
Indonesian Mangrove Sonneratia caseolaris Leaves Ethanol Extract Is a Potential Super Antioxidant and Anti Methicillin-Resistant Staphylococcus aureus Drug
 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Leaves Simplicia Extraction and Solvent Determination
2.2. Quantitative and Qualitative Phytochemical Analysis
2.2.1. Total Phenolic Content (TPC) Determination
2.2.2. Flavonoid Content (FC) Determination
2.3. Antioxidant Activity Determination by DPPH Radical Scavenging Assay
2.4. Brine Shrimp Lethality Test (BSLT) for Extract Toxicity Assay
2.5. Agar Disk-Diffusion Assay for Antibacterial Activity Determination against MRSA Strain
2.6. Liquid Chromatography-Mass Spectroscopy (LC-MS) for Extract Compound Characterization
2.7. Statistical Analysis
3. Results and Discussion
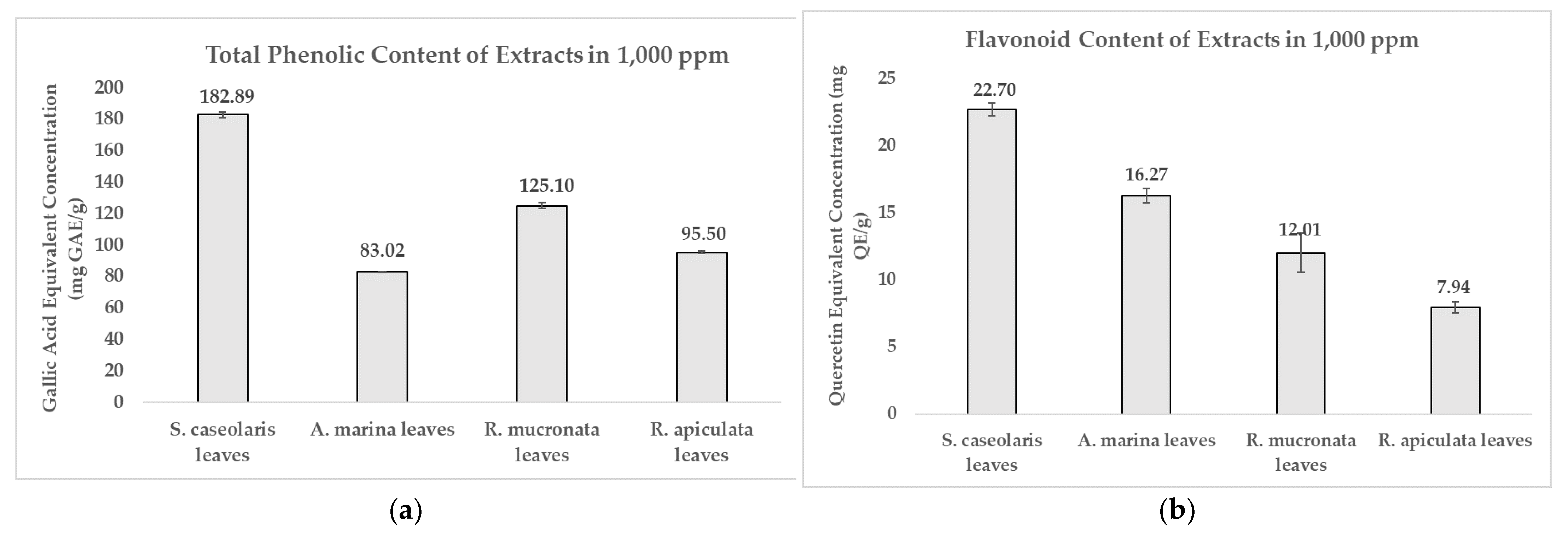
3.1. Dried Leaves Simplicial from Local Mangrove Species Phytochemical Extraction and Characterization
3.2. Toxicity of Various Local Mangrove Species Leaves Extracts
3.3. Antioxidant Activity of Phytochemicals in Various Local Mangrove Species Leaves Extract
3.4. Antibacterial Activity of Various Local Mangrove Species Leaves Extract against MRSA Strain
3.5. Phytochemical Compound Characterization by Liquid Chromatography-Mass Spectrometry (LC-MS) for Ethanol-Based S. caseolaris Leaves Extract Biological Activity-Compound Determination
3.6. Relationship between Antioxidant and Antibacterial Activity from a Phytochemical Standpoint
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Kapoor, G.; Saigal, S.; Elongavan, A. Action and Resistance Mechanisms of Antibiotics: A Guide for Clinicians Basic Anatomy of Bacterial Cell. J. Anaesthesiol. Clin. Pharmacol. 2017, 33, 300–305. [Google Scholar] [CrossRef]
- Levy, S.B.; Marshall, B. Antibacterial Resistance Worldwide: Causes, Challenges and Responses. Nat. Med. 2004, 10, 122–129. [Google Scholar] [CrossRef]
- Ventola, C.L. The Antibiotic Resistance Crisis Part 1: Causes and Threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Richardson, L.A. Understanding and overcoming antibiotic resistance. PLoS Biol. 2017, 15, e2003775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rayamajhi, N.; Cha, S.B.; Yoo, H.S. Antibiotics Resistances: Past, Present and Future. J. Biomed. Res. 2010, 11, 65–80. [Google Scholar]
- Schniederberend, M.; Abdurachim, K.; Murray, S.; Kazmierczak, B.I. The GTPase Activity of FlhF Is Dispensable for Flagellar Localization, but Not Motility, in Pseudomonas Aeruginosa. J. Bacteriol. 2013, 195, 1051–1060. [Google Scholar] [CrossRef] [Green Version]
- Turner, N.A.; Sharma-kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G. Methicillin-Resistant Staphylococcus aureus: An Overview of Basic and Clinical Research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bone, K.; Mills, S. Principles of Herbal Pharmacology. In Principles and Practice of Phytotherapy: Modern Herbal Medicine, 2nd ed.; Churchill Livingstone: London, UK, 2013; pp. 17–82. ISBN 9780443069925. [Google Scholar]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef] [PubMed]
- Reddy, M.K.; Gupta, S.K.; Jacob, M.R.; Khan, S.I.; Ferreira, D. Antioxidant, Antimalarial and Antimicrobial Activities of Tannin-Rich Fractions, Ellagitannins and Phenolic Acids from Punica granatum L. Planta Med. 2007, 73, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Amsyir, J.; Batubara, R.R.; Jogja, E.G.; Batubara, I.; Audah, K.A.; Nunuk, K.N. Introduction of Bioprospecting Opportunities for Indonesian Mangrove Species. In Proceedings of the International Biotechnology Conference on Estate Crops, Jakarta, Indonesia, 18–20 October 2017; pp. 1–5. [Google Scholar]
- Polidoro, B.A.; Carpenter, K.E.; Collins, L.; Duke, N.C.; Ellison, A.M.; Ellison, J.C.; Farnsworth, E.J.; Fernando, E.S.; Kathiresan, K.; Koedam, N.E.; et al. The Loss of Species: Mangrove Extinction Risk and Geographic Areas of Global Concern. PLoS ONE 2010, 5, e10095. [Google Scholar] [CrossRef]
- Genilar, L.A.; Kurniawaty, E.; Mokhtar, R.A.M.; Audah, K.A. Mangroves and Their Medicinal Benefit: A Mini Review. Ann. Rom. Soc. Cell Biol. 2021, 25, 695–709. [Google Scholar]
- Mouafi, F.E.; Abdel-aziz, S.M.; Bashir, A.A.; Fyiad, A.A. Phytochemical Analysis and Antimicrobial Activity of Mangrove Leaves (Avicenna Marina and Rhizophora Stylosa) Against Some Pathogens. World Appl. Sci. J. 2014, 29, 547–554. [Google Scholar] [CrossRef]
- Audah, K.A.; Batubara, R.; Amsyir, J.; Wijaya, E.; Kurniawaty, E.; Batubara, I. Antibacterial Screening of Mangrove Extract Library Showed Potential Activity against Escherichia coli and Staphylococcus aureus Antibacterial Screening of Mangrove Extract Library Showed Potential Activity against Escherichia coli and Staphylococcus aureus. J. Trop. Life Sci. 2020, 10, 105–111. [Google Scholar] [CrossRef]
- Darmadi, J.; Batubara, R.R.; Himawan, S.; Azizah, N.N.; Audah, H.K.; Arsianti, A.; Kurniawaty, E.; Ismail, I.S.; Batubara, I.; Audah, K.A. Evaluation of Indonesian Mangrove Xylocarpus Granatum Leaves Ethyl Acetate Extract as Potential Anticancer Drug. Sci. Rep. 2021, 11, 6080. [Google Scholar] [CrossRef] [PubMed]
- Jadid, N.; Hidayati, D.; Hartanti, S.R.; Arraniry, B.A.; Rachman, R.Y.; Wikanta, W. Antioxidant Activities of Different Solvent Extracts of Piper Retrofractum Vahl. Using DPPH Assay. AIP Conf. Proc. 2017, 1854, 020019. [Google Scholar]
- Dosumu, O.; Oluwaniyi, O.O.; Oyedeji, O.O. Phytochemical Screening and Brine Shrimp Assay Investigation of Vegetables Commonly Consumed in Southern and North Central Parts of Nigeria. Centrepoint J. 2017, 19, 79–88. [Google Scholar]
- Hudzicki, J. Kirby-Bauer Disk Diffusion Susceptibility Test Protocol; Protocols, American Society for Microbiology: Washington, DC, USA, 2009. [Google Scholar]
- Baris, Ö.; Güllüce, M.; Fikrettin, S.; Özer, H.; Kiliç, H.; Özkan, H.; Sökmen, M.; Özbek, T. Biological Activities of the Essential Oil and Methanol Extract of Achillea Biebersteinii Afan. (Asteraceae). Turk. J. Biol. 2006, 30, 65–73. [Google Scholar]
- Meyer, B.N.; Ferrigni, N.R.; Jacobsen, L.B.; Nichols, D.E.; McLaughin, J.L. Brine Shrimp: A Convenient General Bioassay for Active Plant Constituents. Planta Med. 2014, 45, 31–34. [Google Scholar] [CrossRef]
- Widyawati, P.S.; Budianta, T.D.W.; Kusuma, F.A.; Wijaya, E.L. Difference of Solvent Polarity To Phytochemical Content and Antioxidant Activity of Pluchea Indicia Less Leaves Extracts. Int. J. Pharmacogn. Phytochem. Res. 2014, 6, 850–855. [Google Scholar]
- Jadid, N.; Hartanti, S.R.; Abdulgani, N.; Wikanta, W.; Sulthoni, F.R. In Vitro Antioxidant Activity of Methanolic Extract of Piper Retrofractum Vahl. In Proceedings of the 5th Internatoinal Seminar on New Paradigm and Innovation on Natural Sciences and Its Application (5th ISNPINSA), Semarang, Indonesia, 7–8 October 2015; pp. 127–131. [Google Scholar]
- Tiwari, P.; Kumar, B.; Kaur, M.; Kaur, G.; Kaur, H. Phytochemical Screening and Extraction: A Review. Int. Pharm. Sci. 2011, 1, 98–106. [Google Scholar]
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; He, X.; Zhao, D. Factors Affecting Phytochemical Stability. In Handbook of Plant Food Phytochemicals: Sources, Stability and Extraction, First Edition; Tiwari, B.K., Brunton, N.P., Brennan, C.S., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 332–374. [Google Scholar]
- Taghvaei, M.; Jafari, S.M. Application and Stability of Natural Antioxidants in Edible Oils in Order to Substitute Synthetic Additives. J. Food Sci. Technol. 2015, 52, 1272–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audah, K.A. Drug Discovery: A Biodiversity Perspective. In Nanotechnology: Applications in Energy, Drug and Food; Siddiquee, S., Melvin, G.J.H., Rahman, M., Eds.; Springer Nature Switzerland AG: Cham, Switzerland, 2019; pp. 249–265. ISBN 978-3-319-99602-8. [Google Scholar]
- Yami, A.C.L.; Batubara, I.; Audah, K.A. Antioxidant and antibacterial activity of mangrove Bruguiera gymnorrhiza stem extracts against pathogenic bacteria Vibrio Cholerae. Acta Biochim. Indones. 2020, 3, 53–61. [Google Scholar] [CrossRef]
- Geethaa, S.; Thavamany, P.J.; Chiew, S.P.; Thong, O.M. Interference from Ordinarily Used Solvents in the Outcomes of Artemia salina Lethality Test. J. Adv. Pharm. Technol. Res. 2013, 4, 179–182. [Google Scholar] [CrossRef]
- Wu, C. An Important Player in Brine Shrimp Lethality Bioassay: The Solvent. J. Adv. Pharm. Technol. Res. 2014, 5, 57–58. [Google Scholar] [PubMed]
- Tanjung, I.B.; Azizah, N.N.; Arsianti, A.; Anisa, A.S.; Audah, K.A. Evaluation of the Ethyl Acetate Extract of the Roots of Avicennia marina as Potential Anticancer Drug. Adv. Biol. Sci. Res. 2022, 16, 75–81. [Google Scholar] [CrossRef]
- Pavlovic, V.; Cekic, S.; Rankovic, G.; Stoiljkovic, N. Antioxidant and pro-oxidant effect of ascorbic acid. Acta Med. Median. 2005, 44, 65–68. [Google Scholar]
- Phongpaichit, S.; Nikom, J.; Rungjindamai, N.; Sakayaroj, J.; Hutadilok-Towatana, N.; Rukachaisirikul, V.; Kirtikara, K. Biological Activities of Extracts from Endophytic Fungi Isolated from Garcinia Plants. FEMS Immunol. Med. Microbiol. 2007, 51, 517–525. [Google Scholar] [CrossRef]
- Wang, T.; Li, Q.; Bi, K. Bioactive Flavonoids in Medicinal Plants: Structure, Activity and Biological Fate. Asian J. Pharm. Sci. 2018, 13, 12–23. [Google Scholar] [CrossRef]
- Sieniawska, E.; Baj, T. Tannins. In Pharmacognosy; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. 199–232. ISBN 9780128021040. [Google Scholar]
- Sieniawska, E. Activities of Tannins—From In Vitro Studies to Clinical Trials. Nat. Prod. Commun. 2015, 10, 1877–1884. [Google Scholar] [CrossRef] [Green Version]
- Banjarnahor, S.D.S.; Artanti, N. Antioxidant Properties of Flavonoids. Med. J. Indones. 2014, 23, 239–244. [Google Scholar] [CrossRef] [Green Version]
- Isah, T. Stress and Defense Responses in Plant Secondary Metabolites Production. Biol. Res. 2019, 52, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patra, J.K.; Thatoi, H.N. Metabolic Diversity and Bioactivity Screening of Mangrove Plants: A Review. Acta Physiol. Plant. 2011, 33, 1051–1061. [Google Scholar] [CrossRef]
- Reef, R.; Lovelock, C.E. Regulation of Water Balance in Mangroves. Ann. Bot. 2015, 115, 385–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patra, J.K.; Mohanta, Y.K. Antimicrobial Compounds from Mangrove Plants: A Pharmaceutical Prospective. Chin. J. Integr. Tradit. West. Med. 2014, 20, 311–320. [Google Scholar] [CrossRef]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint tables for interpretation of MICs and zone of diameters. Version 12.0. 2022. Available online: http://www.eucast.org (accessed on 12 February 2022).
- Simlai, A.; Rai, A.; Mishra, S.; Mukherjee, K.; Roy, A. Antimicrobial and antioxidative activities in the bark extracts of Sonneratia caseolaris, a mangrove plant. Excli J. 2014, 13, 997–1010. [Google Scholar]
- Yompakdee, C.; Thunyaharn, S.; Phaechamud, T. Bactericidal Activity of Methanol Extracts of Crabapple Mangrove Tree (Sonneratia caseolaris Linn.) Against Multi-Drug Resistant Pathogens. Indian J. Pharm. Sci. 2012, 74, 230–236. [Google Scholar]
- Bartnik, M.; Facey, P.C. Glycosides. In Pharmacognosy; Badal, S., Delgoda, R., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. 101–162. ISBN 9780128021040. [Google Scholar]
- Xiao, J.; Capanoglu, E.; Jassbi, A.R.; Miron, A. Advance on the Flavonoid C-Glycosides and Health Benefits. Crit. Rev. Food Sci. Nutr. 2016, 56, S29–S45. [Google Scholar] [CrossRef]
- Benatti, P.; Peluso, G.; Nicolai, R.; Calvani, M. Polyunsaturated Fatty Acids: Biochemical, Nutritional and Epigenetic Properties. J. Am. Coll. Nutr. 2004, 23, 281–302. [Google Scholar] [CrossRef]
- Bisht, A.; Hemrajani, C.; Rathore, C.; Dhiman, T.; Rolta, R.; Upadhyay, N.; Nidhi, P.; Gupta, G.; Dua, K.; Chellappan, D.K.; et al. Hydrogel Composite Containing Azelaic Acid and Tea Tree Essential Oil as a Therapeutic Strategy for Propionibacterium and Testosterone-induced Acne. Drug Deliv. Transl. Res. 2021, 12, 2501–2517. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.W.L.; Yee, Z.Y.; Raja, I.; Yap, J.K.Y. Synergistic Effect of Non-Steroidal Anti-Inflammatory Drugs (NSAIDs) on Antibacterial Activity of Cefuroxime and Chloramphenicol against Methicillin—Resistant Staphylococcus aureus. J. Glob. Antimicrob. Resist. 2017, 10, 70–74. [Google Scholar] [CrossRef]
- Mu, D.; Xiang, H.; Dong, H.; Wang, D.; Wang, T. Isovitexin, a Potential Candidate Inhibitor of Sortase A of Staphylococcus aureus USA300. J. Microbiol. Biotechnol. 2018, 28, 1426–1432. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Chen, F.; Bi, C.; Wang, L.; Zhong, X.; Cai, H.; Deng, X.; Niu, X.; Wang, D. Quercitrin, an Inhibitor of Sortase A, Interferes with the Adhesion of Staphylococcal aureus. Molecules 2015, 20, 6533–6543. [Google Scholar] [CrossRef] [PubMed]
- Gertsch, J. Botanical Drugs, Synergy, and Network Pharmacology: Forth and Back to Intelligent Mixtures. Planta Med. 2011, 77, 1086–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carocho, M.; Ferreira, I.C.F.R. A Review on Antioxidants, Prooxidants and Related Controversy: Natural and Synthetic Compounds, Screening and Analysis Methodologies and Future Perspectives. Food Chem. Toxicol. 2013, 51, 15–25. [Google Scholar] [CrossRef]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative Stress, Prooxidants, and Antioxidants: The Interplay. Biomed. Res. Int. 2014, 2014, 761264. [Google Scholar] [CrossRef] [Green Version]
- Dumbrava, D.; Popescu, L.A.; Soica, C.M.; Nicolin, A.; Cocan, I.; Negrea, M.; Alexa, E.; Obistioiu, D.; Radulov, I.; Popescu, S.; et al. Nutritional, Antioxidant, Antimicrobial, and Toxicological Profile of Two Innovative Types of Vegan, Sugar-Free Chocolate. Foods 2020, 9, 1844. [Google Scholar] [CrossRef]
- Utakod, N.; Laosripaiboon, W.; Chunhachart, O.; Issakul, K. The Efficiency and the Correlation between Testing Methods on Antimicrobial and Antioxidant Activities of Selected Medicinal Essential Oils. Int. Food Res. J. 2017, 24, 2616–2624. [Google Scholar]
- Okwu, M.U.; Olley, M.; Akpoka, A.O.; Izevbuwa, O.E. Methicillin-Resistant Staphylococcus aureus (MRSA) and Anti-MRSA Activities of Extracts of Some Medicinal Plants: A Brief Review. AIMS Microbiol. 2019, 5, 117–137. [Google Scholar] [CrossRef]
- Turnridge, J.; Rao, N.; Chang, F.-Y.; Fowler, V.G., Jr.; Kellie, S.M.; Arnold, S.; Lee, B.Y.; Tristan, A. Staphylococcus aureus. Available online: www.antimicrobe.org/sample_staphylococcus.asp/ (accessed on 4 January 2022).
- De Freitas Araújo, M.G.; Hilário, F.; Vilegas, W.; Santos, L.C.D.; Brunetti, I.L.; Sotomayor, C.E.; Bauab, T.M. Correlation among Antioxidant, Antimicrobial, Hemolytic, and Antiproliferative Properties of Leiothrix Spiralis Leaves Extract. Int. J. Mol. Sci. 2012, 13, 9260–9277. [Google Scholar] [CrossRef]
- Klingel, T.; Hadamjetz, M.; Fischer, A.; Wefers, D. Glucosylation of Flavonoids and Flavonoid Glycosies by Mutant Dextransucrase from Lactobacillus Reuteri TMW 1.106. Carbohydr. Res. 2019, 483, 107741. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, E.R.; do Carmo Maquiaveli, C.; Magalhães, P.P. The Leishmanicidal Flavonols Quercetin and Quercitrin Target Leishmania (Leishmania) Amazonensis Arginase. Exp. Parasitol. 2012, 130, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Khameneh, B.; Iranshahy, M.; Soheili, V.; Bazzaz, B.S.F. Review on Plant Antimicrobials: A Mechanistic Viewpoint. Antimicrob. Resist. Infect. Control. 2019, 8, 118. [Google Scholar] [CrossRef] [Green Version]
- Naqvi, S.A.R.; Nadeem, S.; Komal, S.; Naqvi, S.A.A.; Mubarik, M.S.; Qureshi, S.Y.; Ahmad, S.; Abbas, A.; Zahid, M.; Khan, N.-U.-H.; et al. Antioxidants: Natural Antibiotics. In Antioxidants; Shalaby, E., Brzozowski, T., Eds.; IntechOpen: London, UK, 2019. [Google Scholar]
- Stacewicz-Sapuntzakis, M.; Bowen, P.E.; Hussain, E.A.; Bernadette, I.; Farnsworth, N.R. Critical Reviews in Food Science and Nutrition Chemical Composition and Potential Health Effects of Prunes: A Functional Food? Crit. Rev. Food Sci. Nutr. 2010, 41, 251–286. [Google Scholar] [CrossRef]
- Faraji, H.; Lindsay, R.C. Characterization of the Antioxidant Activity of Sugars and Polyhydric Alcohols in Fish Oil Emulsions. J. Agric. Food Chem. 2004, 52, 7164–7171. [Google Scholar] [CrossRef]
- Ming, X.; Wang, Y.; Sui, Y. Pretreatment of the Antagonistic Yeast, Debaryomyces Hansenii, With Mannitol and Sorbitol Improves Stress Tolerance and Biocontrol Efficacy. Front. Microbiol. 2020, 11, 601. [Google Scholar] [CrossRef] [Green Version]
- Blaskovich, M.A.T.; Elliott, A.G.; Kavanagh, A.M.; Ramu, S.; Cooper, M.A. In Vitro Antimicrobial Activity of Acne Drugs Against Skin-Associated Bacteria. Sci. Rep. 2019, 9, 14658. [Google Scholar] [CrossRef] [Green Version]
- Maple, P.A.C.; Hamilton-Miller, J.M.T.; Brranfltt, W. Comparison of the In-Vitro Activities of the Topical Antimicrobials Azelaic Acid, Nitroforazone, Silver Sulphadiazine and Mupirocin against Methicillin-Resistant Staphylococcus aureus. J. Antimicrob. Chemother. 1992, 29, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Sieber, M.A.; Hegel, J.K.E. Azelaic Acid: Properties and Mode of Action. Skin Pharmacol. Physiol. 2014, 27, 9–17. [Google Scholar] [CrossRef]
- Hannachi, N.; Habib, G.; Camoin-Jau, L. Aspirin Effect on Staphylococcus aureus—Platelet Interactions During Infectious Endocarditis. Front. Med. 2019, 6, 217. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.P.; Chien, J.C.; Hsiang, W.H.; Yang, S.W.; Chen, C.Y. Aspirin Administration Might Accelerate the Subsidence of Periprosthetic Joint Infection. Sci. Rep. 2020, 10, 15967. [Google Scholar] [CrossRef] [PubMed]
- Badhani, B.; Sharma, N.; Kakkar, R. Gallic Acid: A Versatile Antioxidant with Promising Therapeutic and Industrial Applications. RSC Adv. 2015, 5, 27540–27557. [Google Scholar] [CrossRef]
- Pu, S.; Ma, Z.; Wang, Q. Anti-Staphylococcus aureus Evaluation of Gallic Acid by Isothermal Microcalorimetry and Principle Component Analysis. J. Therm. Anal. Calorim. 2018, 5, 1425–1432. [Google Scholar] [CrossRef]
- Hofmann, T.; Tálos-nebehaj, E.; Albert, L.; Németh, L. Antioxidant Efficiency of Beech (Fagus sylvatica L.) Bark Polyphenols Assessed by Chemometric Methods. Ind. Crop. Prod. 2017, 108, 26–35. [Google Scholar] [CrossRef]
- Wei, X.; Cui, S.; Xie, Y. Synthesis and Antibacterial Properties of Oligomeric Dehydrogenation Polymer from Lignin Precursors. Molecules 2022, 27, 1466. [Google Scholar] [CrossRef] [PubMed]
- Dewijanti, I.D.; Mangunwardoyo, W.; Dwiranti, A.; Hanafi, M.; Artanti, N. Short Communication: Effects of the Various Source Areas of Indonesian Bay Leaves (Syzygium polyanthum) on Chemical Content and Antidiabetic Activity. Biodiversitas 2020, 21, 1190–1195. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Yu, J.S.; Lee, S.R.; Hwang, G.S.; Kang, K.S.; Park, J.G.; Kim, H.Y.; Kim, K.H.; Yamabe, N. Beneficial Effects of Bioactive Compounds in Mulberry Fruits against Cisplatin-Induced Nephrotoxicity. Int. J. Mol. Sci. 2018, 19, 1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckingham, J.; Munasinghe, V.R.N. Dictionary of Flavonoids with CD-ROM, 1st ed.; Taylor & Francis: Abingdon, UK, 2015. [Google Scholar]
- Khole, S.; Panat, N.A.; Suryawanshi, P.; Chatterjee, S.; Devasagayam, T.P.A.; Ghaskadbi, S. Comprehensive Assessment of Antioxidant Activities of Apigenin Isomers: Vitexin and Isovitexin. Free Radic. Antioxid. 2016, 6, 155–166. [Google Scholar] [CrossRef]
- Awolola, G.V.; Koorbanally, N.A.; Chenia, H.; Shode, F.O.; Baijnath, H. Antibacterial and anti-biofilm activity of flavonoids and triterpenes isolated from the extracts of Ficus sansibarica warb. subsp. sansibarica (Moraceae) extract. Afr. J. Tradit. Complement. Altern. Med. 2014, 11, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Adamczak, A.; Ozarowski, M.; Karpinski, T.M. Antibacterial Activity of Some Flavonoids and Organic Acids Widely Distributed in Plants. J. Clin. Med. 2020, 9, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Li, W.; Son, Y.; Sun, L.; Lu, J.; Kim, D.; Wang, X.; Yao, H.; Wang, L.; Pratheeshkumar, P.; et al. Quercitrin Protects Skin from UVB-Induced Oxidative Damage. Toxicol. Appl. Pharmacol. 2013, 269, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Ajibesin, K.K.; Essien, E.E.; Adesanya, S.A. Antibacterial Constituents of the Leaves of Dacryodes Edulis. Afr. J. Pharm. Pharmacol. 2011, 5, 1782–1786. [Google Scholar] [CrossRef] [Green Version]
- Shahid, F.; Alghamdi, Y.S.; Mashraqi, M.; Khurshid, M.; Ashfaq, U.A. Proteome Based Mapping and Molecular Docking Revealed DnaA as a Potential Drug Target against Shigella sonnei. Saudi J. Biol. Sci. 2022, 29, 1147–1159. [Google Scholar] [CrossRef]
- Albert, N.W.; Thrimawithana, A.H.; Mcghie, T.K.; Clayton, W.A.; Deroles, S.C.; Schwinn, K.E.; Bowman, J.L.; Jordan, B.R.; Davies, K.M. Genetic Analysis of the Liverwort Marchantia polymorpha Reveals that R2R3MYB Activation of Flavonoid Production in Response to Abiotic Stress Is an Ancient Character in Land Plants. New Phytol. 2017, 218, 554–566. [Google Scholar] [CrossRef] [Green Version]
- Carella, P.; Gogleva, A.; Hoey, D.J.; Bridgen, A.J.; Stolze, S.C.; Nakagami, H.; Schornack, S. Conserved Biochemical Defenses Underpin Host Responses to Oomycete Infection in an Early-Article Conserved Biochemical Defenses Underpin Host Responses to Oomycete Infection in an Early-Divergent Land Plant Lineage. Curr. Biol. 2019, 29, 2282–2294. [Google Scholar] [CrossRef]
- Tavakoli-Yaraki, M.; Karami-Tehrani, F. Apoptosis Induced by 13-S-Hydroxyoctadecadienoic Acid in the Breast Cancer Cell Lines, MCF-7 and MDA-MB-231. Iran. J. Basic Med. Sci. 2013, 16, 653–659. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Alkaloid | Triterpenoid | Steroid | Quinone | Flavonoid | Saponin | Tannin |
|---|---|---|---|---|---|---|---|
| Sonneratia caseolaris leaves | - | - | + | - | ++ | - | +++ |
| Avicennia marina leaves | - | - | + | - | ++ | ++ | ++ |
| Rhizophora mucronata leaves | - | - | ++ | - | + | +++ | +++ |
| Rhizophora apiculata leaves | - | - | ++ | - | - | + | ++ |
| Plant Species and Parts | Types of Solvent | Mass Yield Percentage (%) | LC50 (ppm) on A. salina |
|---|---|---|---|
| S. caseolaris leaves | N-hexane | 1.517 ± 0.254 | 633.59 |
| Ethyl Acetate | 1.431 ± 0.089 | 546.08 | |
| Ethanol | 9.464 ± 0.096 | 1076.05 | |
| Water | 9.380 ± 4.208 | 488.93 | |
| A. marina leaves | N-hexane | 1.829 ± 0.676 | 878.62 |
| Ethyl Acetate | 1.398 ± 0.072 | 1254.44 | |
| Ethanol | 7.422 ± 1.171 | 229.77 | |
| Water | 26.227 ± 5.130 | 847.63 | |
| R. mucronata leaves | N-hexane | 1.036 ± 0.133 | 160.43 |
| Ethyl Acetate | 2.358 ± 0.741 | 224.45 | |
| Ethanol | 2.485 ± 0.511 | 498.28 | |
| Water | 13.263 ± 3.745 | 772.70 | |
| R. apiculata leaves | N-hexane | 1.281 ± 0.381 | 734.36 |
| Ethyl Acetate | 3.220 ± 2.090 | 918.78 | |
| Ethanol | 6.546 ± 3.695 | 722.40 | |
| Water | 16.196 ± 5.198 | 627.48 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Audah, K.A.; Ettin, J.; Darmadi, J.; Azizah, N.N.; Anisa, A.S.; Hermawan, T.D.F.; Tjampakasari, C.R.; Heryanto, R.; Ismail, I.S.; Batubara, I. Indonesian Mangrove Sonneratia caseolaris Leaves Ethanol Extract Is a Potential Super Antioxidant and Anti Methicillin-Resistant Staphylococcus aureus Drug. Molecules 2022, 27, 8369. https://doi.org/10.3390/molecules27238369
Audah KA, Ettin J, Darmadi J, Azizah NN, Anisa AS, Hermawan TDF, Tjampakasari CR, Heryanto R, Ismail IS, Batubara I. Indonesian Mangrove Sonneratia caseolaris Leaves Ethanol Extract Is a Potential Super Antioxidant and Anti Methicillin-Resistant Staphylococcus aureus Drug. Molecules. 2022; 27(23):8369. https://doi.org/10.3390/molecules27238369
Chicago/Turabian StyleAudah, Kholis Abdurachim, Jufendi Ettin, Jason Darmadi, Norma Nur Azizah, Amalda Siti Anisa, Tedi Dwi Fauzi Hermawan, Conny Riana Tjampakasari, Rudi Heryanto, Intan Safinar Ismail, and Irmanida Batubara. 2022. "Indonesian Mangrove Sonneratia caseolaris Leaves Ethanol Extract Is a Potential Super Antioxidant and Anti Methicillin-Resistant Staphylococcus aureus Drug" Molecules 27, no. 23: 8369. https://doi.org/10.3390/molecules27238369