
Tabersonine Inhibits the Lipopolysaccharide-Induced Neuroinflammatory Response in BV2 Microglia Cells via the NF-κB Signaling Pathway
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Result
2.1. Effect of Tab on the Viability of BV2 Cells
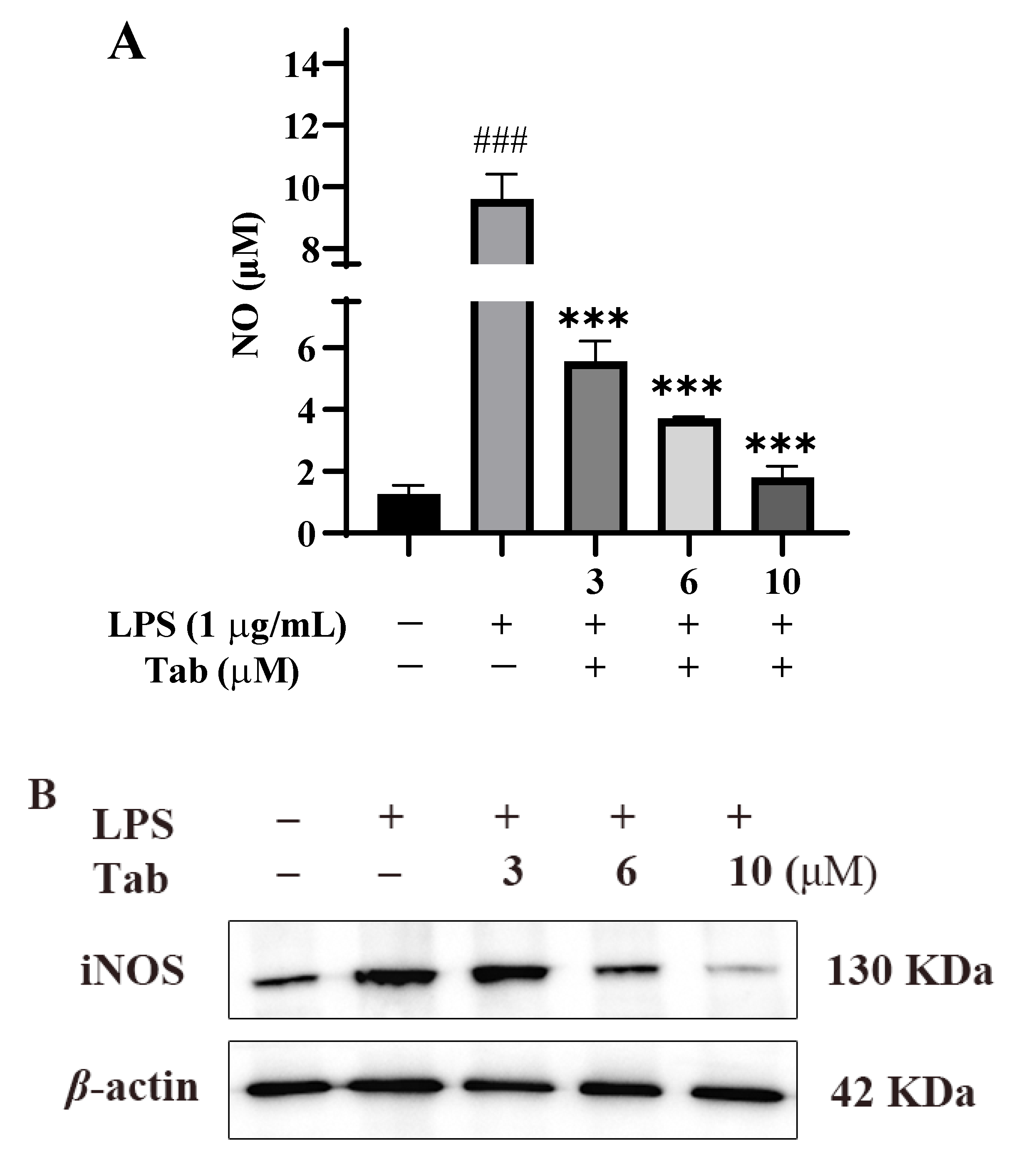
2.2. Effect of Tab on LPS-Induced Neuroinflammation
2.3. Effect of Tab on LPS-Induced Inflammatory Factors in BV2 Cells
2.4. Effect of Tab on LPS-Induced ROS in BV2 Cells
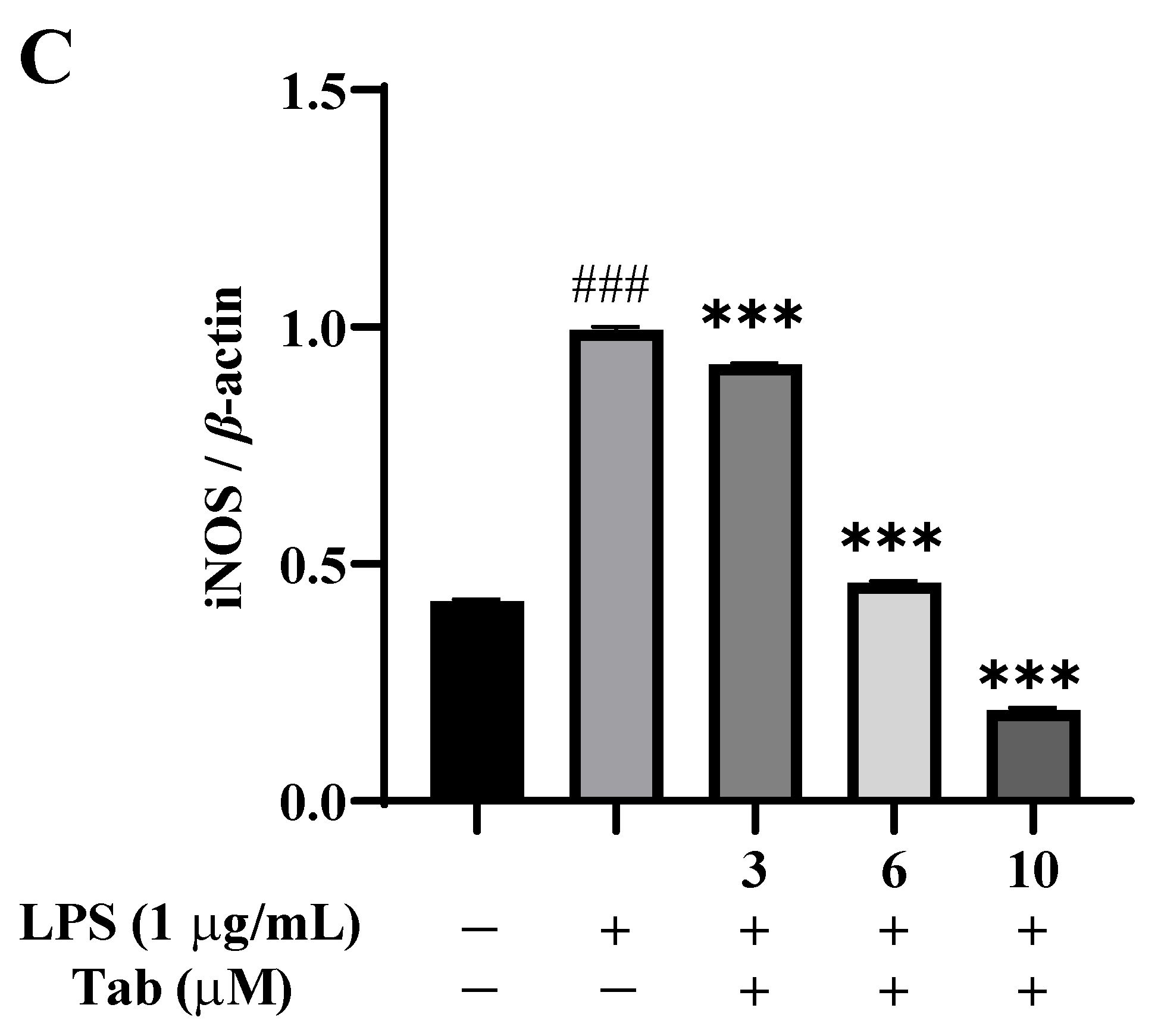
2.5. Tab Inhibits LPS-Induced Microglia Activation through NF-κB Pathway
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Viability
4.3. Nitric Oxide Assay
4.4. Detection of Inflammatory Factor Content
4.5. Immunocytochemical Staining
4.6. Reactive Oxygen Species Assay
4.7. Western Blot
4.8. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Shabab, T.; Khanabdali, R.; Moghadamtousi, S.Z.; Kadir, H.A.; Mohan, G. Neuroinflammation pathways: A general review. Int. J. Neurosci. 2017, 127, 624–633. [Google Scholar] [CrossRef]
- Carson, M.J.; Doose, J.M.; Melchior, B.; Schmid, C.D.; Ploix, C.C. CNS immune privilege: Hiding in plain sight. Immunol. Rev. 2009, 213, 48–65. [Google Scholar] [CrossRef] [PubMed]
- Carson, M.; Thrash, J.C.; Walter, B. The cellular response in neuroinflammation: The role of leukocytes, microglia and astrocytes in neuronal death and survival. Clin. Neurosci. Res. 2006, 6, 237–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, V.H.; Teeling, J. Microglia and macrophages of the central nervous system: The contribution of microglia priming and systemic inflammation to chronic neurodegeneration. Semin. Immunopathol. 2013, 35, 601–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zielasek, J.; Hartung, H.P. Molecular mechanisms of microglial activation. Adv. Neuroimmunol. 1996, 6, 191–192. [Google Scholar] [CrossRef]
- Block, M.L.; Zecca, L.; Hong, J.S. Microglia-mediated neurotoxicity: Uncovering the molecular mechanisms. Nat. Rev. Neurosci. 2007, 8, 57–69. [Google Scholar] [CrossRef]
- Batista, C.R.A.; Gomes, G.F.; Candelario-Jalil, E.; Fiebich, B.L.; de Oliveira, A.C.P. Lipopolysaccharide-Induced Neuroinflammation as a Bridge to Understand Neurodegeneration. Int. J. Mol. Sci. 2019, 20, 2293. [Google Scholar] [CrossRef] [Green Version]
- Gupta, N.; Shyamasundar, S.; Patnala, R.; Karthikeyan, A.; Arumugam, T.V.; Ling, E.A.; Dheen, S.T. Recent progress in therapeutic strategies for microglia-mediated neuroinflammation in neuropathologies. Expert Opin. Ther. Tar. 2018, 22, 765–781. [Google Scholar] [CrossRef]
- Liao, S.; Wu, J.N.; Liu, R.M.; Wang, S.X.; Luo, J.; Yang, Y.; Qin, Y.N.; Li, T.; Zheng, X.P.; Song, J. A novel compound DBZ ameliorates neuroinflammation in LPS-stimulated microglia and ischemic stroke rats: Role of Akt(Ser473)/GSK3β(Ser9)-mediated Nrf2 activation. Redox. Biol. 2020, 36, 101644. [Google Scholar] [CrossRef]
- Nejat, N.; Valdiani, A.; Cahill, D.; Tan, Y.H.; Maziah, M.; Abiri, R. Ornamental exterior versus therapeutic interior of Madagascar periwinkle (Catharanthus roseus): The two faces of a versatile herb. Sci. World J. 2015, 982412. [Google Scholar] [CrossRef]
- Li, X.; Deng, Y.; Kang, L.C.; Chen, L.H.; Zheng, Z.Z.; Huang, W.; Xu, C.J.; Kai, G.Y.; Lin, D.B.; Tong, Q.X.; et al. Cytotoxic active ingredients from the seeds of Voacanga africana. S. Afr. J. Bot. 2021, 137, 311–319. [Google Scholar] [CrossRef]
- Zhang, D.P.; Li, X.Z.; Hu, Y.D.; Jiang, H.C.; Wu, Y.X.; Ding, Y.H.; Yu, K.K.; He, H.Q.; Xu, J.S.; Sun, L.; et al. Tabersonine attenuates lipopolysaccharide-induced acute lung injury via suppressing TRAF6 ubiquitination. Biochem. Pharmacol. 2018, 154, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.Y.; Luo, W.; Chen, Y.H.; Shen, S.Y.; Wang, Z.; Chen, R.J.; Wang, J.; Chattipakorn, N.; Huang, W.J.; Liang, G. Tabersonine attenuates Angiotensin II-induced cardiac remodeling and dysfunction through targeting TAK1 and inhibiting TAK1-mediated cardiac inflammation. Phytomedicine 2022, 103, 154238. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.S.; Kim, Y.J.; Kim, B.Y.; Park, G.; Jeong, S.J. The Anti-neuroinflammatory Activity of Tectorigenin Pretreatment via Downregulated NF-κB and ERK/JNK Pathways in BV-2 Microglial and Microglia Inactivation in Mice With Lipopolysaccharide. Front. Pharmacol. 2018, 9, 462. [Google Scholar] [CrossRef] [PubMed]
- Kempe, S.; Kestler, H.; Lasar, A.; Wirth, T. NF-κB controls the global pro-inflammatory response in endothelial cells: Evidence for the regulation of a pro-atherogenic program. Nucleic Acids Res. 2005, 33, 5308–5319. [Google Scholar] [CrossRef]
- Moreno, R.; Sobotzik, J.M.; Schultz, C.; Schmitz, M.L. Specification of the NF-κB transcriptional response by p65 phosphorylation and TNF-induced nuclear translocation of IKKε. Nucleic Acids Res. 2010, 38, 6029–6044. [Google Scholar] [CrossRef] [Green Version]
- Skaug, B.; Jiang, X.; Chen, Z.J. The role of ubiquitin in NF-κB regulatory pathways. Annu. Rev. Biochem. 2009, 78, 769–796. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [Green Version]
- Wolf, S.A.; Boddeke, H.W.G.M.; Kettenmann, H. Microglia in Physiology and Disease. Annu. Rev. Physiol. 2017, 79, 619–643. [Google Scholar] [CrossRef]
- Jha, M.K.; Jo, M.; Kim, J.H.; Suk, K. Microglia-Astrocyte Crosstalk: An Intimate Molecular Conversation. Neuroscientist 2019, 25, 227–240. [Google Scholar] [CrossRef]
- Maksoud, M.J.E.; Tellios, V.; Xiang, Y.Y.; Lu, W.Y. Nitric oxide signaling inhibits microglia proliferation by activation of protein kinase-G. Nitric Oxide 2020, 94, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.Y.; Bi, W.; Xiao, S.; Lan, X.; Cheng, X.F.; Zhang, J.W.; Lu, D.X.; Wei, W.; Wang, Y.P.; Li, H.M.; et al. Neuroinflammation induced by lipopolysaccharide causes cognitive impairment in mice. Sci. Rep. 2019, 9, 5790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.S.; Liu, M.; Liu, D.N.; Yang, Y.L.; Du, G.H.; Wang, Y.H. TLR4-IN-C34 Inhibits Lipopolysaccharide-Stimulated Inflammatory Responses via Downregulating TLR4/MyD88/NF-κB/NLRP3 Signaling Pathway and Reducing ROS Generation in BV2 Cells. Inflammation 2022, 45, 838–850. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhang, S.S.; Liu, D.N.; Yang, Y.L.; Wang, Y.H.; Du, G.H. Chrysomycin A attenuates neuroinflammation by down-regulating NLRP3/Cleaved caspase-1 signaling pathway in LPS-stimulated mice and BV2 cells. Int. J. Mol. Sci. 2021, 22, 6799. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.L.; Bui, B.P.; Lee, H.; Cho, J. A novel 1,8-naphthyridine-2-carboxamide derivative attenuates inflammatory responses and cell migration in LPS-treated BV2 cells via the suppression of ROS generation and TLR4/Myd88/NF-κB signaling pathway. Int. J. Mol. Sci. 2021, 22, 2527. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Han, M.H.; Park, C.; Jin, C.Y.; Kim, G.Y.; Choi, I.W.; Kim, N.D.; Nam, T.J.; Kwon, T.K.; Choi, Y.H. Anti-inflammatory effects of fucoidan through inhibition of NF-κB, MAPK and Akt activation in lipopolysaccharide-induced BV2 microglia cells. Food Chem. Toxicol. 2011, 49, 1745–1752. [Google Scholar] [CrossRef]
- Guo, L.T.; Wang, S.Q.; Su, J.; Xu, L.X.; Ji, Z.Y.; Zhang, R.Y.; Zhao, Q.W.; Ma, Z.Q.; Deng, X.Y.; Ma, S.P. Baicalin ameliorates neuroinflammation-induced depressive-like behavior through inhibition of toll-like receptor 4 expression via the PI3K/AKT/FoxO1 pathway. J. Neuroinflammation 2019, 16, 95. [Google Scholar] [CrossRef] [Green Version]
- Marschallinger, J.; Iram, T.; Zardeneta, M.; Lee, S.E.; Lehallier, B.; Haney, M.S.; Pluvinage, J.V.; Mathur, V.; Hahn, O.; Morgens, D.W.; et al. Lipid-droplet-accumulating microglia represent a dysfunctional and proinflammatory state in the aging brain. Nature Neurosci. 2020, 23, 194–208. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, J.; Wang, C.; Sang, C.; Nsanzamahoro, S.; Chai, T.; Wang, J.; Yang, A.; Yang, J. Tabersonine Inhibits the Lipopolysaccharide-Induced Neuroinflammatory Response in BV2 Microglia Cells via the NF-κB Signaling Pathway. Molecules 2022, 27, 7521. https://doi.org/10.3390/molecules27217521
Shi J, Wang C, Sang C, Nsanzamahoro S, Chai T, Wang J, Yang A, Yang J. Tabersonine Inhibits the Lipopolysaccharide-Induced Neuroinflammatory Response in BV2 Microglia Cells via the NF-κB Signaling Pathway. Molecules. 2022; 27(21):7521. https://doi.org/10.3390/molecules27217521
Chicago/Turabian StyleShi, Jiaotai, Chengbo Wang, Chunyan Sang, Stanislas Nsanzamahoro, Tian Chai, Jun Wang, Aimei Yang, and Junli Yang. 2022. "Tabersonine Inhibits the Lipopolysaccharide-Induced Neuroinflammatory Response in BV2 Microglia Cells via the NF-κB Signaling Pathway" Molecules 27, no. 21: 7521. https://doi.org/10.3390/molecules27217521