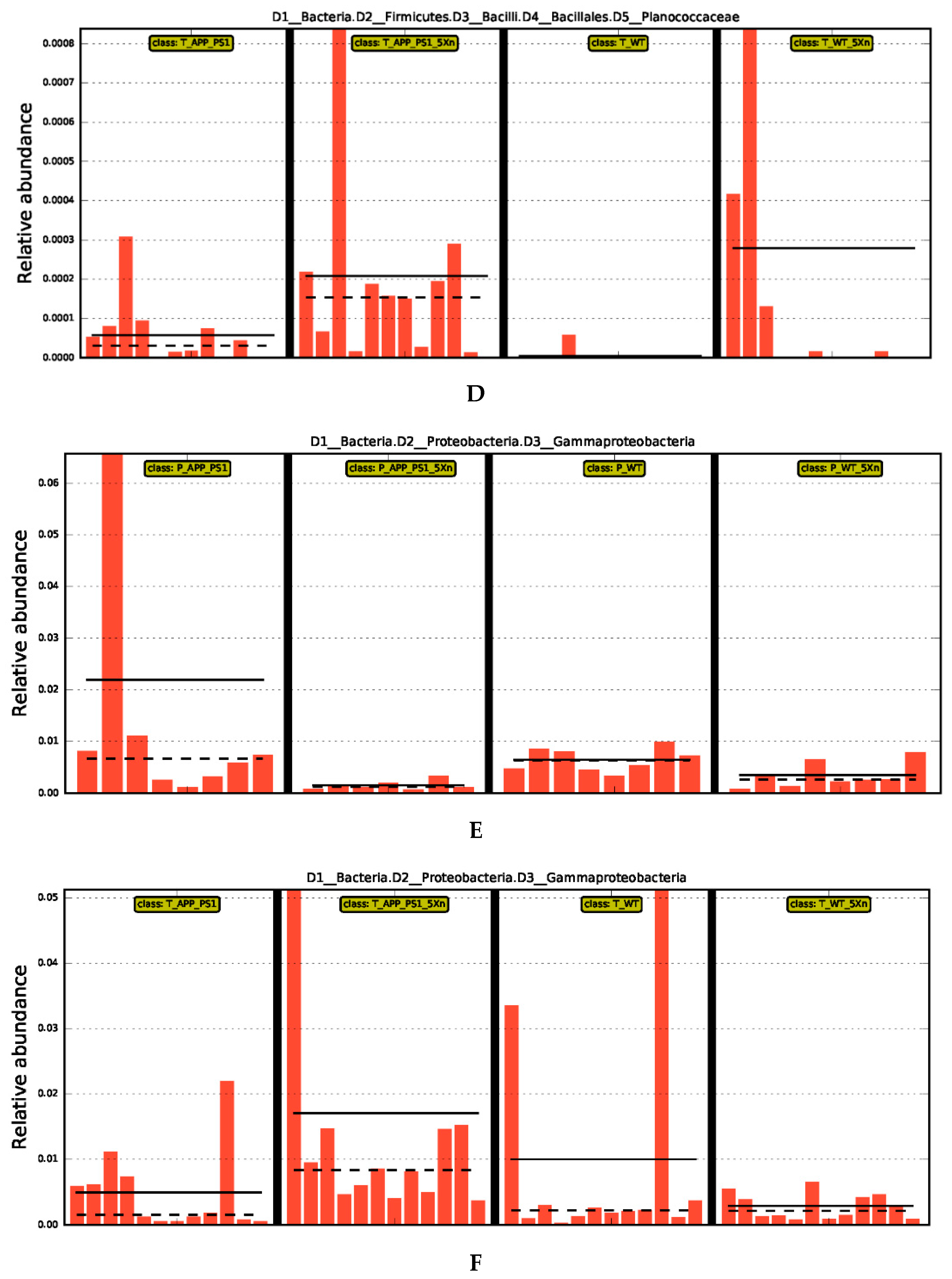
2.2. Xn Regulates the Composition of the Intestinal Microbiome
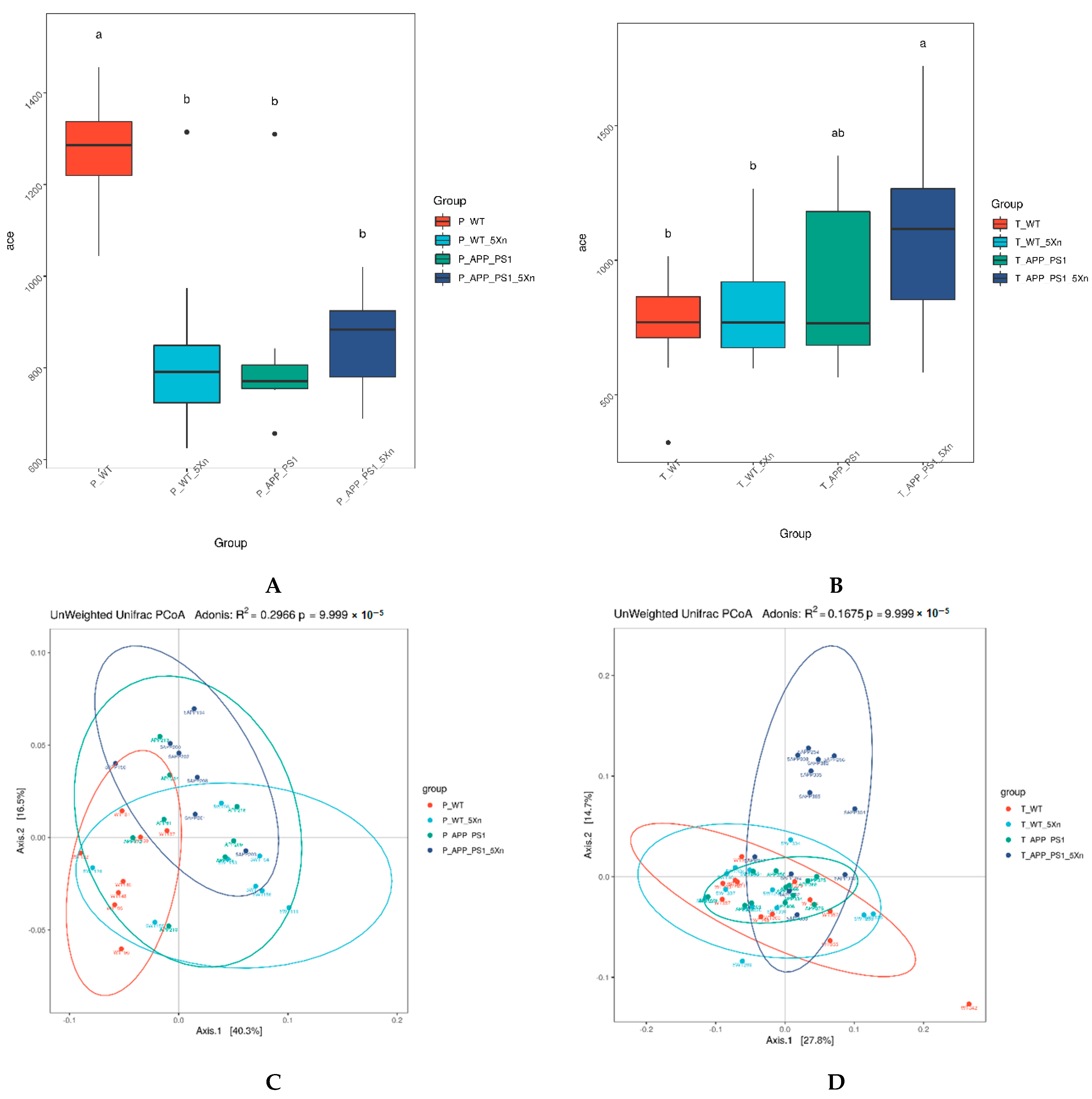
2.3. Xn Regulates the Diversity of Intestinal Microbiota Community
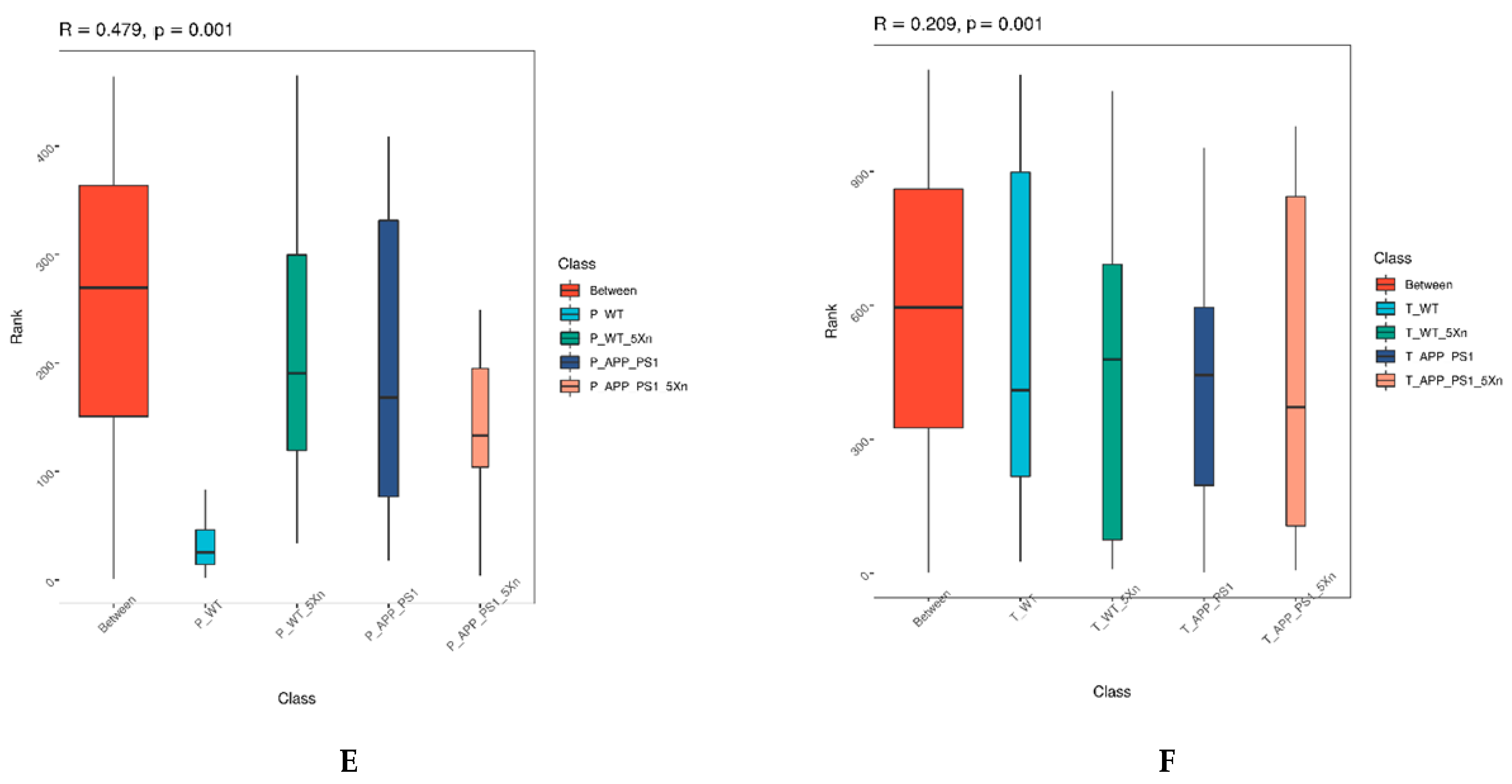
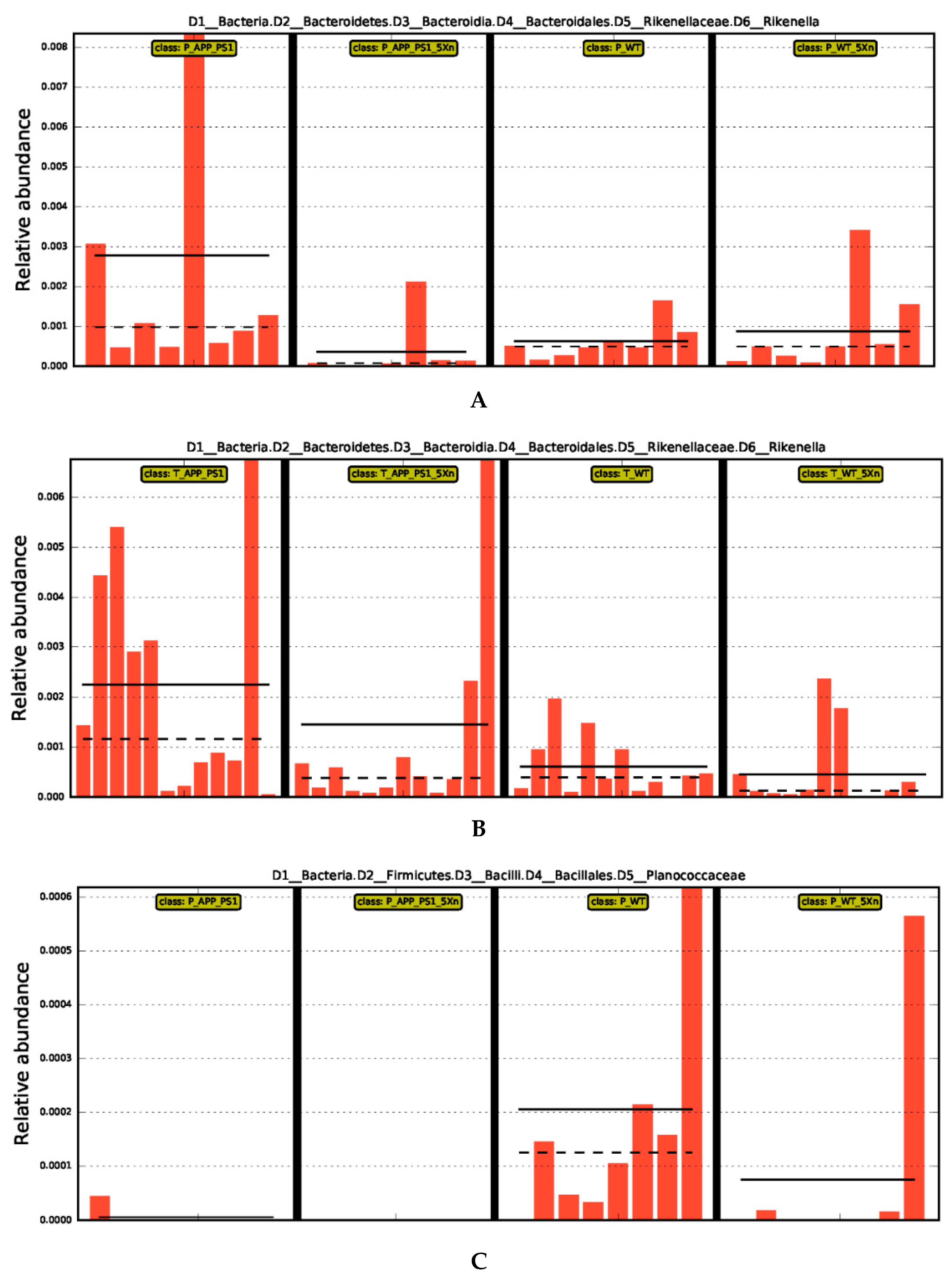
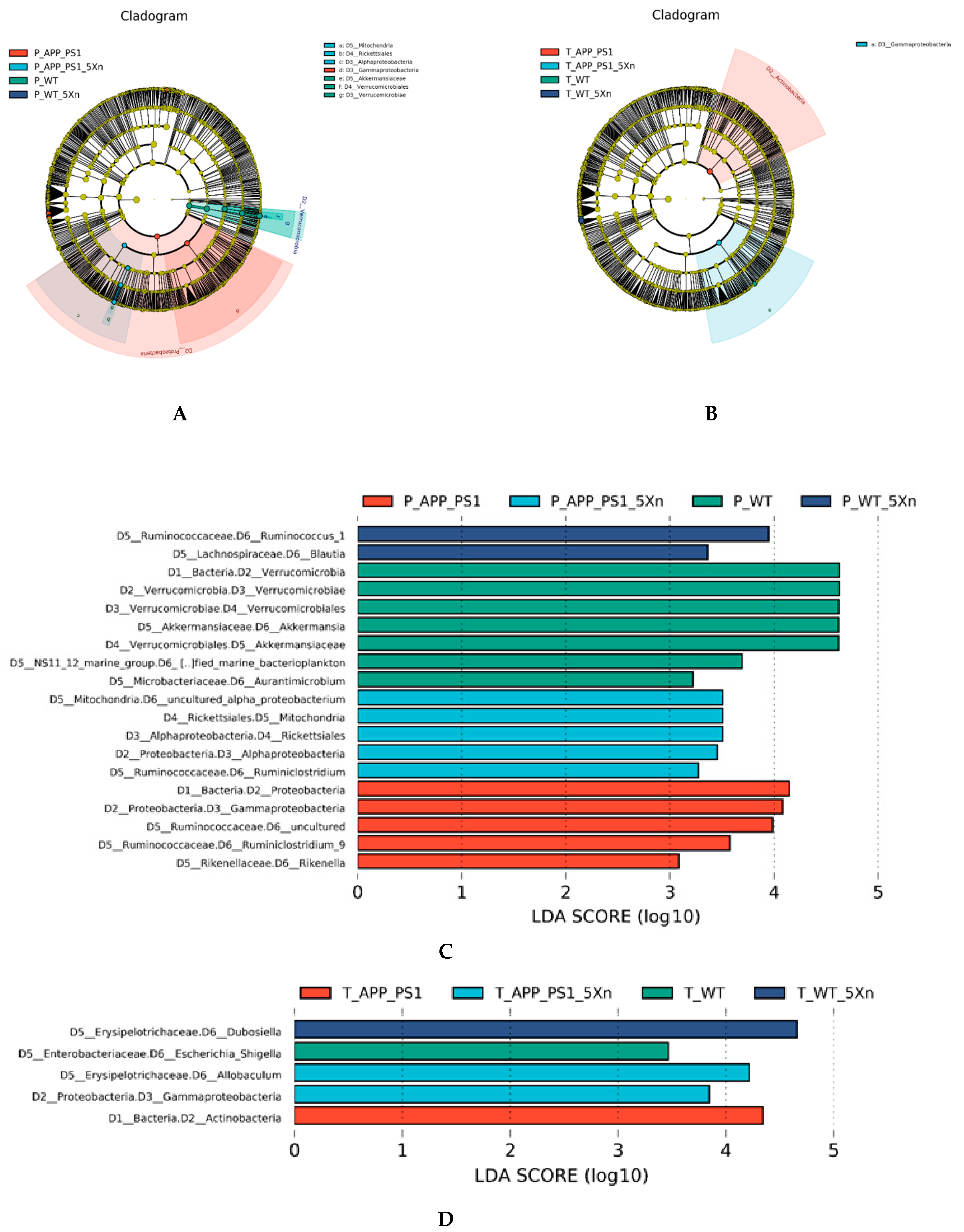
2.4. Distinct Gut Microbiome Signatures by Xn Treatment
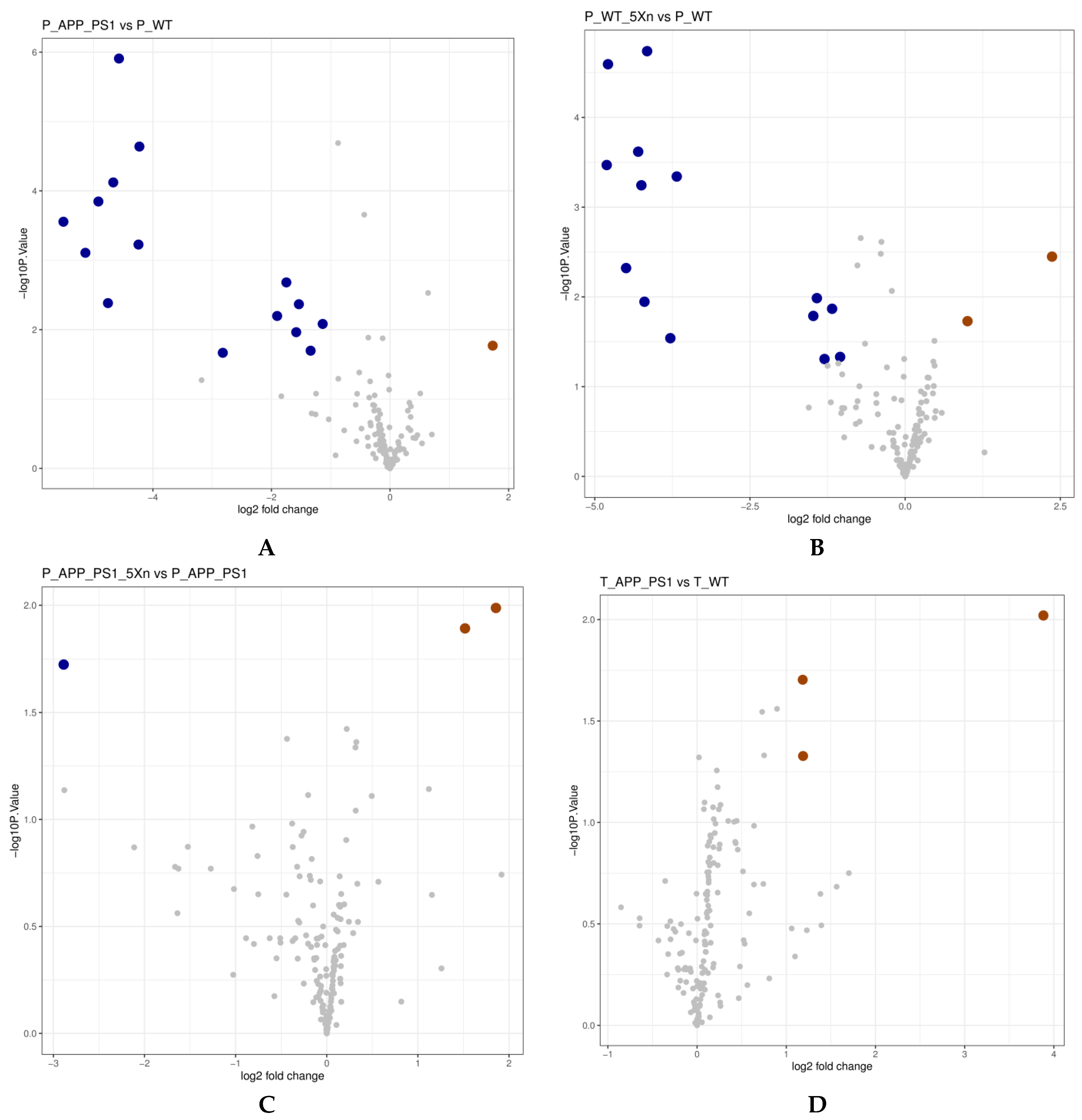
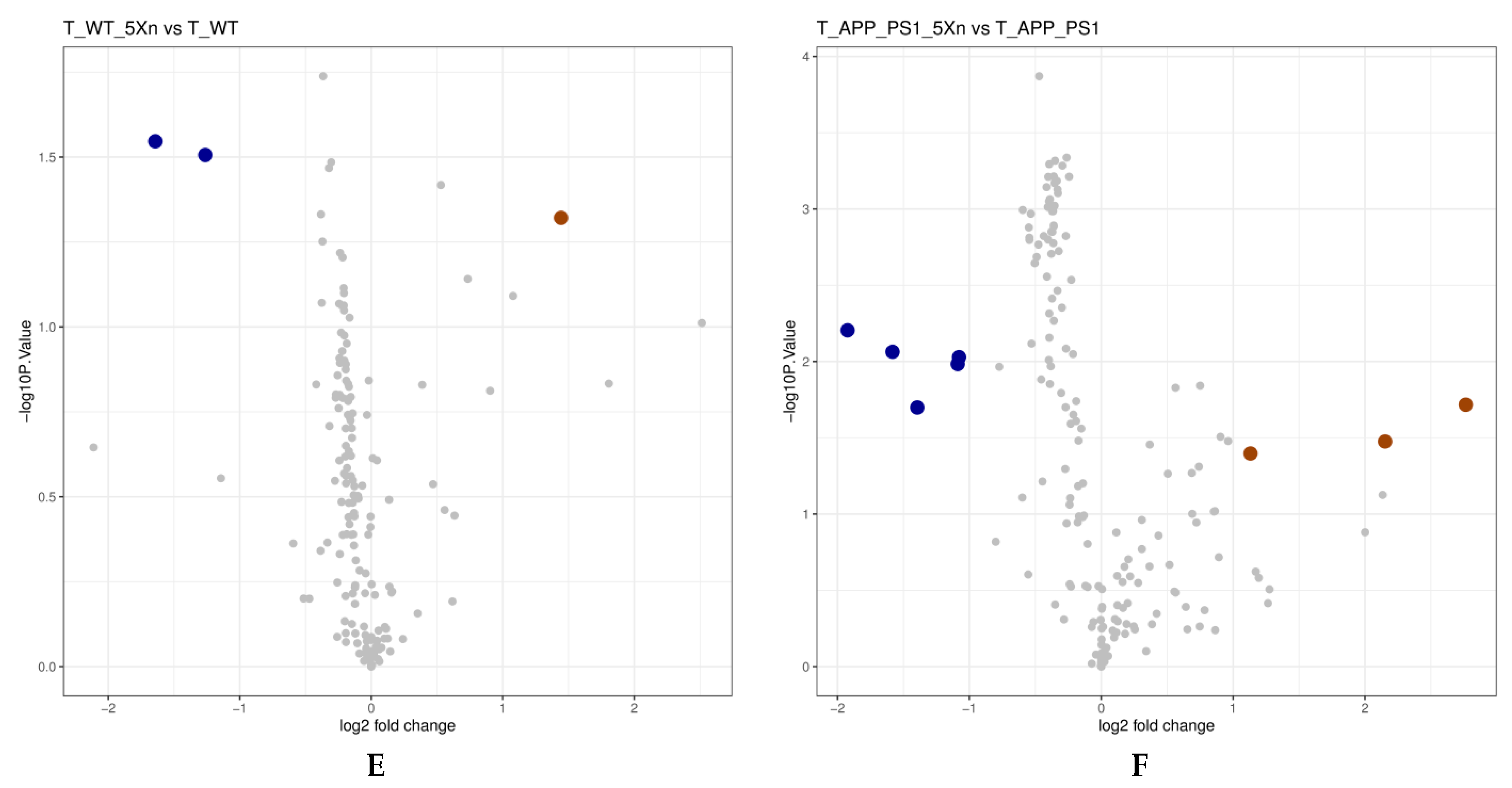
2.5. Xn Treatment May Regulate the Metabolic Function of the Gut Microbiome
3. Discussion
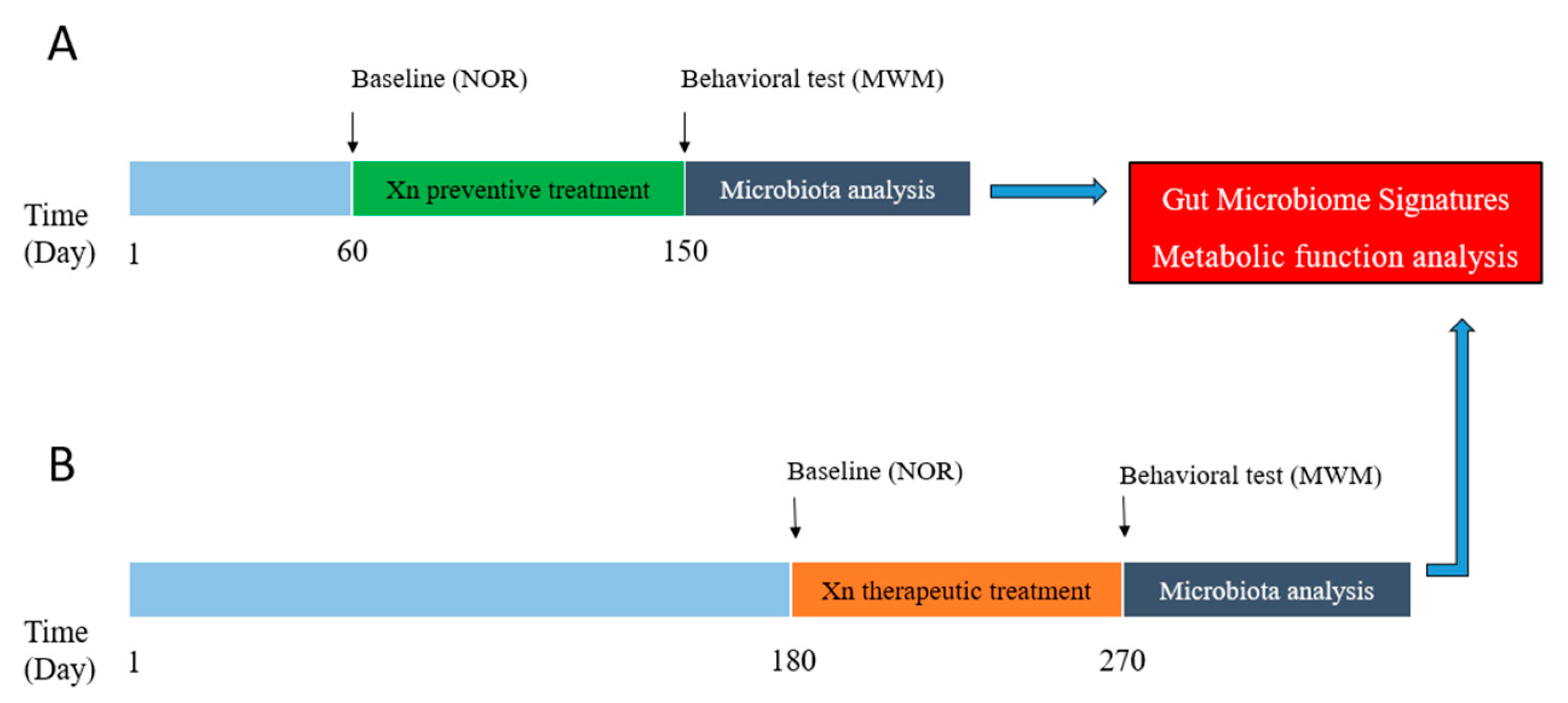
4. Materials and Methods
4.1. APP/PS1 Mice and Xn Administration
4.2. Behavioral Tests
4.3. Stool Sample Collection and Preservation
4.4. 16S rDNA Sequencing
4.5. Data Processing and Bioinformatics
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GBD 2016 Dementia Collaborators. Global, regional, and national burden of Alzheimer’s disease and other dementias, 1990-2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 88–106. [Google Scholar] [CrossRef] [Green Version]
- Jia, L.; Quan, M.; Fu, Y.; Zhao, T.; Li, Y.; Wei, C.; Tang, Y.; Qin, Q.; Wang, F.; Qiao, Y.; et al. Group for the Project of Dementia Situation in China. Dementia in China: Epidemiology, clinical management, and research advances. Lancet Neurol. 2020, 19, 81–92. [Google Scholar] [CrossRef]
- WHO. Risk Reduction of Cognitive Decline and Dementia: WHO Guidelines; World Health Organization: Geneva, Switzerland, 2019; License: CC BY-NC-SA 3.0 IGO. [Google Scholar]
- Willyard, C. How gut microbes could drive brain disorders. Nature 2021, 590, 2–25. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; O’Riordan, K.J.; Sandhu, K.; Peterson, V.; Dinan, T.G. The gut microbiome in neurological disorders. Lancet Neurol. 2020, 19, 179–194. [Google Scholar] [CrossRef]
- Kesika, P.; Suganthy, N.; Sivamaruthi, B.S.; Chaiyasut, C. Role of gut-brain axis, gut microbial composition, and probiotic intervention in Alzheimer’s disease. Life Sci. 2021, 264, 118627. [Google Scholar] [CrossRef]
- Honarpisheh, P.; Reynolds, C.R.; Blasco Conesa, M.P.; Moruno Manchon, J.F.; Putluri, N.; Bhattacharjee, M.B.; Urayama, A.; McCullough, L.D.; Ganesh, B.P. Dysregulated Gut Homeostasis Observed Prior to the Accumulation of the Brain Amyloid-β in Tg2576 Mice. Int. J. Mol. Sci. 2020, 21, 1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodiya, H.B.; Kuntz, T.; Shaik, S.M.; Baufeld, C.; Leibowitz, J.; Zhang, X.; Gottel, N.; Zhang, X.; Butovsky, O.; Gilbert, J.A.; et al. Sex-specific effects of microbiome perturbations on cerebral Aβ amyloidosis and microglia phenotypes. J. Exp. Med. 2019, 216, 1542–1560. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Wu, L.; Peng, G.; Han, Y.; Tang, R.; Ge, J.; Zhang, L.; Jia, L.; Yue, S.; Zhou, K.; et al. Altered microbiomes distinguish Alzheimer’s disease from amnestic mild cognitive impairment and health in a Chinese cohort. Brain Behav. Immun. 2019, 80, 633–643. [Google Scholar] [CrossRef]
- Zanoli, P.; Zavatti, M. Pharmacognostic and pharmacological profile of Humulus lupulus L. J. Ethnopharmacol. 2008, 116, 383–396. [Google Scholar] [CrossRef]
- Cermak, P.; Olsovska, J.; Mikyska, A.; Dusek, M.; Kadleckova, Z.; Vanicek, J.; Nyc, O.; Sigler, K.; Bostikova, V.; Bostik, P. Strong antimicrobial activity of xanthohumol and other derivatives from hops (Humulus lupulus L.) on gut anaerobic bacteria. APMIS 2017, 125, 1033–1038. [Google Scholar] [CrossRef]
- Wang, X.; Ho, S.L.; Poon, C.Y.; Yan, T.; Li, H.W.; Wong, M.S. Amyloid-β aggregation inhibitory and neuroprotective effects of xanthohumol and its derivatives for Alzheimer’s diseases. Curr. Alzheimer Res. 2019, 16, 836–842. [Google Scholar] [CrossRef] [PubMed]
- Rancán, L.; Paredes, S.D.; García, I.; Muñoz, P.; García, C.; López de Hontanar, G.; de la Fuente, M.; Vara, E.; Tresguerres, J.A.F. Protective effect of xanthohumol against age-related brain damage. J. Nutr. Biochem. 2017, 49, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, L.; Li, G.; Gao, Z. Xanthohumol protects against Azoxymethane-induced colorectal cancer in Sprague-Dawley rats. Environ. Toxicol. 2020, 35, 136–144. [Google Scholar] [CrossRef]
- Harish, V.; Haque, E.; Śmiech, M.; Taniguchi, H.; Jamieson, S.; Tewari, D.; Bishayee, A. Xanthohumol for human malignancies: Chemistry, pharmacokinetics and molecular targets. Int. J. Mol. Sci. 2021, 22, 4478. [Google Scholar] [CrossRef] [PubMed]
- Khayyal, M.T.; El-Hazek, R.M.; El-Sabbagh, W.A.; Frank, J.; Behnam, D.; Abdel-Tawab, M. Micellar solubilization enhances the anti-inflammatory effect of xanthohumol. Phytomedicine 2020, 71, 153233. [Google Scholar] [CrossRef]
- Jiao, Y.; Cao, Y.; Lu, X.; Wang, J.; Saitgareeva, A.; Kong, X.; Song, C.; Li, J.; Tian, K.; Zhang, S.; et al. Xanthohumol protects neuron from cerebral ischemia injury in experimental stroke. Mol. Biol. Rep. 2020, 47, 2417–2425. [Google Scholar] [CrossRef]
- Zhang, Y.; Bobe, G.; Revel, J.S.; Rodrigues, R.R.; Sharpton, T.J.; Fantacone, M.L.; Raslan, K.; Miranda, C.L.; Lowry, M.B.; Blakemore, P.R.; et al. Improvements in metabolic syndrome by xanthohumol derivatives are linked to altered gut microbiota and bile acid metabolism. Mol. Nutr. Food Res. 2020, 64, e1900789. [Google Scholar] [CrossRef]
- Huang, X.; Wang, J.; Chen, X.; Liu, P.; Wang, S.; Song, F.; Zhang, Z.; Zhu, F.; Huang, X.; Liu, J.; et al. The prenylflavonoid xanthohumol reduces Alzheimer-like changes and modulates multiple pathogenic molecular pathways in the Neuro2a/APPswe cell model of AD. Front. Pharmacol. 2018, 9, 199. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- de Muinck, E.J.; Trosvik, P.; Gilfillan, G.D.; Hov, J.R.; Sundaram, A.Y.M. A novel ultra high-throughput 16S rRNA gene amplicon sequencing library preparation method for the Illumina HiSeq platform. Microbiome 2017, 5, 68. [Google Scholar] [CrossRef]
- Wang, X.; Sun, G.; Feng, T.; Zhang, J.; Huang, X.; Wang, T.; Xie, Z.; Chu, X.; Yang, J.; Wang, H.; et al. Sodium oligomannate therapeutically remodels gut microbiota and suppresses gut bacterial amino acids-shaped neuroinflammation to inhibit Alzheimer’s disease progression. Cell Res. 2019, 29, 787–803. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Kuang, W.; Chen, W.; Xu, W.; Zhang, L.; Li, Y.; Li, H.; Peng, Y.; Chen, Y.; Wang, B.; et al. A phase II randomized trial of sodium oligomannate in Alzheimer’s dementia. Alzheimers Res. Ther. 2020, 12, 110. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lei, X.; Xie, Z.; Zhang, X.; Cheng, X.; Zhou, W.; Zhang, Y. CA-30, an oligosaccharide fraction derived from Liuwei Dihuang decoction, ameliorates cognitive deterioration via the intestinal microbiome in the senescence-accelerated mouse prone 8 strain. Aging 2019, 11, 3463–3486. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Ko, B.S.; Ryuk, J.A.; Park, S. Tetragonia tetragonioides protected against memory dysfunction by elevating hippocampal amyloid-β deposition through potentiating insulin signaling and altering gut microbiome composition. Int. J. Mol. Sci. 2020, 21, 2900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamzow, D.R.; Elias, V.; Legette, L.L.; Choi, J.; Stevens, J.F.; Magnusson, K.R. Xanthohumol improved cognitive flexibility in young mice. Behav. Brain Res. 2014, 275, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, B.; Thielmann, J.; Hickisch, A.; Muranyi, P.; Wunderlich, J.; Hauser, C. Antimicrobial activity of hop extracts against foodborne pathogens for meat applications. J. Appl. Microbiol. 2015, 118, 648–657. [Google Scholar] [CrossRef]
- Bäuerl, C.; Collado, M.C.; Diaz Cuevas, A.; Viña, J.; Pérez Martínez, G. Shifts in gut microbiota composition in an APP/PSS1 transgenic mouse model of Alzheimer’s disease during lifespan. Lett. Appl. Microbiol. 2018, 66, 464–471. [Google Scholar] [CrossRef]
- Wang, G.; Zhou, H.H.; Luo, L.; Qin, L.Q.; Yin, J.; Yu, Z.; Zhang, L.; Wan, Z. Voluntary wheel running is capable of improving cognitive function only in the young but not the middle-aged male APPSwe/PS1De9 mice. Neurochem. Int. 2021, 145, 105010. [Google Scholar] [CrossRef]
- Zheng, P.; Zeng, B.; Liu, M.; Chen, J.; Pan, J.; Han, Y.; Liu, Y.; Cheng, K.; Zhou, C.; Wang, H.; et al. The gut microbiome from patients with schizophrenia modulates the glutamate-glutamine-GABA cycle and schizophrenia-relevant behaviors in mice. Sci. Adv. 2019, 5, eaau8317. [Google Scholar] [CrossRef] [Green Version]
- Kamaz, M.; Jones, S.M.; Qian, X.; Watts, M.J.; Zhang, W.; Wickramasinghe, S.R. Atrazine removal from municipal wastewater using a membrane bioreactor. Int. J. Environ. Res. Public Health 2020, 17, 2567. [Google Scholar] [CrossRef]
- Legette, L.; Ma, L.; Reed, R.L.; Miranda, C.L.; Christensen, J.M.; Rodriguez-Proteau, R.; Stevens, J.F. Pharmacokinetics of xanthohumol and metabolites in rats after oral and intravenous administration. Mol. Nutr. Food Res. 2012, 56, 466–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Hansen, P.E.; Wang, G.; Qiu, L.; Dong, J.; Yin, H.; Qian, Z.; Yang, M.; Miao, J. Pharmacological profile of xanthohumol, a prenylated flavonoid from hops (Humulus lupulus). Molecules 2015, 20, 754–779. [Google Scholar] [CrossRef] [PubMed]
- Leger, M.; Quiedeville, A.; Bouet, V.; Haelewyn, B.; Boulouard, M.; Schumann-Bard, P.; Freret, T. Object recognition test in mice. Nat. Protoc. 2013, 8, 2531–2537. [Google Scholar] [CrossRef]
- Vorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef] [Green Version]
- Bradley, R.; Langley, B.O.; Ryan, J.J.; Phipps, J.; Hanes, D.A.; Stack, E.; Jansson, J.K.; Metz, T.O.; Stevens, J.F. Xanthohumol microbiome and signature in healthy adults (the XMaS trial): A phase I triple-masked, placebo-controlled clinical trial. Trials 2020, 21, 835. [Google Scholar] [CrossRef] [PubMed]
- Langley, B.O.; Ryan, J.J.; Hanes, D.; Phipps, J.; Stack, E.; Metz, T.O.; Stevens, J.F.; Bradley, R. Xanthohumol microbiome and signature in healthy adults (the XMaS trial): Safety and tolerability results of a phase I triple-masked, placebo-controlled clinical trial. Mol. Nutr. Food Res. 2021, 65, e2001170. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Description | P_APP_PS1_vs_P_WT | P_WT_5Xn_vs_P_WT | P_APP_PS1_5Xn_vs_P_APP_PS1 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| log2 (Fold Change) | p Value | Up/Down | log2 (Fold Change) | p Value | Up/Down | log2 (Fold Change) | p Value | Up/Down | ||
| ko05010 | Alzheimer′s disease | 1.732027242 | 0.017025339 | up | 2.366161715 | 0.003558436 | up | |||
| ko04974 | Protein digestion and absorption | 1.00459934 | 0.018638107 | up | ||||||
| ko00830 | Retinol metabolism | −4.572428141 | 1.24 × 10−5 | down | −4.157153572 | 1.83 × 10−5 | down | |||
| ko04142 | Lysosome | −4.227876257 | 2.30 × 10−5 | down | −3.679825761 | 0.000455767 | down | |||
| ko00980 | Metabolism of xenobiotics by cytochrome P450 | −4.66899817 | 7.55 × 10−5 | down | −4.787486299 | 2.55 × 10−5 | down | |||
| ko00363 | Bisphenol degradation | −4.919555706 | 0.000142318 | down | −4.808225969 | 0.000339327 | down | |||
| ko00195 | Photosynthesis | −5.509890045 | 0.000278555 | down | −4.495865401 | 0.00477363 | down | |||
| ko00642 | Ethylbenzene degradation | −4.243304316 | 0.000594088 | down | −4.249603903 | 0.000570651 | down | |||
| ko00523 | Polyketide sugar unit biosynthesis | −5.139057953 | 0.000781219 | down | −4.199953052 | 0.011321054 | down | |||
| ko00311 | Penicillin and cephalosporin biosynthesis | −1.749057294 | 0.002085877 | down | −1.299383141 | 0.049212447 | down | 1.517322773 | 0.012792112 | up |
| ko00253 | Tetracycline biosynthesis | −4.758378649 | 0.004146843 | down | −3.783153775 | 0.028842448 | down | |||
| ko00364 | Fluorobenzoate degradation | −1.537476491 | 0.004302729 | down | −1.422606062 | 0.010323156 | down | |||
| ko00791 | Atrazine degradation | −1.904490251 | 0.006353534 | down | 1.85619649 | 0.010297892 | up | |||
| ko00906 | Carotenoid biosynthesis | −1.136194803 | 0.008280216 | down | −1.047940043 | 0.046641794 | down | |||
| ko05200 | Pathways in cancer | −1.582997558 | 0.010892181 | down | ||||||
| ko00930 | Caprolactam degradation | −1.339589301 | 0.020127255 | down | −1.479654328 | 0.016294803 | down | |||
| ko00903 | Limonene and pinene degradation | −2.82113561 | 0.021578285 | down | −4.301375008 | 0.000240924 | down | |||
| ko00643 | Styrene degradation | −1.177109587 | 0.013561769 | down | ||||||
| ko00622 | Xylene degradation | −2.887030853 | 0.018878669 | down | ||||||
| Number | Description | T_APP_PS1_vs_T_WT | T_WT_5Xn_vs_T_WT | T_APP_PS1_5Xn_vs_T_APP_PS1 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| log2 (Fold Change) | p Value | Up/Down | log2 (Fold Change) | p Value | Up/Down | log2 (Fold Change) | p Value | Up/Down | ||
| ko00195 | Photosynthesis | 3.882765365 | 0.009566875 | up | ||||||
| ko00930 | Caprolactam degradation | 1.184272747 | 0.019807356 | up | ||||||
| ko05150 | Staphylococcus aureus infection | 1.188139551 | 0.047056443 | up | −1.583094991 | 0.008640976 | down | |||
| ko00472 | D-Arginine and D-ornithine metabolism | 1.443191914 | 0.047738796 | up | ||||||
| ko00791 | Atrazine degradation | −1.642519851 | 0.028425143 | down | −1.924538818 | 0.00624072 | down | |||
| ko00311 | Penicillin and cephalosporin biosynthesis | −1.262387166 | 0.031172616 | down | −1.395234979 | 0.020013002 | down | |||
| ko00623 | Toluene degradation | 2.764724413 | 0.019188937 | up | ||||||
| ko04142 | Lysosome | 2.152419965 | 0.033435027 | up | ||||||
| ko00364 | Fluorobenzoate degradation | 1.131389736 | 0.040093241 | up | ||||||
| ko00621 | Dioxin degradation | −1.078840557 | 0.009368244 | down | ||||||
| ko02060 | Phosphotransferase system (PTS) | −1.088900276 | 0.010383481 | down | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; He, K.; Wu, D.; Zhou, L.; Li, G.; Lin, Z.; Yang, X.; Liu, J.; Pui Man Hoi, M. Natural Dietary Compound Xanthohumol Regulates the Gut Microbiota and Its Metabolic Profile in a Mouse Model of Alzheimer’s Disease. Molecules 2022, 27, 1281. https://doi.org/10.3390/molecules27041281
Liu W, He K, Wu D, Zhou L, Li G, Lin Z, Yang X, Liu J, Pui Man Hoi M. Natural Dietary Compound Xanthohumol Regulates the Gut Microbiota and Its Metabolic Profile in a Mouse Model of Alzheimer’s Disease. Molecules. 2022; 27(4):1281. https://doi.org/10.3390/molecules27041281
Chicago/Turabian StyleLiu, Wei, Kaiwu He, Desheng Wu, Li Zhou, Guowei Li, Zequn Lin, Xifei Yang, Jianjun Liu, and Maggie Pui Man Hoi. 2022. "Natural Dietary Compound Xanthohumol Regulates the Gut Microbiota and Its Metabolic Profile in a Mouse Model of Alzheimer’s Disease" Molecules 27, no. 4: 1281. https://doi.org/10.3390/molecules27041281