
Yeast GH30 Xylanase from Sugiyamaella lignohabitans Is a Glucuronoxylanase with Auxiliary Xylobiohydrolase Activity §


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
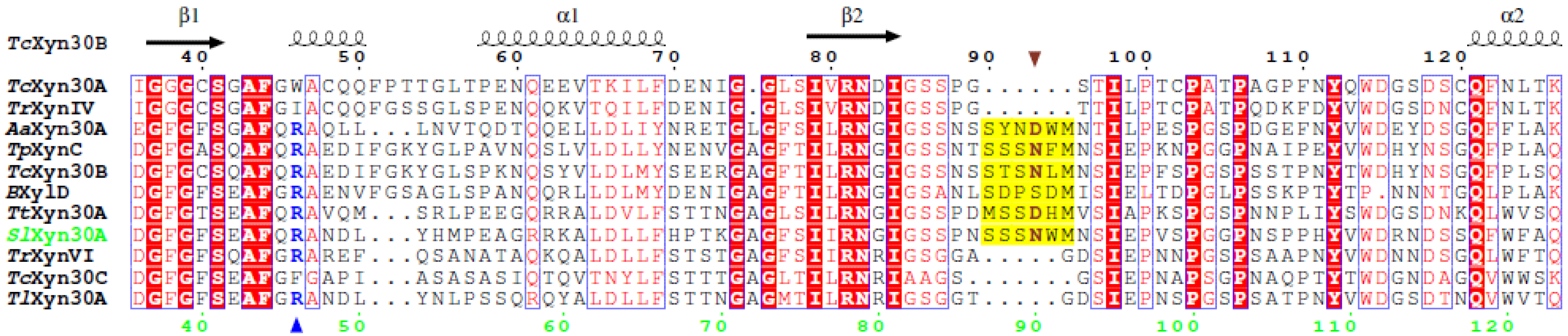
2.1. Sequence Analysis of SlXyn30A
2.2. Recombinant Strain Selection
2.3. Thermal and pH Optima and Stability
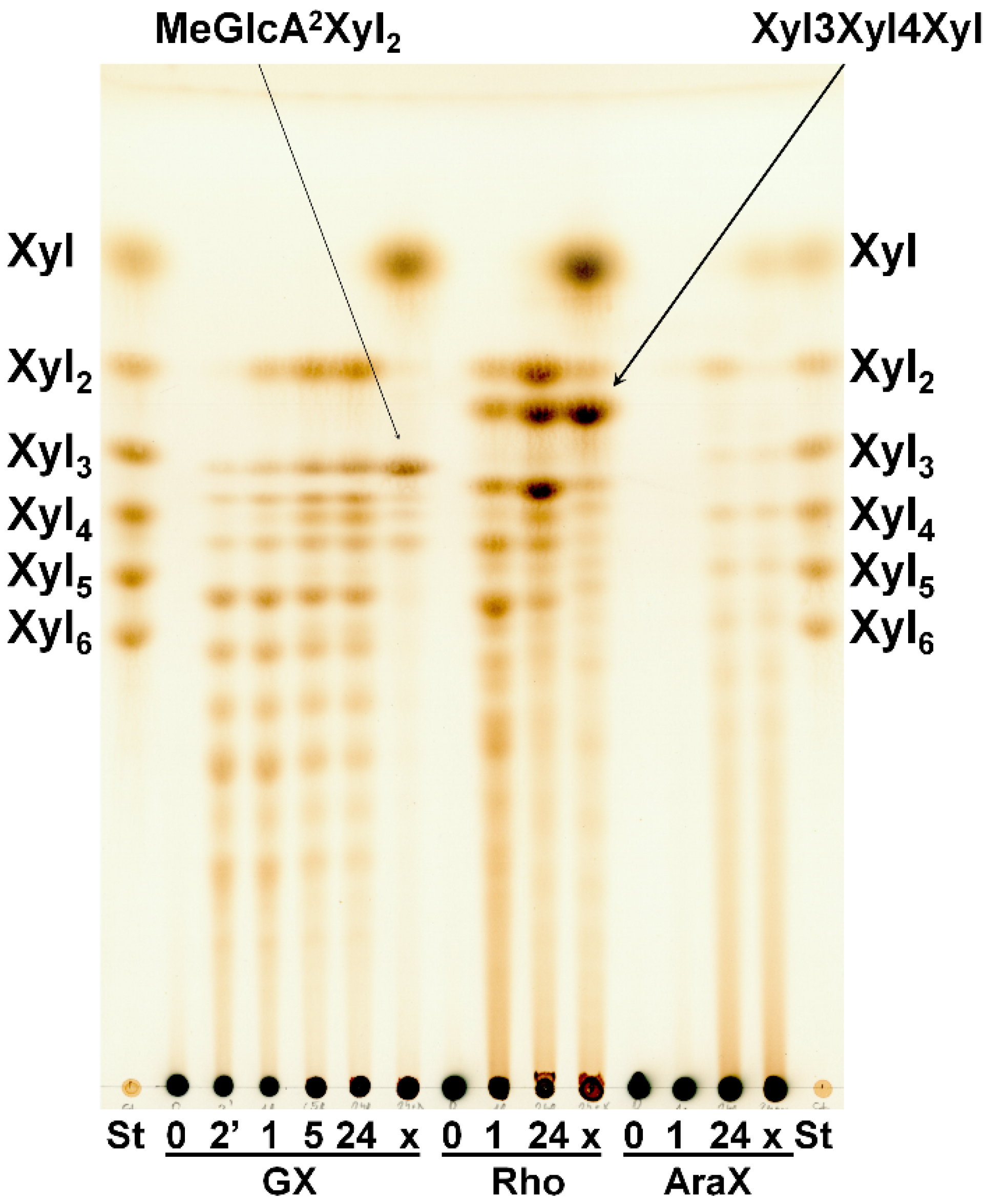
2.4. Hydrolysis of Polysaccharides
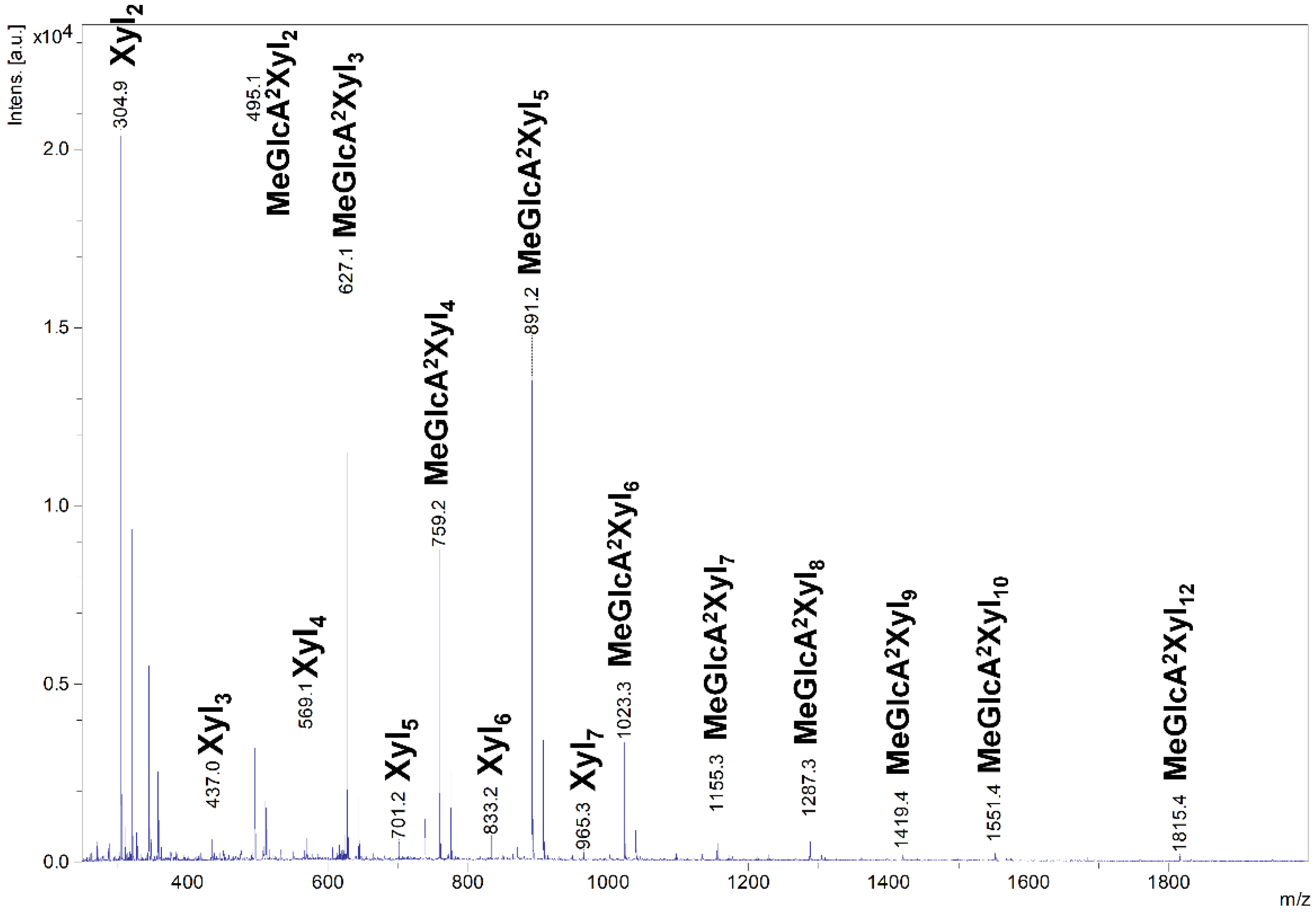
2.5. Hydrolysis of Oligosaccharides
3. Discussion
4. Materials and Methods
4.1. Substrates, Standards and Enzymes
4.2. Amino Acid Sequence Comparison
4.3. Recombinant Strain Preparation
4.4. Determination of pH and Temperature Optimum and Temperature Stability
4.5. Hydrolysis of Polysaccharides and Oligosaccharides
4.6. MALDI-ToF MS
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
| EX | endo-β-1,4-xylanase |
| XOs | xylooligosaccharides |
| GH | glycoside hydrolase |
| GX | 4-O-methylglucuronoxylan |
| MeGlcA | 4-O-methylglucuronic acid |
| AraX | arabinoxylan |
| Rho | rhodymenan (linear β-1,3-1,4-xylan) |
| Xylp | xylopyranosyl |
| Xyln | linear xylooligosaccharide composed of n d-xylopyranosyl residues linked by β-1,4-linkages |
| MeGlcAzXyln | Xyln wherein zth xylopyranosyl residue counted from reducing end is α-glycosylated at position 2 by MeGlcA |
| NP | 4-nitrophenyl |
| YPD | yeast extract peptone dextrose |
| YNB | yeast nitrogen base |
References
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The Carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamichi, Y.; Fouquet, T.; Ito, S.; Watanabe, M.; Matsushika, A.; Inoue, H. Structural and functional characterization of a GH30-7 xylanase B from the filamentous fungus Talaromyces cellulolyticus. J. Biol. Chem. 2019, 294, 4065–4078. [Google Scholar] [CrossRef] [PubMed]
- Katsimpouras, C.; Dedes, G.; Thomaidis, N.S.; Topakas, E. A novel fungal GH30 xylanase with xylobiohydrolase auxiliary activity. Biotechnol. Biofuels 2019, 12, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamichi, Y.; Watanabe, M.; Matsushika, A.; Inoue, H. Substrate recognition by a bifunctional GH30-7 xylanase B from Talaromyces cellulolyticus. FEBS Open Bio 2020, 10, 1180–1189. [Google Scholar] [CrossRef]
- Nakamichi, Y.; Fouquet, T.; Ito, S.; Matsushika, A.; Inoue, H. Mode of action of GH30-7 reducing-end xylose-releasing exoxylanase A (Xyn30A) from the filamentous fungus Talaromyces cellulolyticus. Appl. Environ. Microbiol. 2019, 85, e00552-19. [Google Scholar] [CrossRef] [Green Version]
- Nakamichi, Y.; Fujii, T.; Fouquet, T.; Matsushika, A.; Inoue, H. GH30-7 endoxylanase C from the filamentous fungus Talaromyces cellulolyticus. Appl. Environ. Microbiol. 2019, 85, e0144219. [Google Scholar] [CrossRef]
- Šuchová, K.; Puchart, V.; Spodsberg, N.; Mørkeberg Krogh, K.B.R.; Biely, P. A novel GH30 xylobiohydrolase from Acremonium alcalophilum releasing xylobiose from the non-reducing end. Enzyme Microb. Technol. 2020, 134, 109484. [Google Scholar] [CrossRef]
- Šuchová, K.; Puchart, V.; Biely, P. A novel bacterial GH30 xylobiohydrolase from Hungateiclostridium clariflavum. Appl. Microbiol. Biotechnol. 2021, 105, 185–195. [Google Scholar] [CrossRef]
- Nikolaivits, E.; Pentari, C.; Kosinas, C.; Feiler, C.G.; Spiliopoulou, M.; Weiss, M.S.; Dimarogona, M.; Topakas, E. Unique features of the bifunctional GH30 from Thermothelomyces thermophila revealed by structural and mutational studies. Carbohydr. Polym. 2021, 273, 118553. [Google Scholar] [CrossRef]
- Šuchová, K.; Spodsberg, N.; Mørkeberg Krogh, K.B.R.; Biely, P.; Puchart, V. Non-Specific GH30_7 Endo-β-1,4-xylanase from Talaromyces leycettanus. Molecules 2021, 26, 4614. [Google Scholar] [CrossRef]
- Šuchová, K.; Puchart, V.; Spodsberg, N.; Mørkeberg Krogh, K.B.R.; Biely, P. Catalytic Diversity of GH30 Xylanases. Molecules 2021, 26, 4528. [Google Scholar] [CrossRef] [PubMed]
- Puchart, V.; Šuchová, K.; Biely, P. Xylanases of glycoside hydrolase family 30—An overview. Biotechnol. Adv. 2021, 47, 107704. [Google Scholar] [CrossRef] [PubMed]
- Biely, P.; Puchart, V.; Stringer, M.A.; Mørkeberg Krogh, K.B.R. Trichoderma reesei XYN VI–a novel appendage dependent eukaryotic glucuronoxylan hydrolase. FEBS J. 2014, 281, 3894–3903. [Google Scholar] [CrossRef] [PubMed]
- Vršanská, M.; Kolenová, K.; Puchart, V.; Biely, P. Mode of action of glycoside hydrolase family 5 glucuronoxylan xylanohydrolase from Erwinia chrysanthemi. FEBS J. 2007, 274, 1666–1677. [Google Scholar] [CrossRef]
- St. John, F.J.; Rice, J.D.; Preston, J.F. Characterization of XynC from Bacillus subtilis subsp. subtilis strain 168 and analysis of its role in depolymerization of glucuronoxylan. J. Bacteriol. 2006, 188, 8617–8626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leathers, T.D. Color variants of Aureobasidium pullulans overproduce xylanase with extremely high specific activity. Appl. Environ. Microbiol. 1986, 52, 1026–1030. [Google Scholar] [CrossRef] [Green Version]
- Li, X.L.; Zhang, Z.Q.; Dean, J.F.D.; Eriksson, K.E.L.; Ljungdahl, L.G. Purification and characterization of a new xylanase (APX-II) from the fungus Aureobasidium pullulans Y-2311-1. Appl. Environ. Microbiol. 1993, 59, 3212–3218. [Google Scholar] [CrossRef] [Green Version]
- Biely, P.; Vršanská, M.; Krátky, Z. Xylan-degrading enzymes of the yeast Cryptococcus albidus: Identification and cellular localization. Eur. J. Biochem. 1980, 108, 313–321. [Google Scholar] [CrossRef]
- Petrescu, I.; Lamotte-Brasseur, J.; Chessa, J.-P.; Ntarima, P.; Claeyssens, M.; Devreese, B.; Marino, G.; Gerday, C. Xylanase from the psychrophilic yeast Cryptococcus adeliae. Extremophiles 2000, 4, 137–144. [Google Scholar] [CrossRef]
- Parachin, N.S.; Siqueira, S.; de Faria, F.P.; Torres, F.A.G.; de Moraesa, L.M.P. Xylanases from Cryptococcus flavus isolate I-11: Enzymatic profile, isolation and heterologous expression of CfXYN1 in Saccharomyces cerevisiae. J. Mol. Catal. B Enzym. 2009, 59, 52–57. [Google Scholar] [CrossRef]
- Ding, C.; Li, M.X.; Hu, Y. High-activity production of xylanase by Pichia stipitis: Purification, characterization, kinetic evaluation and xylooligosaccharides production. Int. J. Biol. Macromol. 2018, 117, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Adsul, M.G.; Bastawde, K.B.; Gokhale, D.V. Biochemical characterization of two xylanases from yeast Pseudozyma hubeiensis producing only xylooligosaccharides. Bioresour. Technol. 2009, 100, 6488–6495. [Google Scholar] [CrossRef] [PubMed]
- Ravn, J.L.; Engqvist, M.K.M.; Larsbrink, J.; Geijer, C. CAZyme prediction in ascomycetous yeast genomes guides discovery of novel xylanolytic species with diverse capacities for hemicellulose hydrolysis. Biotechnol. Biofuels 2021, 14, 150. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Robnett, C.J. Multigene phylogenetic analysis of the Trichomonascus, Wickerhamiella and Zygoascus yeast clades, and the proposal of Sugiyamaella gen. nov. and 14 new species combinations. FEMS Yeast Res. 2007, 7, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Houseknecht, J.L.; Hart, E.L.; Suh, S.O.; Zhou, J.J. Yeasts in the Sugiyamaella clade associated with wood-ingesting beetles and the proposal of Candida bullrunensis sp. nov. Int. J. Syst. Evol. Microbiol. 2011, 61, 1751–1756. [Google Scholar] [CrossRef]
- Sena, L.M.; Morais, C.G.; Lopes, M.R.; Santos, R.O.; Uetanabaro, A.P.; Morais, P.B.; Vital, M.J.; de Morais, M.A., Jr.; Lachance, M.A.; Rosa, C.A. d-Xylose fermentation, xylitol production and xylanase activities by seven new species of Sugiyamaella. Antonie Leeuwenhoek 2017, 110, 53–67. [Google Scholar] [CrossRef]
- Bellasio, M.; Mattanovich, D.; Sauer, M.; Marx, H. Organic acids from lignocellulose: Candida lignohabitans as a new microbial cell factory. J. Ind. Microbiol. Biotechnol. 2015, 42, 681–691. [Google Scholar] [CrossRef]
- Ebringerová, A.; Kramár, A.; Rendoš, F.; Domanský, R. Stepwise extraction of hemicellulose from wood of white beech (Carpinus betulus L.). Holzforschung 1967, 21, 74–77. [Google Scholar] [CrossRef]
- Biely, P.; Vršanská, M.; Tenkanen, M.; Kluepfel, D. Endo-β-1,4-xylanases: Differences in catalytic properties. J. Biotechnol. 1997, 57, 151–166. [Google Scholar] [CrossRef]
- Hirsch, J.; Kováč, P. Alternative synthesis of methylated sugars. 21. Synthesis of 2 isomeric methyl β-d-xylotriosides containing a (1-2)-β-linkage. Carbohydr. Res. 1979, 77, 241–244. [Google Scholar] [CrossRef]
- Hirsch, J.; Petráková, E. Sequential synthesis and C-13 NMR-spectra of methyl 3-O and 2-O-(β-d-xylobiosyl)-β-d-xylopyranosides. Chem. Pap. 1984, 38, 409–417. [Google Scholar]
- Hirsch, J.; Petráková, E.; Schraml, J. Stereoselective synthesis and 13C-N.m.r. spectra of two isomeric methyl β-glycosides of trisaccharides related to arabinoxylan. Carbohydr. Res. 1984, 131, 219–226. [Google Scholar] [CrossRef]
- Kováč, P. Alternative synthesis of methylated sugars. 17. Synthesis of methyl 3,4-di-O-(β-d-xylopyranosyl)-β-d-xylopyranoside, a methyl β-xylotrioside related to branched xylans. Collect. Czechoslov. Chem. Commun. 1979, 44, 928–932. [Google Scholar] [CrossRef]
- Kováč, P.; Hirsch, J. Alternative synthesis of methylated sugars. 23. Stepwise synthesis of methyl 4-O-[3-O-(β-d-xylopyranosyl)-β-Dxylopyranosyl]-β-d-xylopyranoside. Carbohydr. Res. 1980, 79, 303–307. [Google Scholar] [CrossRef]
- Kováč, P.; Hirsch, J. Alternative synthesis of methylated sugars. 24. Sequential synthesis and 13C-N.M.R spectra of methyl β-glycosides of (1-4)-β-d-xylo-oligosaccharides. Carbohydr. Res. 1982, 100, 177–193. [Google Scholar] [CrossRef]
- Biely, P.; Hirsch, J.; la Grange, D.C.; van Zyl, W.H.; Prior, B.A. A chromogenic substrate for a β-xylosidase-coupled assay of α-glucuronidase. Anal. Biochem. 2000, 286, 289–294. [Google Scholar] [CrossRef]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBLEBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [Green Version]
- Rosenbergová, Z.; Kántorová, K.; Šimkovič, M.; Breier, A.; Rebroš, M. Optimisation of recombinant myrosinase production in Pichia pastoris. Int. J. Mol. Sci. 2021, 22, 3677. [Google Scholar] [CrossRef]
- Lin-Cereghino, J.; Wong, W.W.; Xiong, S.; Giang, W.; Luong, L.T.; Vu, J.; Johnson, S.D.; Lin-Cereghino, G.P. Condensed Protocol for Competent Cell Preparation and Transformation of the Methylotrophic Yeast Pichia pastoris. Biotechniques 2005, 38, 44–48. [Google Scholar] [CrossRef] [Green Version]
- Markošová, K.; Weignerová, L.; Rosenberg, M.; Křen, V.; Rebroš, M. Upscale of recombinant α-l-rhamnosidase production by Pichia pastoris MutS strain. Front. Microbiol. 2015, 6, 1140. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Paleg, L.G. Citric acid interference in the estimation of reducing sugars with alkaline copper reagents. Anal. Chem. 1959, 31, 1092–1094. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šuchová, K.; Chyba, A.; Hegyi, Z.; Rebroš, M.; Puchart, V. Yeast GH30 Xylanase from Sugiyamaella lignohabitans Is a Glucuronoxylanase with Auxiliary Xylobiohydrolase Activity. Molecules 2022, 27, 751. https://doi.org/10.3390/molecules27030751
Šuchová K, Chyba A, Hegyi Z, Rebroš M, Puchart V. Yeast GH30 Xylanase from Sugiyamaella lignohabitans Is a Glucuronoxylanase with Auxiliary Xylobiohydrolase Activity. Molecules. 2022; 27(3):751. https://doi.org/10.3390/molecules27030751
Chicago/Turabian StyleŠuchová, Katarína, Andrej Chyba, Zuzana Hegyi, Martin Rebroš, and Vladimír Puchart. 2022. "Yeast GH30 Xylanase from Sugiyamaella lignohabitans Is a Glucuronoxylanase with Auxiliary Xylobiohydrolase Activity" Molecules 27, no. 3: 751. https://doi.org/10.3390/molecules27030751