
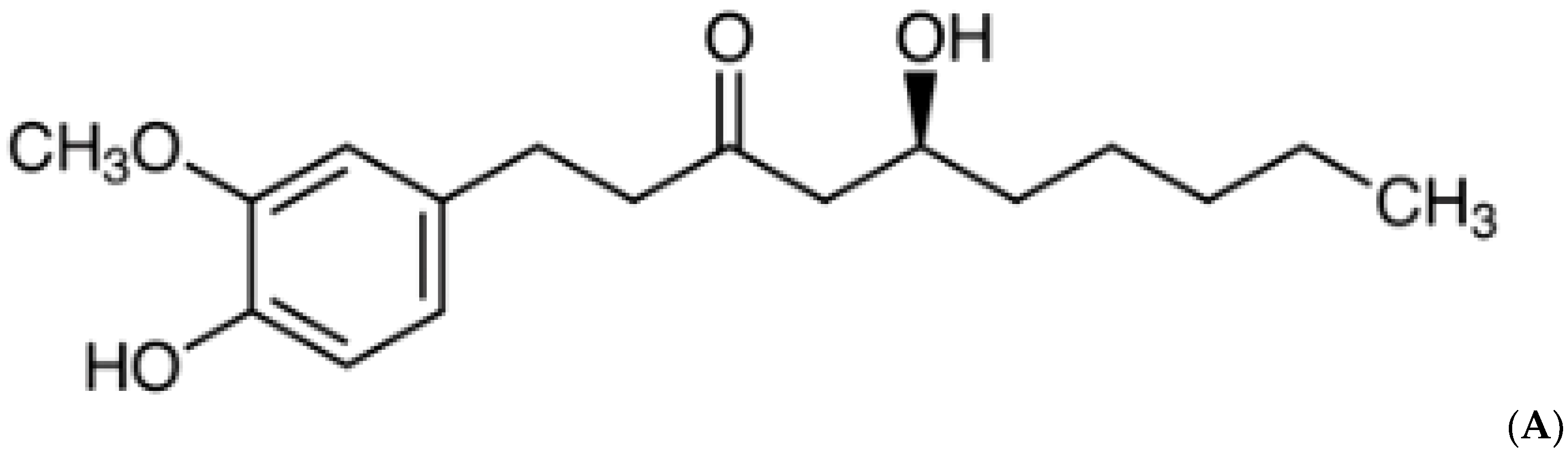
Cardioprotective Effects of 6-Gingerol against Alcohol-Induced ROS-Mediated Tissue Injury and Apoptosis in Rats
 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Results
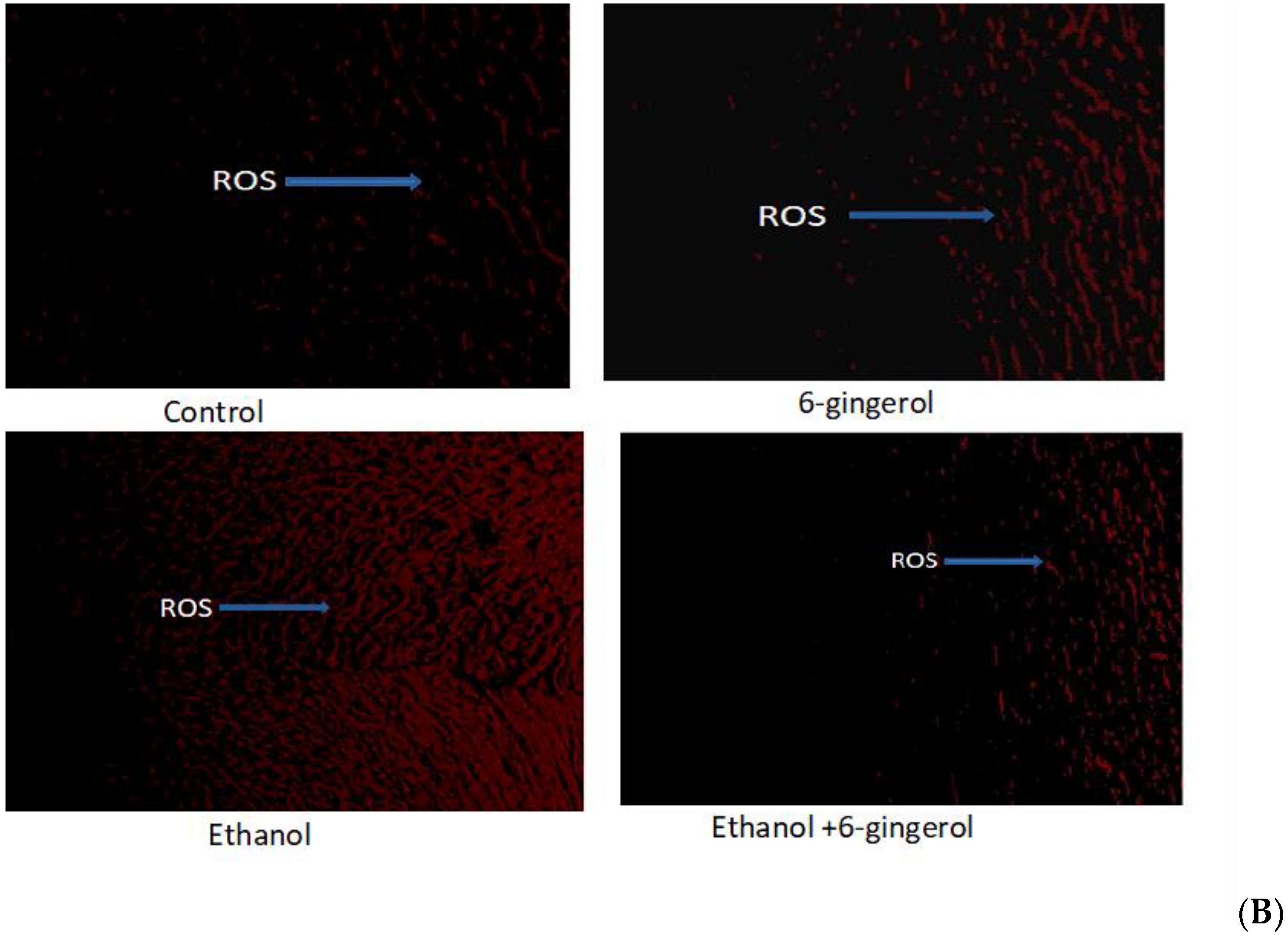
2.1. Six-Gingerol Suppress ROS Production in Alcohol-Fed Rats
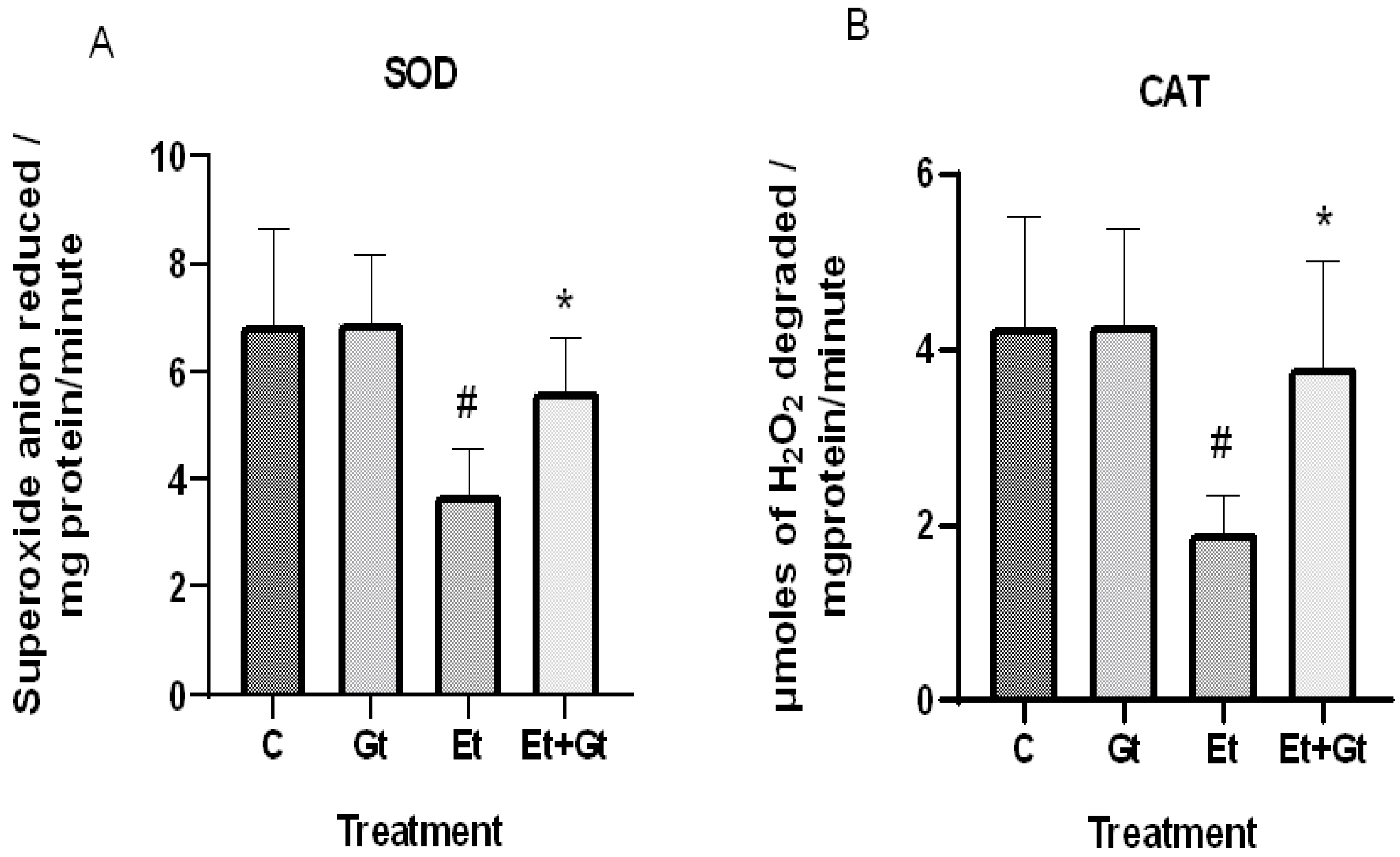
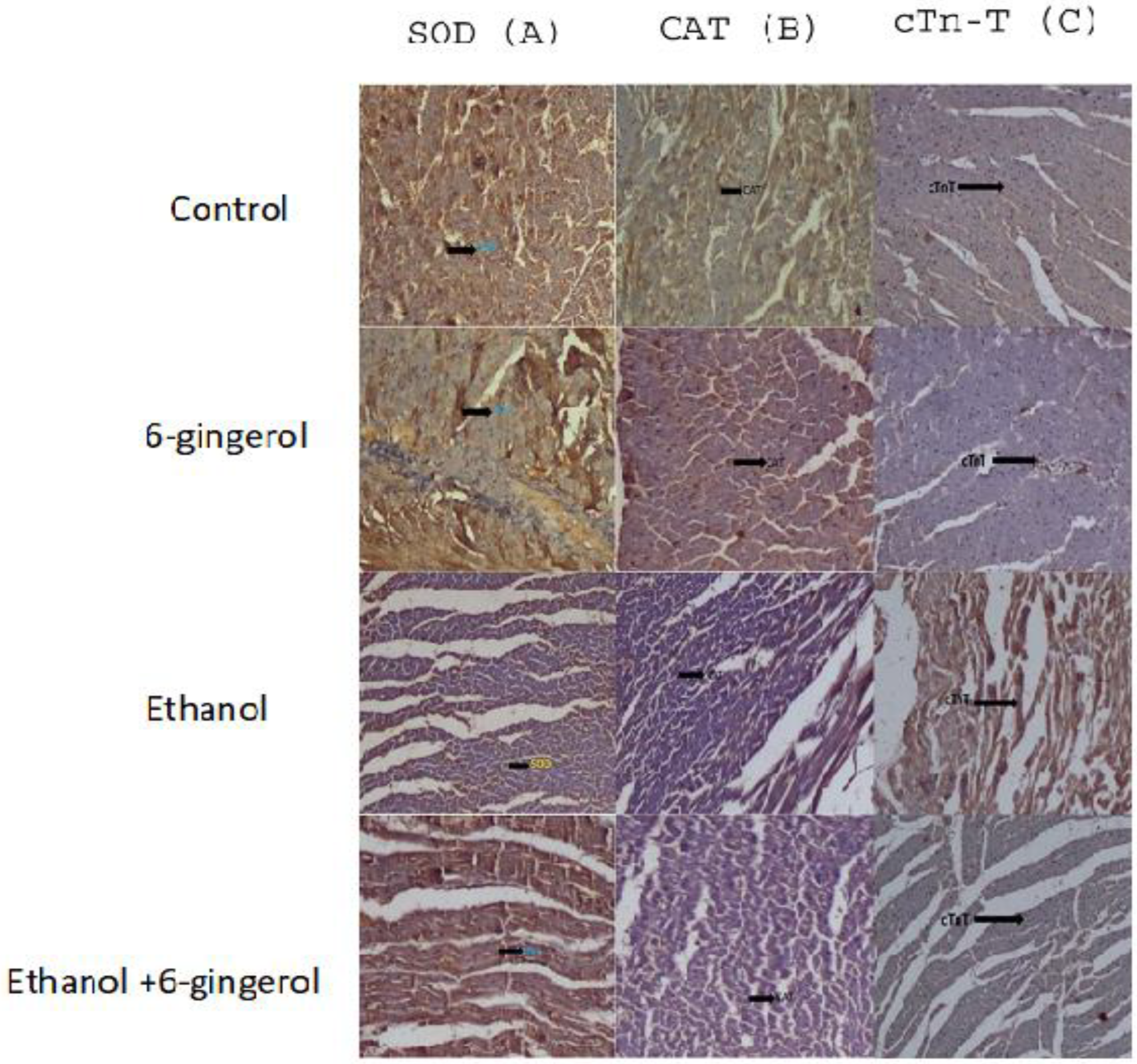
2.2. Treatment of 6-Gingerol Restores Antioxidant Enzymes against Alcohol-Induced Loss
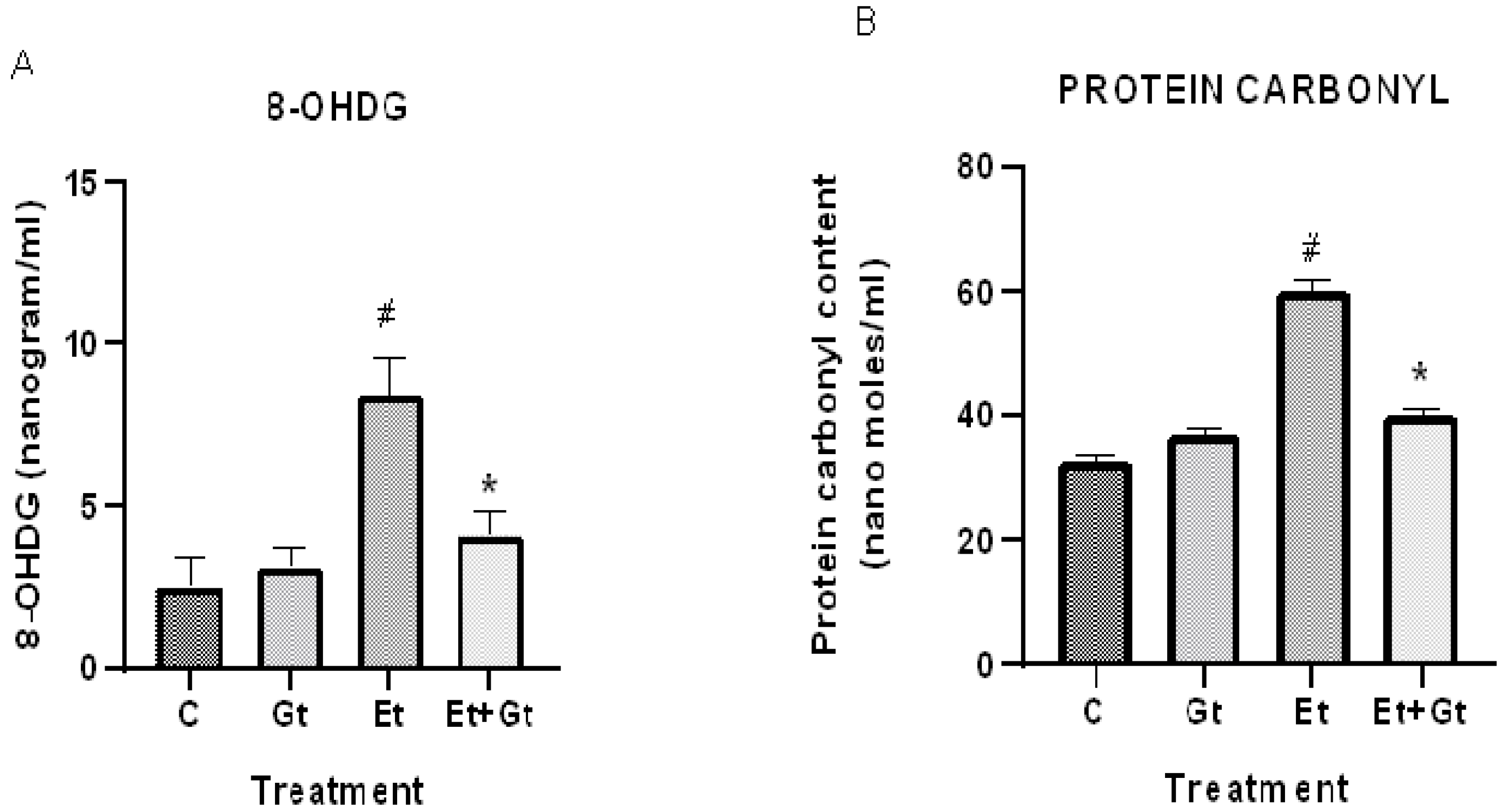
2.3. Gingerol Treatment Reverses Elevated 8-Hydroxyguanosine (8-OHDG) and Protein Carbonyls
2.4. Antilipidemic Properties of 6-Gingerol in Alcohol-Fed Rats
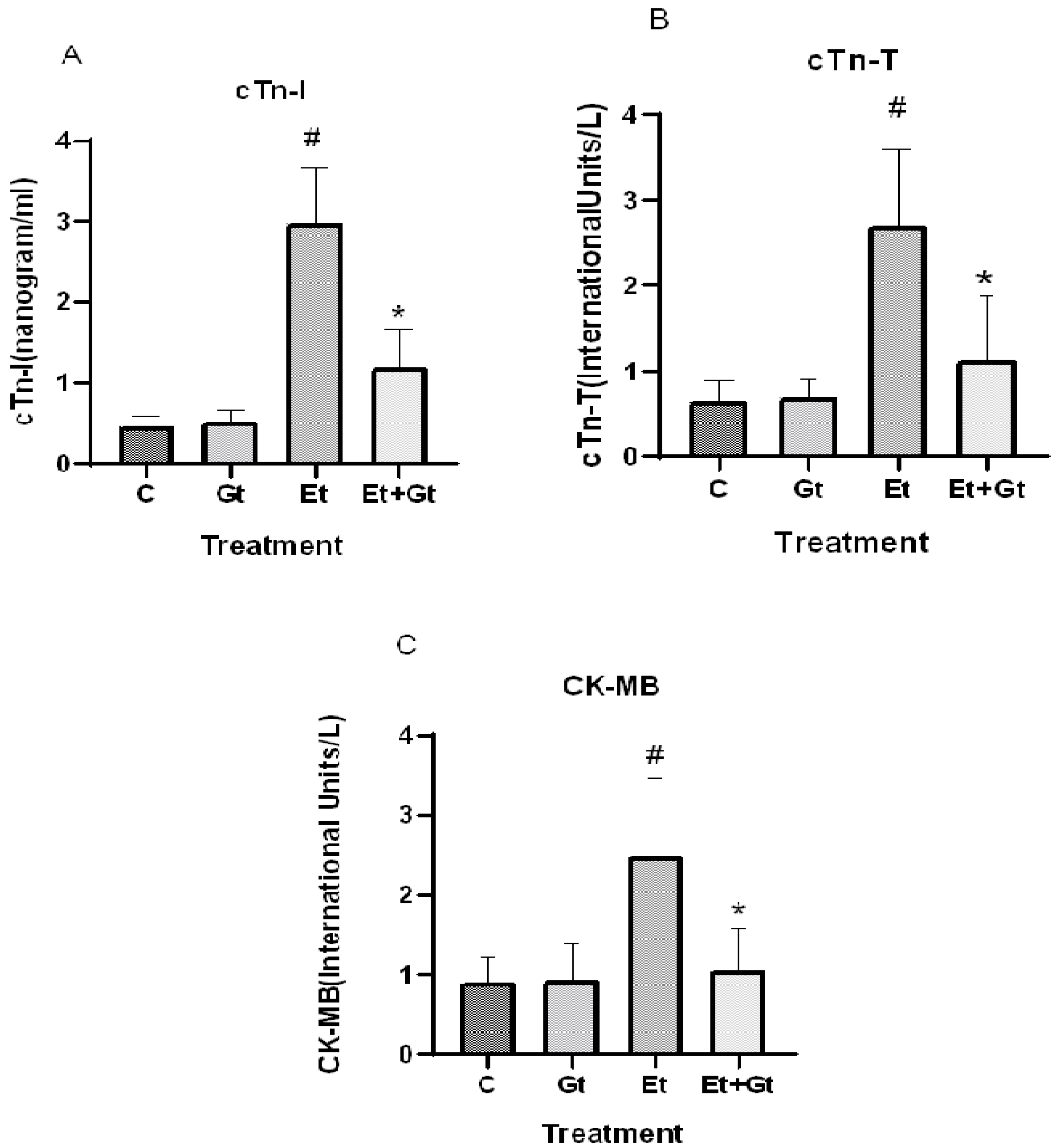
2.5. Gingerol Prevents Alcohol-Induced Elevation of Cardiac Biomarkers
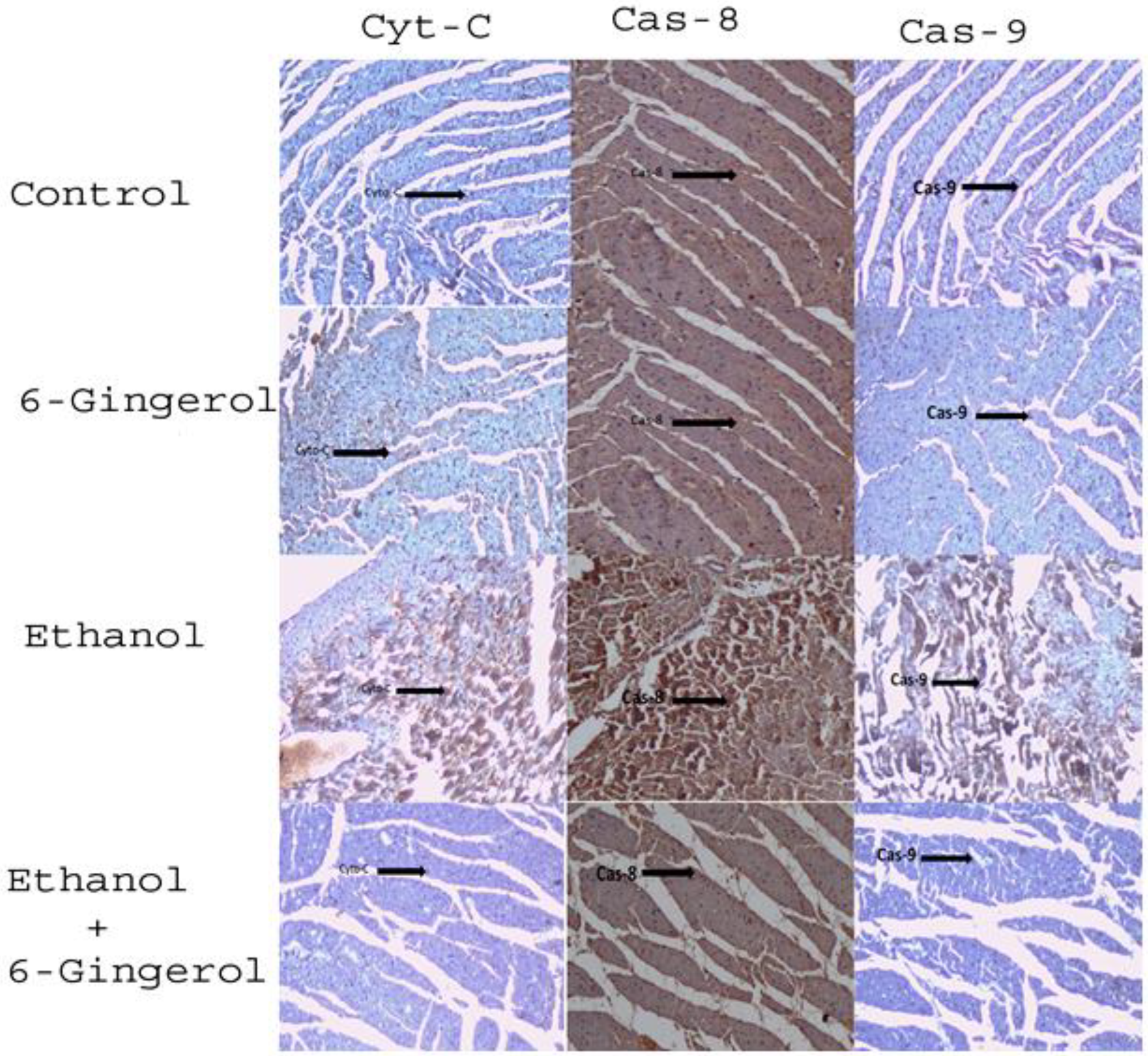
2.6. Gingerol Treatment Attenuates Upregulated Apoptotic Markers in Alcohol-Fed Cardiac Tissue
2.7. Transmission Electron Microscopy (TEM) Demonstrates Cardioprotective Effects of Gingerol
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Experimental Animals
4.3. Animal Grouping and Treatment
4.4. ROS Detection by Dihydroethidium (DHE) Fluorescent Dye
4.5. Determination of 8-Hydroxydeoxyguanosine (8-OHDG)
4.6. Estimation of Protein Carbonyl Content
4.7. Assessment of Superoxide Dismutase and Catalase Activities
4.8. Estimation Cardiovascular Risk Factors
4.9. Evaluation of Cardiac Biomarkers
4.10. Immunohistochemistry
4.11. Transmission Electron Microscopy
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Cho, L.; Davis, M.; Elgendy, I.; Epps, K.; Lindley, K.J.; Mehta, P.K.; Michos, E.D.; Minissian, M.; Pepine, C.; Vaccarino, V. Summary of updated recommendations for primary prevention of cardiovascular disease in women: JACC state-of-the-art review. J. Am. Coll. Cardiol. 2020, 75, 2602–2618. [Google Scholar] [CrossRef]
- Guidot, D.M.; Hart, M.C. Alcohol abuse and acute lung injury: Epidemiology and pathophysiology of a recently recognized association. J. Investig. Med. 2005, 53, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Mallikarjuna, K.; Shanmugam, K.R.; Nishanth, K.; Wu, M.-C.; Hou, C.-W.; Kuo, C.-H.; Reddy, K.S. Alcohol-induced deterioration in primary antioxidant and glutathione family enzymes reversed by exercise training in the liver of old rats. Alcohol 2010, 44, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Djoussé, L.; Gaziano, J.M. Alcohol consumption and heart failure: A systematic review. Curr. Atheroscler. Rep. 2008, 10, 117–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steiner, J.L.; Lang, C.H. Etiology of alcoholic cardiomyopathy: Mitochondria, oxidative stress and apoptosis. Int. J. Biochem. Cell Biol. 2017, 89, 125–135. [Google Scholar] [CrossRef]
- Dai, W.; Chen, C.; Feng, H.; Li, G.; Peng, W.; Liu, X.; Yang, J.; Hu, X. Protection of Ficus pandurata Hance against acute alcohol-induced liver damage in mice via suppressing oxidative stress, inflammation, and apoptosis. J. Ethnopharmacol. 2021, 275, 114140. [Google Scholar] [CrossRef]
- Chen, K.-N.; Peng, W.-H.; Hou, C.-W.; Chen, C.-Y.; Chen, H.-H.; Kuo, C.-H.; Korivi, M. Codonopsis javanica root extracts attenuate hyperinsulinemia and lipid peroxidation in fructose-fed insulin resistant rats. J. Food Drug Anal. 2013, 21, 347–355. [Google Scholar] [CrossRef] [Green Version]
- Ye, L.; Pan, Y.; Zheng, W.; Hu, J. miR-186-5p is Expressed Highly in Ethanol-induced Cardiomyocytes and Regulates Apoptosis by Target Gene XIAP. China Biotechnol. 2019, 39, 53–62. [Google Scholar]
- Raymond, A.R.; Becker, J.; Woodiwiss, A.J.; Booysen, H.L.; Norton, G.R.; Brooksbank, R.L. Ethanol-associated cardiomyocyte apoptosis and left ventricular dilation are unrelated to changes in myocardial telomere length in rats. J. Card. Fail. 2016, 22, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, J.; Yang, W.; Bi, Y.; Chi, J.; Tian, J.; Li, W. High-dose alcohol induces reactive oxygen species-mediated apoptosis via PKC-β/p66Shc in mouse primary cardiomyocytes. Biochem. Biophys. Res. Commun. 2015, 456, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Subbaiah, G.; Mallikarjuna, K.; Shanmugam, B.; Ravi, S.; Taj, P.; Reddy, K. Ginger treatment ameliorates alcohol-induced myocardial damage by suppression of hyperlipidemia and cardiac biomarkers in rats. Pharmacogn. Mag. 2017, 13, 69–75. [Google Scholar]
- Kumar Gupta, S.; Sharma, A. Medicinal properties of Zingiber officinale Roscoe-A review. J. Pharm. Biol. Sci. 2014, 9, 124–129. [Google Scholar]
- Wang, S.; Zhang, C.; Yang, G.; Yang, Y. Biological properties of 6-gingerol: A brief review. Nat. Prod. Commun. 2014, 9, 1027–1030. [Google Scholar] [CrossRef] [PubMed]
- Yagihashi, S.; Miura, Y.; Yagasaki, K. Inhibitory effect of gingerol on the proliferation and invasion of hepatoma cells in culture. Cytotechnology 2008, 57, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Sampath, C.; Sang, S.; Ahmedna, M. In vitro and in vivo inhibition of aldose reductase and advanced glycation end products by phloretin, epigallocatechin 3-gallate and [6]-gingerol. Biomed. Pharmacother. 2016, 84, 502–513. [Google Scholar] [CrossRef] [PubMed]
- El-Bakly, W.M.; Louka, M.L.; El-Halawany, A.M.; Schaalan, M.F. 6-gingerol ameliorated doxorubicin-induced cardiotoxicity: Role of nuclear factor kappa B and protein glycation. Cancer Chemother. Pharmacol. 2012, 70, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Liu, P.; Liu, M.; Wei, Z.; Fan, S.; Wang, X.; Sun, S.; Chu, L. [6]-Gingerol ameliorates ISO-induced myocardial fibrosis by reducing oxidative stress, inflammation, and apoptosis through inhibition of TLR4/MAPKs/NF-κB pathway. Mol. Nutr. Food Res. 2020, 64, 2000003. [Google Scholar] [CrossRef] [PubMed]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [Green Version]
- Zick, S.M.; Djuric, Z.; Ruffin, M.T.; Litzinger, A.J.; Normolle, D.P.; Alrawi, S.; Feng, M.R.; Brenner, D.E. Pharmacokinetics of 6-Gingerol, 8-Gingerol, 10-Gingerol, and 6-Shogaol and Conjugate Metabolites in Healthy Human Subjects. Cancer Epidemiol. Biomark. Prev. 2008, 17, 1930–1936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mustroph, J.; Lebek, S.; Maier, L.S.; Neef, S. Mechanisms of cardiac ethanol toxicity and novel treatment options. Pharmacol. Ther. 2019, 197, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Das, S.K.; Vasudevan, D. Alcohol-induced oxidative stress. Life Sci. 2007, 81, 177–187. [Google Scholar] [CrossRef]
- Zima, T.; Fialová, L.; Mestek, O.; Janebová, M.; Crkovská, J.; Malbohan, I.; Štípek, S.; Mikulíková, L.; Popov, P. Oxidative stress, metabolism of ethanol and alcohol-related diseases. J. Biomed. Sci. 2001, 8, 59–70. [Google Scholar] [CrossRef]
- Sato, Y.; Fujiwara, H.; Takatsu, Y. Biochemical markers in heart failure. J. Cardiol. 2012, 59, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Alsahli, M.A.; Almatroodi, S.A.; Almatroudi, A.; Khan, A.A.; Anwar, S.; Almutary, A.G.; Alrumaihi, F.; Rahmani, A.H. 6-Gingerol, a major ingredient of ginger attenuates Diethylnitrosamine-induced liver injury in rats through the modulation of oxidative stress and anti-inflammatory activity. Mediat. Inflamm. 2021, 2021, 6661937. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, F.A.P.; Prata, M.M.G.; Oliveira, I.C.M.; Alves, N.T.Q.; Freitas, R.E.M.; Monteiro, H.S.A.; Silva, J.S.A.; Vieira, P.C.; Viana, D.A.; Libório, A.B.; et al. Gingerol Fraction from Zingiber officinale Protects against Gentamicin-Induced Nephrotoxicity. Antimicrob. Agents Chemother. 2014, 58, 1872–1878. [Google Scholar] [CrossRef] [Green Version]
- Subbaiah, G.V.; Reddy, K.S.; JayavardhanaRao, Y.; Shanmugam, B.; Ravi, S.; Shanmugam, K.R.; Narasimha, G. 6-Gingerol prevents free transition metal Ion[Fe(Ii)] Induced free radicals mediated alterations by In vitro and Ndv growth in chicken eggs by In ovo. Pharmacogn. Mag. 2018, 14, 167. [Google Scholar] [CrossRef]
- Han, X.; Yang, Y.; Zhang, M.; Chu, X.; Zheng, B.; Liu, C.; Xue, Y.; Guan, S.; Sun, S.; Jia, Q. Protective Effects of 6-Gingerol on Cardiotoxicity Induced by Arsenic Trioxide Through AMPK/SIRT1/PGC-1α Signaling Pathway. Front. Pharmacol. 2022, 13, 1298. [Google Scholar] [CrossRef] [PubMed]
- Fathi, R.; Nasiri, K.; Akbari, A.; Ahmadi-KaniGolzar, F.; Farajtabar, Z. Exercise protects against ethanol-induced damage in rat heart and liver through the inhibition of apoptosis and activation of Nrf2/Keap-1/HO-1 pathway. Life Sci. 2020, 256, 117958. [Google Scholar] [CrossRef] [PubMed]
- Harishekar, M.; Kiran, B. Effect of Lead, Alcohol and Vitamin E on Protein carbonyl content in rats. J. Appl. Pharm. Sci. 2011, 9, 154–156. [Google Scholar]
- Varga, Z.V.; Ferdinandy, P.; Liaudet, L.; Pacher, P. Drug-induced mitochondrial dysfunction and cardiotoxicity. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1453–H1467. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Gao, X.; Jiang, L.; Sun, X.; Liu, X.; Chen, M.; Yao, X.; Sun, Q.; Wang, S. 6-Gingerol prevents MEHP-induced DNA damage in human umbilical vein endothelia cells. Hum. Exp. Toxicol. 2017, 36, 1177–1185. [Google Scholar] [CrossRef]
- Li, Y.; Xu, B.; Xu, M.; Chen, D.; Xiong, Y.; Lian, M.; Sun, Y.; Tang, Z.; Wang, L.; Jiang, C.; et al. 6-Gingerol protects intestinal barrier from ischemia/reperfusion-induced damage via inhibition of p38 MAPK to NF-κB signalling. Pharmacol. Res. 2017, 119, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Yon, J.-M.; Baek, I.-J.; Lee, S.-R.; Kim, M.-R.; Hong, J.T.; Yong, H.; Lee, B.J.; Yun, Y.W.; Nam, S.-Y. Protective effect of [6]-gingerol on the ethanol-induced teratogenesis of cultured mouse embryos. Arch. Pharmacal Res. 2012, 35, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.S.; Li, T.D.; Zeng, Z.H. Mechanisms underlying direct actions of hyperlipidemia on myocardium: An updated review. Lipids Health Dis. 2020, 19, 23. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, G.; Ponmurugan, P.; Deepa, M.A.; Senthilkumar, B. Anti-obesity action of gingerol: Effect on lipid profile, insulin, leptin, amylase and lipase in male obese rats induced by a high-fat diet. J. Sci. Food Agric. 2014, 94, 2972–2977. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.B.; Singh, N.; Maurya, R.; Kumar, A. Anti-hyperglycaemic, lipid lowering and anti-oxidant properties of (6)-gingerol in db/db mice. Int. J. Med. Med. Sci. 2009, 12, 536–544. [Google Scholar]
- Csengeri, D.; Sprünker, N.-A.; Di Castelnuovo, A.; Niiranen, T.; Vishram-Nielsen, J.K.; Costanzo, S.; Söderberg, S.; Jensen, S.M.; Vartiainen, E.; Donati, M.B.; et al. Alcohol consumption, cardiac biomarkers, and risk of atrial fibrillation and adverse outcomes. Eur. Heart J. 2021, 42, 1170–1177. [Google Scholar] [CrossRef]
- Luo, H.; Malik, N.; Nai, Q.; Jessani, N.; Alam, M.; Islam, M.A.; Yousif, A. Troponin I Elevation Due to Alcoholism in Absence of Acute Coronary Syndrome: A Case Report. J. Med. Cases 2014, 5, 545–548. [Google Scholar] [CrossRef] [Green Version]
- Foley, J.D.; Miller, C.S.; Sneed, J.D.; Ebersole, J.L.; Kryscio, R.J.; McDevitt, J.T.; Campbell, C.L. Serum biomarker release patterns following alcohol septal ablation for treatment of hypertrophic cardiomyopathy. Curr. Biomark. Find. 2014, 4, 161–168. [Google Scholar]
- Ren, Q.; Zhao, S.; Ren, C. 6-Gingerol protects cardiocytes H9c2 against hypoxia-induced injury by suppressing BNIP3 expression. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2016–2023. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Qin, G.; Jiang, W.; Zhao, Y.; Xu, Y.; Lv, X. 6-Gingerol Protects Heart by Suppressing Myocardial Ischemia/Reperfusion Induced Inflammation via the PI3K/Akt-Dependent Mechanism in Rats. Evid. Based Complement. Altern. Med. 2018, 2018, 6209679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajnóczky, G.; Buzas, C.J.; Pacher, P.; Hoek, J.B.; Rubin, E. Alcohol and mitochondria in cardiac apoptosis: Mechanisms and visualization. Alcohol. Clin. Exp. Res. 2005, 29, 693–701. [Google Scholar] [CrossRef]
- Del Re, D.P.; Miyamoto, S.; Brown, J.H. RhoA/Rho kinase up-regulate Bax to activate a mitochondrial death pathway and induce cardiomyocyte apoptosis. J. Biol. Chem. 2007, 282, 8069–8078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starkov, A.A. The role of mitochondria in reactive oxygen species metabolism and signaling. Ann. N. Y. Acad. Sci. 2008, 1147, 37–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullen, S.; Martin, S. Caspase activation pathways: Some recent progress. Cell Death Differ. 2009, 16, 935–938. [Google Scholar] [CrossRef]
- Chen, D.B.; Wang, L.; Wang, P.H. Insulin-like growth factor I retards apoptotic signaling induced by ethanol in cardiomyocytes. Life Sci. 2000, 67, 1683–1693. [Google Scholar] [CrossRef]
- Rajbanshi, S.L.; Pandanaboina, C.S. Alcohol stress on cardiac tissue–Ameliorative effects of Thespesia populnea leaf extract. J. Cardiol. 2014, 63, 449–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakazawa, T.; Ohsawa, K. Metabolism of [6]-gingerol in rats. Life Sci. 2002, 70, 2165–2175. [Google Scholar] [CrossRef]
- Srinivasan, S.; Dubey, K.K.; Singhal, R.S. Influence of food commodities on hangover based on alcohol dehydrogenase and aldehyde dehydrogenase activities. Curr. Res. Food Sci. 2019, 1, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Cutler, R.G. Human longevity and aging: Possible role of reactive oxygen species. Ann. N. Y. Acad. Sci. 1991, 621, 1859082. [Google Scholar] [CrossRef] [PubMed]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. [13] Catalase In Vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Liu, H.; Lin, F. Application of immunohistochemistry in thyroid pathology. Arch. Pathol. Lab. Med. 2015, 139, 67–82. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, I.-S.; Yamada, E. The fine structure of lamellated nerve endings found in the rat gingiva. Arch. Histol. Jpn. 1983, 46, 173–182. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid Profile (mg/dL) | Groups | |||
|---|---|---|---|---|
| C | Gt | Et | Et + Gt | |
| TG | 38.83 ± 5.05 | 39.38 ± 4.17 | 89.20 ± 6.08 # | 44.17 ± 4.98 * |
| TC | 66.94 ± 4.12 | 64.24 ± 5.13 | 119.86 ± 5.45 # | 72.45 ± 6.13 * |
| HDL | 25.13 ± 2.36 | 24.51 ± 2.08 | 12.62 ± 3.06 # | 21.33 ± 1.94 * |
| LDL | 40.48 ± 2.11 | 38.73 ± 1.91 | 109.11 ± 2.37 # | 56.77 ± 1.98 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ganjikunta, V.S.; Maddula, R.R.; Bhasha, S.; Sahukari, R.; Kondeti Ramudu, S.; Chenji, V.; Kesireddy, S.R.; Zheng, Z.; Korivi, M. Cardioprotective Effects of 6-Gingerol against Alcohol-Induced ROS-Mediated Tissue Injury and Apoptosis in Rats. Molecules 2022, 27, 8606. https://doi.org/10.3390/molecules27238606
Ganjikunta VS, Maddula RR, Bhasha S, Sahukari R, Kondeti Ramudu S, Chenji V, Kesireddy SR, Zheng Z, Korivi M. Cardioprotective Effects of 6-Gingerol against Alcohol-Induced ROS-Mediated Tissue Injury and Apoptosis in Rats. Molecules. 2022; 27(23):8606. https://doi.org/10.3390/molecules27238606
Chicago/Turabian StyleGanjikunta, Venkata Subbaiah, Ramana Reddy Maddula, Shanmugam Bhasha, Ravi Sahukari, Shanmugam Kondeti Ramudu, Venkatrayulu Chenji, Sathyavelu Reddy Kesireddy, Zhe Zheng, and Mallikarjuna Korivi. 2022. "Cardioprotective Effects of 6-Gingerol against Alcohol-Induced ROS-Mediated Tissue Injury and Apoptosis in Rats" Molecules 27, no. 23: 8606. https://doi.org/10.3390/molecules27238606