
Biological Activity of Pumpkin Byproducts: Antimicrobial and Antioxidant Properties

 , ,
, ,  , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Phenolic Compounds Profile
2.2. Antioxidant Acitivity
2.3. Antimicrobial and Antifungal Activity
2.4. Cytotoxic Potential
3. Materials and Methods
3.1. Sample Preparation
3.2. Characterization of the Phenolic Compounds Profile
3.3. Bioactive Performance
3.4. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rocchetti, G.; Pagnossa, J.P.; Blasi, F.; Cossignani, L.; Hilsdorf Piccoli, R.; Zengin, G.; Montesano, D.; Cocconcelli, P.S.; Lucini, L. Phenolic profiling and in vitro bioactivity of Moringa oleifera leaves as affected by different extraction solvents. Food Res. Int. 2020, 127, 108712. [Google Scholar] [CrossRef] [PubMed]
- Ceccanti, C.; Finimundy, T.C.; Melgar, B.; Pereira, C.; Ferreira, I.C.F.R.; Barros, L. Sequential steps of the incorporation of bioactive plant extracts from wild Italian Plantago coronopus L. and Cichorium intybus L. leaves in fresh egg pasta. Food Chem. 2022, 384, 132462. [Google Scholar] [CrossRef] [PubMed]
- Takwa, S.; Caleja, C.; Barreira, J.C.M.; Soković, M.; Achour, L.; Barros, L.; Ferreira, I.C.F.R. Arbutus unedo L. and Ocimum basilicum L. as sources of natural preservatives for food industry: A case study using loaf bread. LWT 2018, 88, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Jesus, M.S.; Genisheva, Z.; Romaní, A.; Pereira, R.N.; Teixeira, J.A.; Domingues, L. Bioactive compounds recovery optimization from vine pruning residues using conventional heating and microwave-assisted extraction methods. Ind. Crops Prod. 2019, 132, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Besrour, N.; Oludemi, T.; Mandim, F.; Pereira, C.; Dias, M.I.; Soković, M.; Stojković, D.; Ferreira, O.; Ferreira, I.C.F.R.; Barros, L. Valorization of Juglans regia Leaves as Cosmeceutical Ingredients: Bioactivity Evaluation and Final Formulation Development. Antioxidants 2022, 11, 677. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.C.; Chen, C.C.; Lin, K.H.; Chao, P.Y.; Lin, H.H.; Huang, M.Y. Bioactive compounds, antioxidants, and health benefits of sweet potato leaves. Molecules 2021, 26, 1820. [Google Scholar] [CrossRef]
- Roriz, C.L.; Heleno, S.A.; Alves, M.J.; Oliveira, M.B.P.P.; Pinela, J.; Dias, M.I.; Calhelha, R.C.; Morales, P.; Ferreira, I.C.F.R.; Barros, L. Red pitaya (Hylocereus costaricensis) peel as a source of valuable molecules: Extraction optimization to recover natural colouring agents. Food Chem. 2022, 372, 131344. [Google Scholar] [CrossRef]
- Sharma, P.; Kaur, G.; Kehinde, B.A.; Chhikara, N.; Panghal, A.; Kaur, H. Pharmacological and biomedical uses of extracts of pumpkin and its relatives and applications in the food industry: A review. Int. J. Veg. Sci. 2020, 26, 79–95. [Google Scholar] [CrossRef]
- Dhiman, A.K.; Sharma, K.D.; Attri, S. Functional constituents and processing of pumpkin: A review. J. Food Sci. Technol. 2009, 46, 411–417. [Google Scholar]
- Kaur, S.; Panghal, A.; Garg, M.K.; Mann, S.; Khatkar, S.K.; Sharma, P.; Chhikara, N. Functional and nutraceutical properties of pumpkin—A review. Nutr. Food Sci. 2020, 50, 384–401. [Google Scholar] [CrossRef]
- Ratnam, N.; Vandana; Najibullah, M.; Ibrahim, M. A Review on Cucurbita pepo. Int. J. Pharmacogn. Phytochem. Res. 2017, 9, 1190–1194. [Google Scholar] [CrossRef]
- Adnan, M.; Gul, S.; Batool, S.; Fatima, B.; Rehman, A.; Yaqoob, S.; Shabir, H.; Yousaf, T.; Mussarat, S.; Ali, N.; et al. A review on the ethnobotany, phytochemistry, pharmacology and nutritional composition of Cucurbita pepo L. J. Phytopharm. 2017, 6, 133–139. [Google Scholar] [CrossRef]
- Rico, X.; Gullón, B.; Alonso, J.L.; Yáñez, R. Recovery of high value-added compounds from pineapple, melon, watermelon and pumpkin processing by-products: An overview. Food Res. Int. 2020, 132, 109086. [Google Scholar] [CrossRef]
- Amin, M.Z.; Rity, T.I.; Uddin, M.R.; Rahman, M.M.; Uddin, M.J. A comparative assessment of anti-inflammatory, anti-oxidant and anti-bacterial activities of hybrid and indigenous varieties of pumpkin (Cucurbita maxima Linn.) seed oil. Biocatal. Agric. Biotechnol. 2020, 28, 101767. [Google Scholar] [CrossRef]
- Villamil, R.A.; Escobar, N.; Romero, L.N.; Huesa, R.; Plazas, A.V.; Gutiérrez, C.; Robelto, G.E. Perspectives of pumpkin pulp and pumpkin shell and seeds uses as ingredients in food formulation. Nutr. Food Sci. 2022. ahead-of-print. [Google Scholar] [CrossRef]
- Mala, K.S.; Kurian, A.E. Nutritional composition and antioxidant activity of pumpkin wastes. Int. J. Pharm. Chem. Biol. Sci. 2016, 6, 336–344. [Google Scholar]
- Adeel, R.; Sohail, A.; Masud, T. Characterization and antibacterial study of pumpkin seed oil (Cucurbita pepo). Life Sci. Leafl. 2014, 49, 53–64. [Google Scholar]
- Rezig, L.; Chouaibi, M.; Msaada, K.; Hamdi, S. Chemical composition and profile characterisation of pumpkin (Cucurbita maxima) seed oil. Ind. Crops Prod. 2012, 37, 82–87. [Google Scholar] [CrossRef]
- Yang, Z.; Shi, L.; Qi, Y.; Xie, C.; Zhao, W.; Barrow, C.J.; Dunshea, F.R.; Suleria, H.A.R. Effect of processing on polyphenols in butternut pumpkin (Cucurbita moschata). Food Biosci. 2022, 49, 101925. [Google Scholar] [CrossRef]
- Nyam, K.L.; Lau, M.; Tan, C.P. Fibre from pumpkin (Cucurbita pepo L.) Seeds and rinds: Physico-chemical properties, antioxidant capacity and application as bakery product ingredients. Malays. J. Nutr. 2013, 19, 99–110. [Google Scholar]
- Jaiswal, R.; Kuhnert, N. Identification and characterization of the phenolic glycosides of Lagenaria siceraria stand. (Bottle Gourd) fruit by liquid chromatography-tandem mass spectrometry. J. Agric. Food Chem. 2014, 62, 1261–1271. [Google Scholar] [CrossRef]
- Iswaldi, I.; Gómez-Caravaca, A.M.; Lozano-Sánchez, J.; Arráez-Román, D.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Profiling of phenolic and other polar compounds in zucchini (Cucurbita pepo L.) by reverse-phase high-performance liquid chromatography coupled to quadrupole time-of-flight mass spectrometry. Food Res. Int. 2013, 50, 77–84. [Google Scholar] [CrossRef]
- Busuioc, A.C.; Botezatu, A.V.D.; Furdui, B.; Vinatoru, C.; Maggi, F.; Caprioli, G.; Dinica, R.M. Comparative study of the chemical compositions and antioxidant activities of fresh juices from Romanian Cucurbitaceae varieties. Molecules 2020, 25, 5468. [Google Scholar] [CrossRef] [PubMed]
- Mokhtar, M.; Bouamar, S.; Di Lorenzo, A.; Temporini, C.; Daglia, M.; Riazi, A. The influence of ripeness on the phenolic content, antioxidant and antimicrobial activities of pumpkins (Cucurbita moschata Duchesne). Molecules 2021, 26, 3623. [Google Scholar] [CrossRef] [PubMed]
- Takebayashi, J.; Chen, J.; Tai, A. A. A Method for Evaluation of Antioxidant Activity Based on Inhibition of Free Radical-Induced Erythrocyte Hemolysis. In Advanced Protocols in Oxidative Stress II; Humana Press: Totowa, NJ, USA, 2010; Volume 594, pp. 287–296. [Google Scholar]
- Akomolafe, S.F.; Oboh, G.; Oyeleye, S.I.; Molehin, O.R.; Ogunsuyi, O.B. Phenolic composition and inhibitory ability of methanolic extract from pumpkin (Cucurbita pepo L.) seeds on Fe-induced thiobarbituric acid reactive species in albino rat’s testicular tissue in-vitro. J. Appl. Pharm. Sci. 2016, 6, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Saavedra, M.J.; Aires, A.; Dias, C.; Almeida, J.A.; De Vasconcelos, M.C.B.M.; Santos, P.; Rosa, E.A. Evaluation of the potential of squash pumpkin by-products (seeds and shell) as sources of antioxidant and bioactive compounds. J. Food Sci. Technol. 2015, 52, 1008–1015. [Google Scholar] [CrossRef] [Green Version]
- Longato, E.; Lucas-González, R.; Peiretti, P.G.; Meineri, G.; Pérez-Alvarez, J.A.; Viuda-Martos, M.; Fernández-López, J. The effect of natural ingredients (Amaranth and Pumpkin Seeds) on the quality properties of chicken burgers. Food Bioprocess Technol. 2017, 10, 2060–2068. [Google Scholar] [CrossRef]
- European Food Safety Authority and European Centre for Disease Prevention and Control (EFSA and ECDC) The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 2018, 16, e05500. [CrossRef] [Green Version]
- Wang, J.; Liu, M.; Wang, H.; Wu, Q.; Ding, Y.; Xu, T.; Ma, G.; Zhong, Y.; Zhang, J.; Chen, M.; et al. Occurrence, molecular characterization, and antimicrobial susceptibility of Yersinia enterocolitica isolated from retail food samples in China. LWT 2021, 150, 111876. [Google Scholar] [CrossRef]
- Soares, K.; Moura, A.T.; García-Díez, J.; Oliveira, I.; Esteves, A.; Saraiva, C. Evaluation of Hygienic Quality of Food Served in Universities Canteens of Northem Portugal. Indian J. Microbiol. 2020, 60, 107–114. [Google Scholar] [CrossRef]
- Paterson, R.R.M.; Lima, N. Filamentous fungal human pathogens from food emphasising Aspergillus, Fusarium and Mucor. Microorganisms 2017, 5, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Racchi, I.; Scaramuzza, N.; Hidalgo, A.; Cigarini, M.; Berni, E. Sterilization of food packaging by UV-C irradiation: Is Aspergillus brasiliensis ATCC 16404 the best target microorganism for industrial bio-validations? Int. J. Food Microbiol. 2021, 357, 109383. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Cho, A.R.; Han, J. Antioxidant and antimicrobial activities of leafy green vegetable extracts and their applications to meat product preservation. Food Control 2013, 29, 112–120. [Google Scholar] [CrossRef]
- Dotto, J.M.; Chacha, J.S. The potential of pumpkin seeds as a functional food ingredient: A review. Sci. Afr. 2020, 10, e00575. [Google Scholar] [CrossRef]
- Abou Seif, H.S. Ameliorative effect of pumpkin oil (Cucurbita pepo L.) against alcohol-induced hepatotoxicity and oxidative stress in albino rats. Beni-Suef Univ. J. Basic Appl. Sci. 2014, 3, 178–185. [Google Scholar] [CrossRef] [Green Version]
- Barros, L.; Pereira, E.; Calhelha, R.C.; Dueñas, M.; Carvalho, A.M.; Santos-Buelga, C.; Ferreira, I.C.F.R. Bioactivity and chemical characterization in hydrophilic and lipophilic compounds of Chenopodium ambrosioides L. J. Funct. Foods 2013, 5, 1732–1740. [Google Scholar] [CrossRef]
- Takebayashi, J.; Iwahashi, N.; Ishimi, Y.; Tai, A. Development of a simple 96-well plate method for evaluation of antioxidant activity based on the oxidative haemolysis inhibition assay (OxHLIA). Food Chem. 2012, 134, 606–610. [Google Scholar] [CrossRef]
- Barros, L.; Carvalho, A.M.; Morais, J.S.; Ferreira, I.C.F.R. Strawberry-tree, blackthorn and rose fruits: Detailed characterisation in nutrients and phytochemicals with antioxidant properties. Food Chem. 2010, 120, 247–254. [Google Scholar] [CrossRef]
- Heleno, S.A.; Ferreira, I.C.F.R.; Esteves, A.P.; Ćirić, A.; Glamočlija, J.; Martins, A.; Soković, M.; João, M.; Queiroz, R.P. Antimicrobial and demelanizing activity of Ganoderma lucidum extract, p-hydroxybenzoic and cinnamic acids and their synthetic acetylated glucuronide methyl esters. Food Chem. Toxicol. 2013, 58, 95–100. [Google Scholar] [CrossRef]
- Kuete, V.; Ango, P.Y.; Fotso, G.W.; Dwf Kapche, G.; Dzoyem, J.P.; Wouking, A.G.; Ngadjui, B.T.; Abegaz, B.M. Antimicrobial activities of the methanol extract and compounds from Artocarpus communis (Moraceae). BMC Complement. Altern. Med. 2011, 25, 11–42. [Google Scholar] [CrossRef] [Green Version]
- Guimarães, R.; Barros, L.; Dueñas, M.; Calhelha, R.C.; Carvalho, A.M.; Santos-Buelga, C.; Queiroz, M.J.R.P.; Ferreira, I.C.F.R. Nutrients, phytochemicals and bioactivity of wild Roman chamomile: A comparison between the herb and its preparations. Food Chem. 2013, 136, 718–725. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
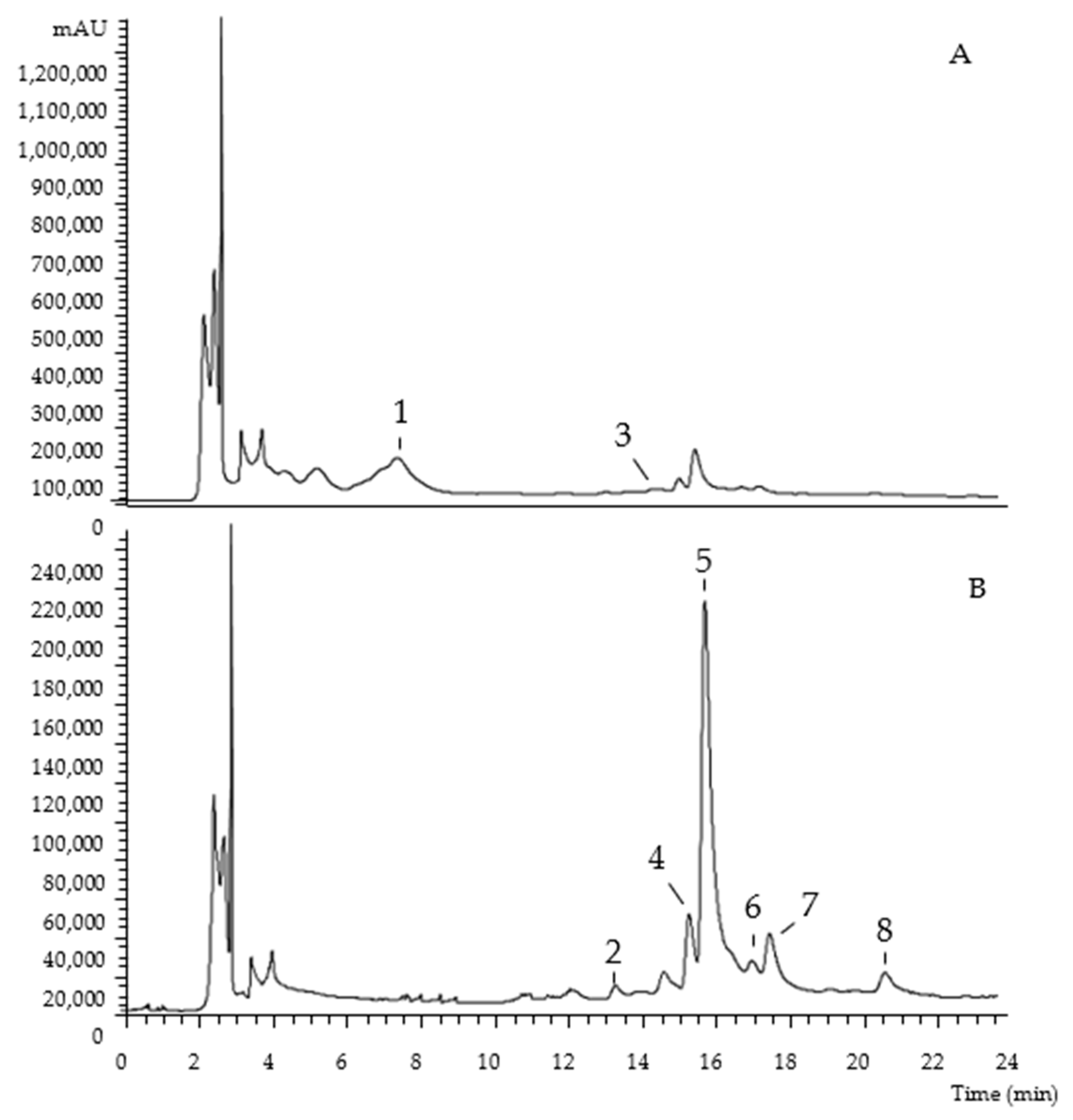
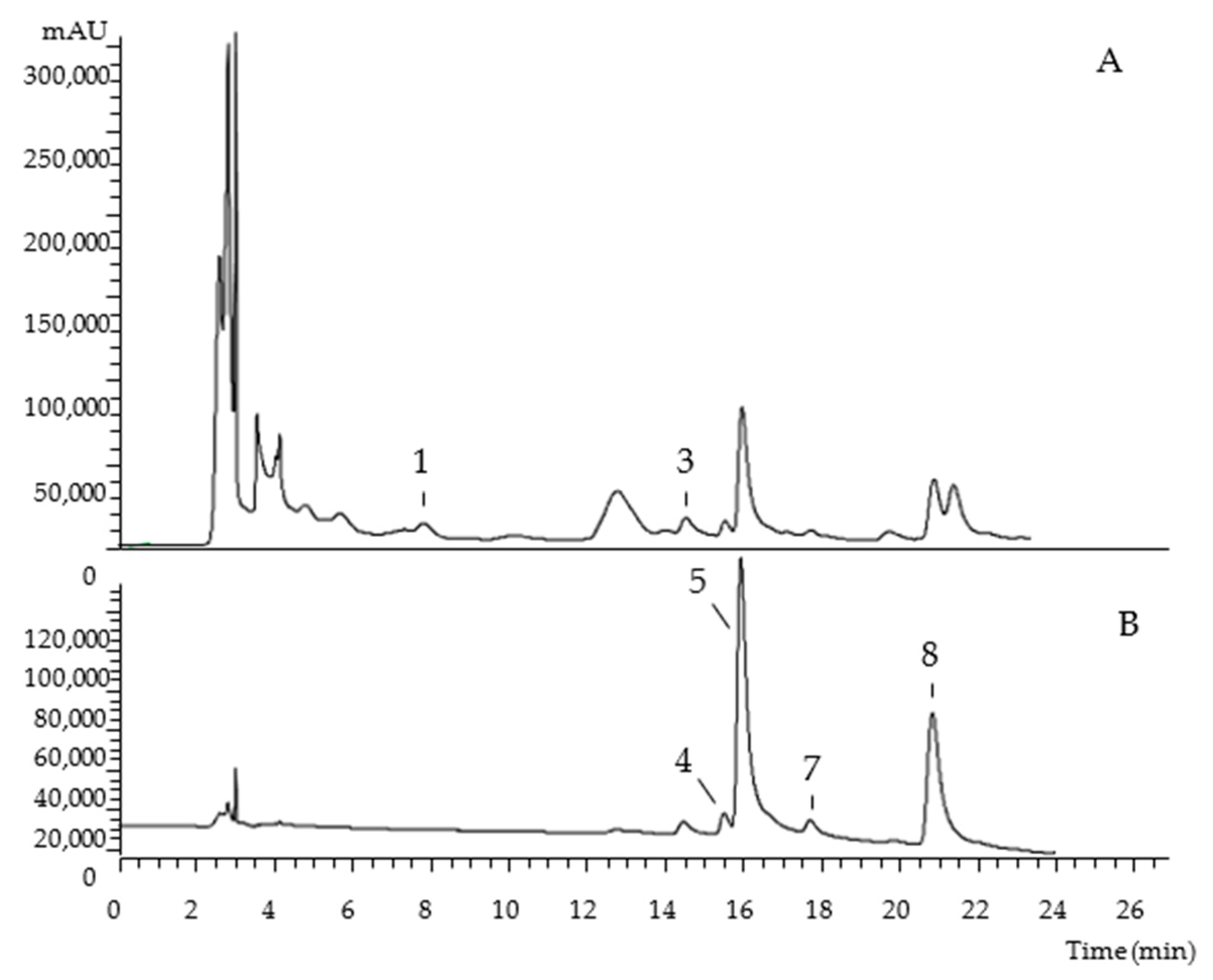
| Peak | Rt (min) | λmax (nm) | [M-H]− (m/z) | MS2 (m/z) | Tentative Identification |
|---|---|---|---|---|---|
| 1 | 7.71 | 280 | 289 | 245 (100), 205 (45) | (-)-Epicatechin |
| 2 | 13.42 | 345 | 775 | 301 (100) | Quercetin-O-dideoxyhexosyl-hexoside |
| 3 | 14.73 | 263 | 405 | 281 (100), 137 (12), 93 (5) | 7 4-O-(6′-O-Glucosyl-4″-hydroxybenzoyl)-4-hydroxybenzyl alcohol |
| 4 | 15.42 | 344 | 739 | 285 (100) | Kaempferol-O-dideoxyhexosyl-hexoside |
| 5 | 15.85 | 354 | 769 | 315 (100) | Isorhamnetin-O-dideoxyhexosyl-hexoside |
| 6 | 17.12 | 348 | 593 | 285 (100) | Kaempferol-O-deoxyhexosyl-hexoside |
| 7 | 17.6 | 365 | 623 | 315 (100) | Isorhamnetin-O-deoxyhexosyl-hexoside |
| 8 | 20.73 | 365 | 623 | 315 (100) | Isorhamnetin-O-deoxyhexosyl-hexoside |
| Peak | Common Pumpkin | Butternut Squash | Kabocha Squash | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Peel | Fibrous Strands | Seeds | Peel | Fibrous Strands | Seeds | Peel | Fibrous Strands | Seeds | |
| 1 | 4.58 ± 0.08 a | 3.04 ± 0.05 b | 1.74 ± 0.03 f | 2.56 ± 0.03 d,e | 2.47 ± 0.07 e | 2.63 ± 0.02 c,d | 1.50 ± 0.07 g | 2.7 ± 0.1 c | 1.29 ± 0.05 h |
| 2 | 0.50 ± 0.02 b | 0.484 ± 0.006 b | 0.49 ± 0.02 b | n.d. | n.d. | n.d. | n.d. | 0.533 ± 0.003 a | 0.474 ± 0.001 b |
| 3 | 0.214 ± 0.009 a | n.d. | n.d. | n.d. | n.d. | n.d. | 0.116 ± 0.004 b | n.d. | n.d. |
| 4 | 0.65 ± 0.03 a | 0.487 ± 0.007 e | 0.458 ± 0.008 f | 0.6096 ± 0.0002 b | n.d. | 0.457 ± 0.008 f | 0.58 ± 0.02 c | 0.5218 ± 0.0004 d | 0.461 ± 0.003 e,f |
| 5 | 1.60 ± 0.08 a | n.d. | n.d. | 0.543 ± 0.004 c | n.d. | 0.454 ± 0.007 d | 0.62 ± 0.03 b | 0.494 ± 0.002 c,d | n.d. |
| 6 | 0.59 ± 0.03 a | n.d. | n.d. | 0.519 ± 0.005 b | n.d. | n.d. | 0.58 ± 0.03 a | 0.493 ± 0.003 b | n.d. |
| 7 | 0.69 ± 0.03 a | n.d. | n.d. | 0.496 ± 0.005 c | n.d. | n.d. | 0.56 ± 0.02 b | n.d. | n.d. |
| 8 | 0.55 ± 0.03 a | n.d. | n.d. | n.d. | n.d. | n.d. | 0.54 ± 0.03 a | n.d. | n.d. |
| Total flavan-3-ols | 4.58 ± 0.08 a | 3.04 ± 0.05 b | 1.74 ± 0.03 f | 2.56 ± 0.03 d,e | 2.47 ± 0.07 e | 2.63 ± 0.02 c,d | 1.50 ± 0.07 g | 2.7 ± 0.1 c | 1.29 ± 0.05 h |
| Total phenolic acids | 0.214 ± 0.009 a | n.d. | n.d. | n.d. | n.d. | n.d. | 0.116 ± 0.004 b | n.d. | n.d. |
| Total flavonoids | 4.6 ± 0.2 a | 0.97 ± 0.01 d | 0.95 ± 0.03 d | 2.17 ± 0.01 c | n.d. | 0.91 ± 0.02 d | 2.9 ± 0.1 b | 2.042 ± 0.003 c | 0.934 ± 0.002 d |
| Total phenolic compounds | 9.4 ± 0.3 a | 4.01 ± 0.06 d | 2.693 ± 0.004 f | 4.73 ± 0.01 b,c | 2.47 ± 0.07 f | 3.538 ± 0.007 e | 4.50 ± 0.06 c | 4.8 ± 0.1 b | 2.23 ± 0.04 g |
| Peak | Gold Nugget Pumpkin | Butternut Squash | Musquée de Provence | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Fibrous Strands | Seeds | Peel | Fibrous Strands | Seeds | Peel | Fibrous Strands | Seeds | Peel | |
| 1 | 1.66 ± 0.07 a | n.d. | 0.51 ± 0.02 e | 0.97 ± 0.03 c | n.d. | 0.346 ± 0.009 f | 1.13 ± 0.01 b | n.d. | 0.577 ± 0.008 d |
| 3 | 2.27 ± 0.02 a | n.d. | 0.134 ± 0.006 d | 0.176 ± 0.006 c | n.d. | tr. | 0.207 ± 0.007 b | 0.0762 ± 0.0002 e | n.d. |
| 4 | n.d. | n.d. | 0.94 ± 0.05 a | 0.462 ± 0.002 c | n.d. | 0.4526 ± 0.0003 c | 0.4790 ± 0.0007 b | n.d. | 0.476 ± 0.003 b |
| 5 | n.d. | n.d. | 1.65 ± 0.07 a | n.d. | n.d. | 0.44904 ± 0.00006 b | n.d. | 0.460 ± 0.005 b | 0.474 ± 0.002 b |
| 7 | n.d. | n.d. | n.d. | n.d. | n.d. | 0.4418 ± 0.0001 | n.d. | n.d. | n.d. |
| 8 | n.d. | n.d. | 0.85 ± 0.02 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Total flavan-3-ols | 1.66 ± 0.07 a | n.d. | 0.51 ± 0.02 e | 0.97 ± 0.03 c | n.d. | 0.346 ± 0.009 f | 1.13 ± 0.01 b | n.d. | 0.577 ± 0.008 d |
| Total phenolic acids | 2.27 ± 0.02 a | n.d. | 0.134 ± 0.006 d | 0.176 ± 0.006 c | n.d. | tr. | 0.207 ± 0.007 b | 0.0762 ± 0.0002 e | n.d. |
| Total flavonoids | n.d. | n.d. | 3.4 ± 0.1 a | 0.462 ± 0.002 d | n.d. | 1.3434 ± 0.0004 b | 0.4790 ± 0.0007 d | 0.460 ± 0.005 d | 0.951 ± 0.005 c |
| Total phenolic compounds | 3.93 ± 0.05 b | n.d. | 4.1 ± 0.1 a | 1.61 ± 0.03 d,e | n.d. | 1.689 ± 0.008 d | 1.818 ± 0.007 c | 0.536 ± 0.005 f | 1.53 ± 0.01 e |
| Pumpkin Genotype from Portugal | Part | OxHLIA 60 min IC50 1, μg/mL | TBARS IC50 1, μg/mL |
|---|---|---|---|
| Butternut Squash | Peel | 88 ± 3 c | 7461 ± 315 b |
| Seeds | 59 ± 6 d | 185 ± 7 h | |
| Fibrous strands | 44 ± 4 d | 6887 ± 53 c | |
| Common Pumpkin | Peel | 90 ± 3 c | 3921 ± 33 e |
| Seeds | 43 ± 3 d | 756 ± 27 g | |
| Fibrous strands | 365 ± 13 a | 6375 ± 68 d | |
| Kabocha Squash | Peel | 209 ± 10 b | 7765 ± 31 a |
| Seeds | 46 ± 2 d | 164 ± 8 h | |
| Fibrous strands | 96 ± 2 c | 1568 ± 53 f | |
| Trolox | 21.8 ± 0.2 e | 139 ± 5 h |
| Pumpkin Genotype from Algeria | Part | OxHLIA 60 min IC50 1, μg/mL | TBARS IC50 1, μg/mL |
|---|---|---|---|
| Butternut Squash | Peel | 588 ± 18 a | 4569 ± 277 a |
| Seeds | 115 ± 6 f | 573 ± 31 e | |
| Fibrous strands | 257 ± 13 d | 3508 ± 91 b | |
| Gold Nugget Pumpkin | Peel | 362 ± 8 b,c | 3123 ± 136 c |
| Seeds | n.d. 2 | 91 ± 4 f | |
| Fibrous strands | 566 ± 13 a | 3659 ± 199 b | |
| Musquée de Provence | Peel | 335 ± 4 c | 2123 ± 101 d |
| Seeds | 400 ± 34 b | 549 ± 27 e | |
| Fibrous strands | 188 ± 2 e | 4385 ± 242 a | |
| Trolox | 21.8 ± 0.2 g | 139 ± 5 f |
| Butternut Squash | Common Pumpkin | Kabocha Squash | Streptomicin 1 mg/mL | Methicilin 1 mg/mL | Ampicillin 10 mg/mL | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Seeds | Peel | Fibrous Strands | Seeds | Peel | Fibrous Strands | Seeds | Peel | Fibrous Strands | ||||||||||||||||
| MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | |
| Gram-negative bacteria | ||||||||||||||||||||||||
| Enterobacter cloacae | 10 | >10 | 10 | >10 | 2.5 | >10 | 10 | >10 | 10 | >10 | >10 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | 0.007 | 0.007 | n.t. | n.t | 0.15 | 0.15 |
| Escherichia coli | >10 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | >10 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | 0.01 | 0.01 | n.t. | n.t. | 0.15 | 0.15 |
| Pseudomonas aeruginosa | >10 | >10 | >10 | >10 | 10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | 0.06 | 0.06 | n.t. | n.t. | 0.63 | 0.63 |
| Salmonella enterica | >10 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | >10 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | 0.007 | 0.007 | n.t. | n.t. | 0.15 | 0.15 |
| Yersinia enterocolitica | 10 | >10 | 5 | >10 | 5 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | 5 | >10 | 0.007 | 0.007 | n.t. | n.t. | 0.15 | 0.15 |
| Gram-positive bacteria | ||||||||||||||||||||||||
| Bacillus cereus | >10 | >10 | >10 | >10 | 5 | >10 | 10 | >10 | 2.5 | >10 | 10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | 0.007 | 0.007 | n.t. | n.t. | n.t. | n.t. |
| Listeria monocytogenes | >10 | >10 | >10 | >10 | 5 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | >10 | >10 | 10 | >10 | 10 | >10 | 0.007 | 0.007 | n.t. | n.t. | 0.15 | 0.15 |
| Staphylococcus aureus | 10 | >10 | >10 | >10 | 10 | >10 | 10 | >10 | 5 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | 0.007 | 0.007 | 0.007 | 0.007 | 0.15 | 0.15 |
| Butternut Squash | Common Pumpkin | Kabocha Squash | Ketoconazole | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Seeds | Peel | Fibrous Strands | Seeds | Peel | Fibrous Strands | Seeds | Peel | Fibrous Strands | ||||||||||||
| MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | |
| Aspergillus brasiliensis | 10 | >10 | 10 | >10 | 5 | >10 | 5 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | 0.06 | 0.125 |
| Aspergillus fumigatus | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | 0.5 | 1 |
| Gold Nugget Pumpkin | Butternut Squash | Musquée de Provence | Streptomicin 1 mg/mL | Methicilin 1 mg/mL | Ampicillin 10 mg/mL | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Seeds | Peel | Fibrous Strands | Seeds | Peel | Fibrous Strands | Seeds | Peel | Fibrous Strands | ||||||||||||||||
| MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | |
| Gram-negative bacteria | ||||||||||||||||||||||||
| Enterobacter Cloacae | 5 | >10 | 5 | >10 | >10 | >10 | 5 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | >10 | >10 | >10 | >10 | 0.007 | 0.007 | n.t. | n.t | 0.15 | 0.15 |
| Escherichia coli | 10 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | 10 | >10 | >10 | >10 | 10 | >10 | >10 | >10 | 0.01 | 0.01 | n.t. | n.t. | 0.15 | 0.15 |
| Pseudomonas aeruginosa | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | 0.06 | 0.06 | n.t. | n.t. | 0.63 | 0.63 |
| Salmonella enterica | 10 | >10 | 10 | >10 | >10 | >10 | 10 | >10 | 10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | 0.007 | 0.007 | n.t. | n.t. | 0.15 | 0.15 |
| Yersinia enterocolitica | 5 | >10 | 5 | >10 | 10 | >10 | 5 | >10 | 10 | >10 | >10 | >10 | 10 | >10 | >10 | >10 | >10 | >10 | 0.007 | 0.007 | n.t. | n.t. | 0.15 | 0.15 |
| Gram-positive bacteria | ||||||||||||||||||||||||
| Bacillus cereus | 5 | >10 | >10 | >10 | >10 | >10 | 2.5 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | >10 | 0.007 | 0.007 | n.t. | n.t. | n.t. | n.t. |
| Listeria monocytogenes | 2.5 | >10 | 5 | >10 | 10 | >10 | 10 | >10 | >10 | >10 | 5 | >10 | >10 | >10 | >10 | >10 | 5 | >10 | 0.007 | 0.007 | n.t. | n.t. | 0.15 | 0.15 |
| Staphylococcus aureus | 2.5 | >10 | 5 | >10 | 10 | >10 | 5 | >10 | 5 | >10 | 10 | >10 | 10 | >10 | 5 | >10 | 10 | >10 | 0.007 | 0.007 | 0.007 | 0.007 | 0.15 | 0.15 |
| Gold Nugget Pumpkin | Butternut Squash | Musquée de Provence | Ketoconazole | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Seeds | Peel | Fibrous Strands | Seeds | Peel | Fibrous Strands | Seeds | Peel | Fibrous Strands | ||||||||||||
| MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | |
| Aspergillus brasiliensis | 5 | >10 | 5 | >10 | 10 | >10 | 5 | >10 | 10 | >10 | 10 | >10 | 5 | >10 | 10 | >10 | 10 | >10 | 0.06 | 0.125 |
| Aspergillus fumigatus | >10 | >10 | >10 | >10 | 10 | >10 | >10 | >10 | >10 | >10 | 10 | >10 | >10 | >10 | >10 | >10 | 10 | >10 | 0.5 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leichtweis, M.G.; Molina, A.K.; Pires, T.C.S.; Dias, M.I.; Calhelha, R.; Bachari, K.; Ziani, B.E.C.; Oliveira, M.B.P.P.; Pereira, C.; Barros, L. Biological Activity of Pumpkin Byproducts: Antimicrobial and Antioxidant Properties. Molecules 2022, 27, 8366. https://doi.org/10.3390/molecules27238366
Leichtweis MG, Molina AK, Pires TCS, Dias MI, Calhelha R, Bachari K, Ziani BEC, Oliveira MBPP, Pereira C, Barros L. Biological Activity of Pumpkin Byproducts: Antimicrobial and Antioxidant Properties. Molecules. 2022; 27(23):8366. https://doi.org/10.3390/molecules27238366
Chicago/Turabian StyleLeichtweis, Maria G., Adriana K. Molina, Tânia C. S. Pires, Maria Inês Dias, Ricardo Calhelha, Khaldoun Bachari, Borhane E. C. Ziani, M. Beatriz P. P. Oliveira, Carla Pereira, and Lillian Barros. 2022. "Biological Activity of Pumpkin Byproducts: Antimicrobial and Antioxidant Properties" Molecules 27, no. 23: 8366. https://doi.org/10.3390/molecules27238366