
Thyroid-Stimulating Hormone Favors Runx2-Mediated Matrix Mineralization in HOS and SaOS2 Cells: An In Vitro and In Silico Approach

 , , , ,
, , , ,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results
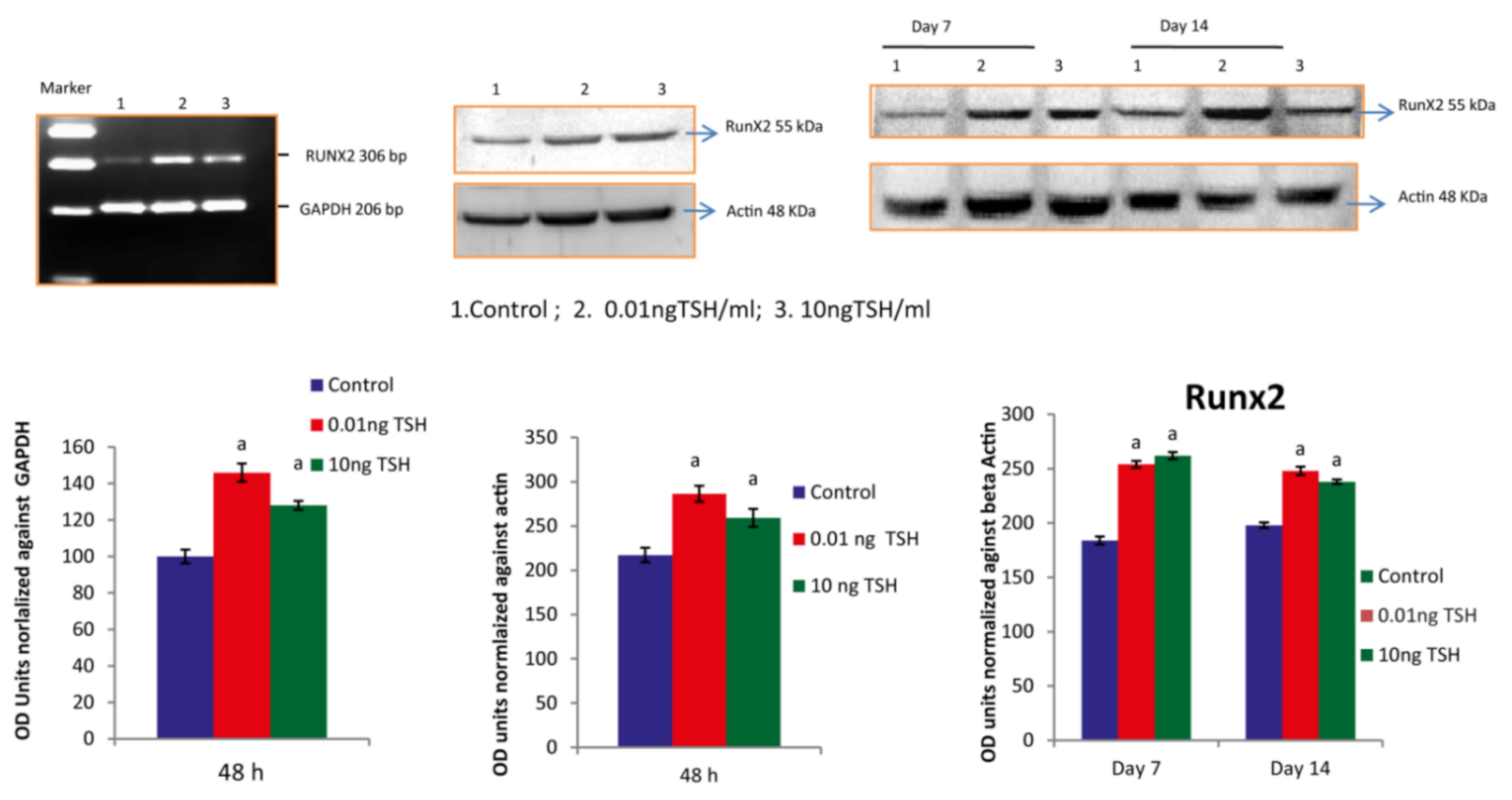
2.1. TSH Induces Runx2 mRNA and Protein Levels in HOS and SaOS2 Cells
2.2. TSH Stimulates IGF-IR in SaOS-2 Cells
2.3. TSH Promotes mRNA Expression of Differentiation Markers (ALP, COLLAGEN and OCN) in SaOS-2 Cells
2.4. Impact of TSH on Mineralization of Matric in SaOS-2 Cells
2.5. Runx2 Homology Modeling
2.6. Protein-Protein Interactions of RUNX-2 with CREB and ELK
3. Discussion
4. Materials and Methods
4.1. Culturing and Maitenance of Cells
4.2. Chemicals
4.3. Gene Expression Study
4.4. Protein Expression Study
4.5. Assessment of Mineralization of Matrix
4.6. Molecular Docking Study
4.6.1. Retrieval of Protein Structures
4.6.2. Homology Modeling of RUNX-2
4.6.3. Protein–Protein Docking
4.6.4. Visualization of Protein–Protein Interactions
4.7. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Abe, E.; Marians, R.C.; Yu, W.; Wu, X.B.; Ando, T.; Li, Y.; Iqbal, J.; Eldeiry, L.; Rajendren, G.; Blair, H.C.; et al. TSH is a negative regulator of skeletal remodeling. Cell 2003, 115, 151–162. [Google Scholar] [CrossRef] [Green Version]
- Sampath, T.K.; Simic, P.; Sendak, R.; Draca, N.; Bowe, A.E.; O’Brien, S.; Schiavi, S.C.; McPherson, J.M.; Vukicevic, S. Thyroid-stimulating hormone restores bone volume, microarchitecture, and strength in aged ovariectomized rats. J. Bone Miner. Res. 2007, 22, 849–859. [Google Scholar] [CrossRef]
- Sun, L.; Vukicevic, S.; Baliram, R.; Yang, G.; Sendak, R.; McPherson, J.; Zhu, L.L.; Iqbal, J.; Latif, R.; Natrajan, A.; et al. Intermittent recombinant TSH injections prevent ovariectomy-induced bone loss. Proc. Natl. Acad. Sci. USA 2008, 105, 4289–4294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazziotti, G.; Sorvillo, F.; Piscopo, M.; Cioffi, M.; Pilla, P.; Biondi, B.; Iorio, S.; Giustina, A.; Amato, G.; Carella, C. Recombinant human TSH modulates in vivo C-telopeptides of type-1 collagen and bone alkaline phosphatase, but not osteoprotegerin production in postmenopausal women monitored for differentiated thyroid carcinoma. J. Bone Miner. Res. 2005, 20, 480–486. [Google Scholar] [CrossRef]
- Reddi, A.H.; Sullivan, N.E. Matrix-induced endochondral bone differentiation: Influence of hypophysectomy, growth hormone, and thyroid-stimulating hormone. Endocrinology 1980, 107, 1291–1299. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Tawata, M.; Yokomori, N.; Endo, T.; Onaya, T. Expression of thyrotropin receptor on clonal osteoblast-like rat osteosarcoma cells. Thyroid 1998, 8, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.R. Does serum TSH level have thyroid hormone independent effects on bone turnover? Nat. Clin. Pract. Endocrinol. Metab. 2009, 5, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.A.; Janson, A.; Bucht, E.; Kindmark, H.; Marcus, C.; Stark, A.; Zemack, H.R.; Torring, O. Weak evidence of thyrotropin receptors in primary cultures of human osteoblast-like cells. Calcif. Tissue Int. 2004, 74, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Morimura, T.; Tsunekawa, K.; Kasahara, T.; Seki, K.; Ogiwara, T.; Mori, M.; Murakami, M. Expression of type 2 iodothyronine deiodinase in human osteoblast is stimulated by thyrotropin. Endocrinology 2005, 146, 2077–2084. [Google Scholar] [CrossRef] [Green Version]
- Ramajayam, G.; Vignesh, R.C.; Karthikeyan, S.; Kumar, K.S.; Karthikeyan, G.D.; Veni, S.; Sridhar, M.; Arunakaran, J.; Aruldhas, M.M.; Srinivasan, N. Regulation of insulin-like growth factors and their binding proteins by thyroid stimulating hormone in human osteoblast-like (SaOS2) cells. Mol. Cell. Biochem. 2012, 368, 77–88. [Google Scholar] [CrossRef]
- Boutin, A.; Eliseeva, E.; Gershengorn, M.C.; Neumann, S. β-Arrestin-1 mediates thyrotropin-enhanced osteoblast differentiation. FASEB J. 2014, 28, 3446–3455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komori, T. Regulation of osteoblast differentiation by Runx2. Adv. Exp. Med. Biol. 2010, 658, 43–49. [Google Scholar] [PubMed]
- Selvamurugan, N.; Pulumati, M.R.; Tyson, D.R.; Partridge, N.C. Parathyroid hormone regulation of the rat collagenase-3 promoter by protein kinase A-dependent transactivation of core binding factor alpha1. J. Biol. Chem. 2000, 275, 5037–5042. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, T.L.; Chang, W.Z.; Liu, Y.; Centrella, M. Runx2 integrates estrogen activity in osteoblasts. J. Biol. Chem. 2003, 278, 43121–43129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.; Xu, Y.; Fu, Q.; He, M. Parathyroid hormone regulates osteoblast differentiation in a Wnt/β-catenin-dependent manner. Mol. Cell. Biochem. 2011, 355, 211–2016. [Google Scholar] [CrossRef] [PubMed]
- Vanderschueren, D.; Vandenput, L.; Boonen, S.; Lindberg, M.K.; Bouillon, R.; Ohlsson, C. Androgens and bone. Endocr. Rev. 2004, 25, 389–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubina, E.; Lakatos, P.; Kovács, L.; Szabolcs, I.; Rácz, K.; Tóth, M.; Szucs, N.; Góth, M.I. Effects of 24 months of growth hormone (GH) treatment on serum carboxylated and undercarboxylated osteocalcin levels in GH-deficient adults. Calcif. Tissue Int. 2004, 74, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Giustina, A.; Mazziotti, G.; Canalis, E. Growth hormone, insulin-like growth factors, and the skeleton. Endocr. Rev. 2008, 29, 535–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moutsatsou, P.; Kassi, E.; Papavassiliou, A.G. Glucocorticoid receptor signaling in bone cells. Trends Mol. Med. 2012, 18, 348–359. [Google Scholar] [CrossRef]
- Varga, F.; Rumpler, M.; Zoehrer, R.; Turecek, C.; Spitzer, S.; Thaler, R.; Paschalis, E.P.; Klaushofer, K. T3 affects expression of collagen I and collagen cross-linking in bone cell cultures. Biochem. Biophys. Res. Commun. 2010, 402, 180–185. [Google Scholar] [CrossRef] [Green Version]
- Tintut, Y.; Patel, J.; Parhami, F.; Demer, L.L. Tumor necrosis factor-alpha promotes in vitro calcification of vascular cells via the cAMP pathway. Circulation 2000, 102, 2636–2642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.C.; Xie, Z.; Konaka, H.; Sodek, J.; Zhau, H.E.; Chung, L.W. Human osteocalcin and bone sialoprotein mediating osteomimicry of prostate cancer cells: Role of cAMP-dependent protein kinase A signaling pathway. Cancer Res. 2005, 65, 2303–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Hassan, M.Q.; Xie, R.L.; Hawse, J.R.; Spelsberg, T.C.; Montecino, M.; Stein, J.L.; Lian, J.B.; van Wijnen, A.J.; Stein, G.S. Co-stimulation of the bone-related Runx2 P1 promoter in mesenchymal cells by SP1 and ETS transcription factors at polymorphic purine-rich DNA sequences (Y-repeats). J. Biol. Chem. 2009, 284, 3125–3135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baliram, R.; Sun, L.; Cao, J.; Li, J.; Latif, R.; Huber, A.K.; Yuen, T.; Blair, H.C.; Zaidi, M.; Davies, T.F. Hyperthyroid-associated osteoporosis is exacerbated by the loss of TSH signaling. J. Clin. Investig. 2012, 122, 3737–3741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endo, T.; Ohta, K.; Kobayashi, T. Expression and function of Cbfa-1/Runx2 in thyroid papillary carcinoma cells. J. Clin. Endocrinol. Metab. 2008, 93, 2409–2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franceschi, R.T.; Xiao, G.; Jiang, D.; Gopalakrishnan, R.; Yang, S.; Reith, E. Multiple signaling pathways converge on the Cbfa1/Runx2 transcription factor to regulate osteoblast differentiation. Connect. Tissue Res. 2003, 44, 109–116. [Google Scholar] [CrossRef]
- Kim, B.G.; Kim, H.J.; Park, H.J.; Kim, Y.J.; Yoon, W.J.; Lee, S.J.; Ryoo, H.M.; Cho, J.Y. Runx2 phosphorylation induced by fibroblast growth factor-2/protein kinase C pathways. Proteomics 2006, 6, 1166–1174. [Google Scholar] [CrossRef] [PubMed]
- Rivas, M.; Santisteban, P. TSH-activated signaling pathways in thyroid tumorigenesis. Mol. Cell. Endocrinol. 2003, 213, 31–45. [Google Scholar] [CrossRef]
- Baliram, R.; Latif, R.; Berkowitz, J.; Frid, S.; Colaianni, G.; Sun, L.; Zaidi, M.; Davies, T.F. Thyroid-stimulating hormone induces a Wnt-dependent, feed-forward loop for osteoblastogenesis in embryonic stem cell cultures. Proc. Natl. Acad. Sci. USA 2011, 108, 16277–16282. [Google Scholar] [CrossRef] [Green Version]
- Ihn, H.; Ohnishi, K.; Tamaki, T.; LeRoy, E.C.; Trojanowska, M. Transcriptional regulation of the human alpha2(I) collagen gene. Combined action of upstream stimulatory and inhibitory cis-acting elements. J. Biol. Chem. 1996, 271, 26717–26723. [Google Scholar] [CrossRef] [Green Version]
- Pei, Y.; Meng, X.W.; Zhou, X.Y.; Xing, X.P.; Xia, W.B. Expression of core binding factor alpha1 up-regulated by IGF-I, GM-CSF, and EGF through MAPK pathway in MC3T3-E1 and C2C12 cells. Acta Pharmacol. Sin. 2003, 24, 975–984. [Google Scholar]
- Aubin, J.E.; Liu, F.; Malaval, L.; Gupta, A.K. Osteoblast and chondroblast differentiation. Bone 1995, 17, 77–83. [Google Scholar] [CrossRef]
- Lian, J.B.; Stein, G.S.; Stein, J.L.; van Wijnen, A.J. Osteocalcin gene promoter: Unlocking the secrets for regulation of osteoblast growth and differentiation. J. Cell. Biochem. 1998, 72, 62–72. [Google Scholar] [CrossRef]
- Ducy, P.; Starbuck, M.; Priemel, M.; Shen, J.; Pinero, G.; Geoffroy, V.; Amling, M.; Karsenty, G. A Cbfa1-dependent genetic pathway controls bone formation beyond embryonic development. Genes Dev. 1999, 13, 1025–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boskey, A.L. Biomineralization: Conflicts, challenges, and opportunities. J. Cell. Biochem. 1998, 72, 83–91. [Google Scholar] [CrossRef]
- UniProt Consortium. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.; Hashimoto, S.; Lotz, M.; Pritzker, K.; Goding, J.; Terkeltaub, R. Up-regulated expression of the phosphodiesterase nucleotide pyrophosphatase family member PC-1 is a marker and pathogenic factor for knee meniscal cartilage matrix calcification. Arthritis Rheum. 2001, 44, 1071–1081. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneidman-Duhovny, D.; Inbar, Y.; Nussinov, R.; Wolfson, H.J. PatchDock and SymmDock: Servers for rigid and symmetric docking. Nucleic Acids Res. 2005, 33, W363–W367. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruthamuthu, M.K.; Ganesh, I.; Ravikumar, S.; Hong, S.H. Evaluation of zraP gene expression characteristics and construction of a lead (Pb) sensing and removal system in a recombinant Escherichia coli. Biotechnol. Lett. 2015, 37, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Somasundaram, S.; Maruthamuthu, M.K.; Ganesh, I.; Eom, G.T.; Hong, S.H. Enchancement of Gamma-Aminobutyric Acid Production by Co-Localization of Neurospora crassa OR74A Glutamate Decarboxylase with Escherichia coli GABA Transporter Via Synthetic Scaffold Complex. J. Microbiol. Biotechnol. 2017, 27, 1664–1669. [Google Scholar] [CrossRef] [Green Version]
- Kannan, M.M.; Vanitha, J.; Jiang, S.; Ramachandran, S. Impact of colchicine treatment on sorghum bicolor BT × 623. Mol. Plant Breed. 2013, 4, 128–135. [Google Scholar] [CrossRef]
- Maruthamuthu, M.K.; Hong, J.; Arulsamy, K.; Somasundaram, S.; Hong, S.; Choe, W.S.; Yoo, I.K. Development of bisphenol A-removing recombinant Escherichia coli by monomeric and dimeric surface display of bisphenol A-binding peptide. Bioprocess Biosyst. Eng. 2018, 41, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Davidson, J.L.; Wang, J.; Maruthamuthu, M.K.; Dextre, A.; Pascual-Garrigos, A.; Mohan, S.; Putikam, S.V.S.; Osman, F.O.I.; McChesney, D.; Seville, J.; et al. A paper-based colorimetric molecular test for SARS-CoV-2 in saliva. Biosens. Bioelectron. X 2021, 9, 100076. [Google Scholar] [CrossRef] [PubMed]
- Maruthamuthu, M.K.; Nadarajan, S.P.; Ganesh, I.; Ravikumar, S.; Yun, H.; Yoo, I.K.; Hong, S.H. Construction of a high efficiency copper adsorption bacterial system via peptide display and its application on copper dye polluted wastewater. Bioprocess Biosyst. Eng. 2015, 38, 2077–2084. [Google Scholar] [CrossRef] [PubMed]
- Maruthamuthu, M.K.; Selvamani, V.; Nadarajan, S.P.; Yun, H.; Oh, Y.K.; Eom, G.T.; Hong, S.H. Manganese and cobalt recovery by surface display of metal binding peptide on various loops of OmpC in Escherichia coli. J. Ind. Microbiol. Biotechnol. 2018, 45, 31–41. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer | Size (bp) | Accession No. |
|---|---|---|---|---|
| ALP | ACCTCGTTGACACCTGGAAG | CCACCATCTCGGAGAGTGAC | 189 | NM_000478 |
| Collagen | GGCCCAGAAGAACTGGTAC | CGCTGTTCTTGCAGTGGTAG | 200 | NM_000088 |
| Runx2 | ACCTCGTTGACACCTGGAAG | TCTCGGTGGCTGGTAGTGA | 309 | NM_001015051 |
| IGF1R | TGGGGAATGGAGTGCTGTAT | ACGTTTGGCCTCCCTGAACG | 438 | NM_001291858.2 |
| OCN | GAATGGTCACTGGGCTGTTT | CTGGCAGCTTTGCACAATTA | 109 | NM_199173.4 |
| GAPDH | GGAGTCAACGGATTTGGT | ATCAATGGAAATCCCATCAC | 206 | NM_002046 |
| GAPDH | GCTCTCCAGAACATCATCCCT | CGTTGTCATACCAGGAAATGAGCT | 346 | NM_001289745 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Govindan, R.; El-Sherbiny, M.; Ibraheem, K.M.M.; Narasimhan, S.; Salama, M.E.-D.M.; Ahmad, F.; Jayaraman, S.; Veeraraghavan, V.P.; Vengadassalapathy, S.; Mohan, S.K.; et al. Thyroid-Stimulating Hormone Favors Runx2-Mediated Matrix Mineralization in HOS and SaOS2 Cells: An In Vitro and In Silico Approach. Molecules 2022, 27, 613. https://doi.org/10.3390/molecules27030613
Govindan R, El-Sherbiny M, Ibraheem KMM, Narasimhan S, Salama ME-DM, Ahmad F, Jayaraman S, Veeraraghavan VP, Vengadassalapathy S, Mohan SK, et al. Thyroid-Stimulating Hormone Favors Runx2-Mediated Matrix Mineralization in HOS and SaOS2 Cells: An In Vitro and In Silico Approach. Molecules. 2022; 27(3):613. https://doi.org/10.3390/molecules27030613
Chicago/Turabian StyleGovindan, Ramajayam, Mohamed El-Sherbiny, Khalid Mohamed Morsy Ibraheem, Srinivasan Narasimhan, Mohamed EL-Dosoky Mohamed Salama, Fazil Ahmad, Selvaraj Jayaraman, Vishnu Priya Veeraraghavan, Srinivasan Vengadassalapathy, Surapaneni Krishna Mohan, and et al. 2022. "Thyroid-Stimulating Hormone Favors Runx2-Mediated Matrix Mineralization in HOS and SaOS2 Cells: An In Vitro and In Silico Approach" Molecules 27, no. 3: 613. https://doi.org/10.3390/molecules27030613