

Chemotaxonomic Markers for the Leaf Buds of Common Finnish Trees and Shrubs: A Rapid UHPLC MS Fingerprinting Tool for Species Identification


Abstract
:1. Introduction
2. Results and Discussion
2.1. Two Approaches in the Development of the LC–MS Fingerprinting Method and Selection of Marker Candidates
2.1.1. Method I: Species-Specific Markers from the High-Resolution MS Data Obtained with MZmine 2
2.1.2. Method II: Main Ions from the QqQ Full Scan Spectra
2.1.3. Repeatability of the Fingerprints
2.1.4. Specificity of the Fingerprints
2.2. Comparison between Different Fingerprinting Methods
2.3. Identification of the Markers Based on the UHPLC–DAD–QOrbitrap–MS/MS Data
2.3.1. Flavonoids
2.3.2. Hydroxycoumarins
2.3.3. Glycerol Esters of Cinnamic Acid Derivatives
2.3.4. Other Cinnamic Acid Derivatives
2.3.5. Salicylate-like Phenolic Glycosides
2.3.6. Secoiridoids
2.3.7. Phenylethanoids
2.3.8. Benzoic Acid Glycosides
2.3.9. Triterpenoids and Triterpenoid Derivatives
2.3.10. Other Compounds
3. Materials and Methods
3.1. Chemicals
3.2. Plant Samples
3.3. UHPLC–QqQ Full Scan Screening and Fingerprinting Analysis
3.4. UHPLC–QOrbitrap–MS/MS Analysis
3.5. Data Analysis by MZmine 2
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Manninen, M.; Vesterinen, V.M.; Vainio, A.K.; Korhonen, H.; Karonen, M.; Salminen, J.P. Identification of Tree Species by Their Defense Compounds: A Study with Leaf Buds of White and Silver Birches. J. Chem. Educ. 2021, 98, 973–981. [Google Scholar] [CrossRef]
- Lahtinen, M.; Lempa, K.; Salminen, J.P.; Pihlaja, K. HPLC Analysis of Leaf Surface Flavonoids for the Preliminary Classification of Birch Species. Phytochem. Anal. 2006, 17, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Melnyk, N.; Pawłowska, K.A.; Ziaja, M.; Wojnowski, W.; Koshovyi, O.; Granica, S.; Bazylko, A. Characterization of Herbal Teas Containing Lime Flowers – Tiliae flos by HPTLC Method with Chemometric Analysis. Food Chem. 2021, 346, 128929. [Google Scholar] [CrossRef]
- Ding, S.; Dudley, E.; Plummer, S.; Tang, J.; Newton, R.P.; Brenton, A.G. Fingerprint Profile of Ginkgo biloba Nutritional Supplements by LC/ESI-MS/MS. Phytochemistry 2008, 69, 1555–1564. [Google Scholar] [CrossRef] [PubMed]
- Bertrams, J.; Müller, M.B.; Kunz, N.; Kammerer, D.R.; Stintzing, F.C. Phenolic Compounds as Marker Compounds for Botanical Origin Determination of German Propolis Samples Based on TLC and TLC-MS. J. Appl. Bot. Food Qual. 2013, 86, 143–153. [Google Scholar] [CrossRef]
- Giffen, J.E.; Lesiak, A.D.; Dane, A.J.; Cody, R.B.; Musah, R.A. Rapid Species-Level Identification of Salvias by Chemometric Processing of Ambient Ionisation Mass Spectrometry-Derived Chemical Profiles. Phytochem. Anal. 2017, 28, 16–26. [Google Scholar] [CrossRef]
- Lesiak, A.D.; Cody, R.B.; Dane, A.J.; Musah, R.A. Plant Seed Species Identification from Chemical Fingerprints: A High-Throughput Application of Direct Analysis in Real Time Mass Spectrometry. Anal. Chem. 2015, 87, 8748–8757. [Google Scholar] [CrossRef]
- Kharyuk, P.; Nazarenko, D.; Oseledets, I.; Rodin, I.; Shpigun, O.; Tsitsilin, A.; Lavrentyev, M. Employing Fingerprinting of Medicinal Plants by Means of LC-MS and Machine Learning for Species Identification Task. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- Alcalde-Eon, C.; García-Estévez, I.; Rivas-Gonzalo, J.C.; Rodríguez De La Cruz, D.; Escribano-Bailón, M.T. Anthocyanins of the Anthers as Chemotaxonomic Markers in the Genus Populus L. Differentiation between Populus nigra, Populus alba and Populus tremula. Phytochemistry 2016, 128, 35–49. [Google Scholar] [CrossRef] [Green Version]
- Abreu, I.N.; Ahnlund, M.; Moritz, T.; Albrectsen, B.R. UHPLC-ESI/TOFMS Determination of Salicylate-like Phenolic Gycosides in Populus tremula Leaves. J. Chem. Ecol. 2011, 37, 857–870. [Google Scholar] [CrossRef]
- Julkunen-Tiitto, R. Phenolic Constituents of Salix: A Chemotaxonomic Survey of Further Finnish Species. Phytochemistry 1989, 28, 2115–2125. [Google Scholar] [CrossRef]
- Vidakovic, V.; Novakovic, M.; Popovic, Z.; Jankovic, M.; Matic, R.; Teševic, V.; Bojovic, S. Significance of Diarylheptanoids for Chemotaxonomical Distinguishing between Alnus glutinosa and Alnus incana. Holzforschung 2017, 72, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Sołtys, A.; Galanty, A.; Podolak, I. Ethnopharmacologically Important but Underestimated Genus Sorbus: A Comprehensive Review. Phytochem. Rev. 2020, 19, 491–526. [Google Scholar] [CrossRef]
- Jensen, S.R.; Franzyk, H.; Wallander, E. Chemotaxonomy of the Oleaceae: Iridoids as Taxonomic Markers. Phytochemistry 2002, 60, 213–231. [Google Scholar] [CrossRef]
- Vihakas, M.; Pälijärvi, M.; Karonen, M.; Roininen, H.; Salminen, J.P. Rapid Estimation of the Oxidative Activities of Individual Phenolics in Crude Plant Extracts. Phytochemistry 2014, 103, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Rank, N.E.; Köpf, A.; Julkunen-Tiitto, R.; Tahvanainen, J. Host Preference and Larval Performance of the Salicylate-Using Leaf Beetle Phratora vitellinae. Ecology 1998, 79, 618–631. [Google Scholar] [CrossRef] [Green Version]
- Fan, L.; Tong, Q.; Dong, W.; Yang, G.; Hou, X.; Xiong, W.; Shi, C.; Fang, J.; Wang, W. Tissue Distribution, Excretion, and Metabolic Profile of Dihydromyricetin, a Flavonoid from Vine Tea (Ampelopsis grossedentata) after Oral Administration in Rats. J. Agric. Food Chem. 2017, 65, 4597–4604. [Google Scholar] [CrossRef]
- Abad-García, B.; Garmón-Lobato, S.; Berrueta, L.A.; Gallo, B.; Vicente, F. A Fragmentation Study of Dihydroquercetin Using Triple Quadrupole Mass Spectrometry and Its Application for Identification of Dihydroflavonols in Citrus Juices. Rapid Commun. Mass Spectrom. 2009, 23, 2785–2792. [Google Scholar] [CrossRef]
- Fanzone, M.; González-Manzano, S.; Pérez-Alonso, J.; Escribano-Bailón, M.T.; Jofré, V.; Assof, M.; Santos-Buelga, C. Evaluation of Dihydroquercetin-3-O-Glucoside from Malbec Grapes as Copigment of Malvidin-3-O-Glucoside. Food Chem. 2015, 175, 166–173. [Google Scholar] [CrossRef] [Green Version]
- Ziaja, M.; Pawłowska, K.A.; Józefczyk, K.; Pruś, A.; Stefańska, J.; Granica, S. UHPLC-DAD-MS/MS Analysis of Extracts from Linden Flowers (Tiliae flos): Differences in the Chemical Composition between Five Tilia Species Growing in Europe. Ind. Crops Prod. 2020, 154, 112691. [Google Scholar] [CrossRef]
- Zhu, H.; Poojary, M.M.; Andersen, M.L.; Lund, M.N. Effect of PH on the Reaction between Naringenin and Methylglyoxal: A Kinetic Study. Food Chem. 2019, 298, 125086. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, C.; Hofmann, T.; Vršanská, M.; Schlosserová, N.; Visi-Rajczi, E.; Voběrková, S.; Pásztory, Z. In Vitro Antioxidant and Antibacterial Activities with Polyphenolic Profiling of Wild Cherry, the European Larch and Sweet Chestnut Tree Bark. Eur. Food Res. Technol. 2021, 247, 2355–2370. [Google Scholar] [CrossRef]
- Mikulic-Petkovsek, M.; Stampar, F.; Veberic, R.; Sircelj, H. Wild Prunus Fruit Species as a Rich Source of Bioactive Compounds. J. Food Sci. 2016, 81, C1928–C1937. [Google Scholar] [CrossRef]
- Eyles, A.; Jones, W.; Riedl, K.; Cipollini, D.; Schwartz, S.; Chan, K.; Herms, D.A.; Bonello, P. Comparative Phloem Chemistry of Manchurian (Fraxinus mandshurica) and Two North American Ash Species (Fraxinus americana and Fraxinus pennsylvanica). J. Chem. Ecol. 2007, 33, 1430–1448. [Google Scholar] [CrossRef] [PubMed]
- Jensen, S.R.; Nielsen, B.J. A New Coumarin Fraxidin 8-O-β-d-Glucoside and 10-Hydroxyligstroside from Bark of Fraxinus exelsior. Phytochemistry 1976, 15, 221–223. [Google Scholar] [CrossRef]
- Okińczyc, P.; Widelski, J.; Szperlik, J.; Żuk, M.; Mroczek, T.; Skalicka-Woźniak, K.; Sakipova, Z.; Widelska, G.; Kuś, P.M. Impact of Plant Origin on Eurasian Propolis on Phenolic Profile and Classical Antioxidant Activity. Biomolecules 2021, 11, 68. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lin, Z.; Fang, J.; Liu, M.; Niu, Y.; Chen, S.; Wang, H. An On-Line High-Performance Liquid Chromatography-Diode-Array Detector-Electrospray Ionization-Ion-Trap-Time-of-Flight-Mass Spectrometry-Total Antioxidant Capacity Detection System Applying Two Antioxidant Methods for Activity Evaluation of the Edible Flowers from Prunus mume. J. Chromatogr. A 2015, 1414, 88–102. [Google Scholar] [CrossRef]
- Kammerer, B.; Kahlich, R.; Biegert, C.; Gleiter, C.H.; Heide, L. HPLC-MS/MS Analysis of Willow Bark Extracts Contained in Pharmaceutical Preparations. Phytochem. Anal. 2005, 16, 470–478. [Google Scholar] [CrossRef]
- Meier, B.; Julkunen-Tiitto, R.; Tahvanainen, J.; Sticher, O. Comparative High-Performance Liquid and Gas-Liquid Chromatographic Determination of Phenolic Glucosides in Salicaceae Species. J. Chromatogr. A 1988, 442, 175–186. [Google Scholar] [CrossRef]
- Tóth, G.; Barabás, C.; Tóth, A.; Kéry, Á.; Béni, S.; Boldizsár, I.; Varga, E.; Noszál, B. Characterization of Antioxidant Phenolics in Syringa vulgaris L. Flowers and Fruits by HPLC-DAD-ESI-MS. Biomed. Chromatogr. 2016, 30, 923–932. [Google Scholar] [CrossRef]
- Wozniak, M.; Michalak, B.; Wyszomierska, J.; Dudek, M.K.; Kiss, A.K. Effects of Phytochemically Characterized Extracts from Syringa vulgaris and Isolated Secoiridoids on Mediators of Inflammation in a Human Neutrophil Model. Front. Pharmacol. 2018, 9, 349. [Google Scholar] [CrossRef]
- Geoffroy, T.R.; Stevanovic, T.; Fortin, Y.; Poubelle, P.E.; Meda, N.R. Metabolite Profiling of Two Maple-Derived Products Using Dereplication Based on High-Performance Liquid Chromatography-Diode Array Detector-Electrospray Ionization-Time-of-Flight-Mass Spectrometry: Sugar Maple Bark and Bud Hot-Water Extracts. J. Agric. Food Chem. 2019, 67, 8819–8838. [Google Scholar] [CrossRef] [PubMed]
- Haddock, E.A.; Gupta, R.K.; Al-Shafi, S.M.K.; Haslam, E.; Magnolato, D. The Metabolism of Gallic Acid and Hexahydroxydiphenic Acid in Plants. Part 1. Introduction. Naturally Occurring Galloyl Esters. J. Chem. Soc. Perkin Trans. 1 1982, 2515–2524. [Google Scholar] [CrossRef]
- De Tommasi, N.; De Simone, F.; Pizza, C.; Mahmood, N.; Moore, P.S.; Conti, C.; Orsi, N.; Stein, M.L. Constituents of Eriobotrya japonica. A Study of Their Antiviral Properties. J. Nat. Prod. 1992, 55, 1067–1073. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.G.; Li, H.R.; Wang, L.Y.; Li, Y.H.; Lu, S.G.; Wen, X.F.; Wang, J.; Daikonya, A.; Kitanaka, S. Triterpenoids from Hippophae rhamnoides L. and Their Nitric Oxide Production-Inhibitory and DPPH Radical-Scavenging Activities. Chem. Pharm. Bull. 2007, 55, 15–18. [Google Scholar] [CrossRef] [Green Version]
- Kazmi, M.H.; Ahmed, E.; Hameed, S.; Malik, A.; Fatima, I.; Ashraf, M. Cashmirols A and B, New Lipoxygenase Inhibiting Triterpenes from Sorbus cashmiriana. Chem. Biodivers. 2009, 6, 1471–1476. [Google Scholar] [CrossRef]
- Sun, Y.; Feng, F.; Nie, B.; Cao, J.; Zhang, F. High Throughput Identification of Pentacyclic Triterpenes in Hippophae rhamnoides Using Multiple Neutral Loss Markers Scanning Combined with Substructure Recognition (MNLSR). Talanta 2019, 205, 120011. [Google Scholar] [CrossRef]
- Xu, J.P.; Xu, R.S.; Li, X.Y. Glycosides of a Cycloartane Sapogenin from Curculigo orchioides. Phytochemistry 1992, 31, 233–236. [Google Scholar] [CrossRef]
- Ren, X.; He, T.; Chang, Y.; Zhao, Y.; Chen, X.; Bai, S.; Wang, L.; Shen, M.; She, G. The Genus Alnus, a Comprehensive Outline of Its Chemical Constituents and Biological Activities. Molecules 2017, 22, 1383. [Google Scholar] [CrossRef] [Green Version]
- Aoki, T.; Ohta, S.; Aratani, S.; Hirata, T.; Suga, T. The Structures of Four Novel C31-Secodammarane-Type Triterpenoid Saponins from the Female Flowers of Alnus serrulatoides. J. Chem. Soc. Perkin Trans. 1 1982, 1399–1403. [Google Scholar] [CrossRef]
- Aoki, T.; Ohta, S.; Suga, T. Triterpenoids, Diarylheptanoids and Their Glycosides in the Flowers of Alnus Species. Phytochemistry 1990, 29, 3611–3614. [Google Scholar] [CrossRef]
- Chambers, M.C.; MacLean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A Cross-Platform Toolkit for Mass Spectrometry and Proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef] [PubMed]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular Framework for Processing, Visualizing, and Analyzing Mass Spectrometry-Based Molecular Profile Data. BMC Bioinformatics 2010, 11, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, O.D.; Sumner, S.J.; Li, S.; Barnes, S.; Du, X. One Step Forward for Reducing False Positive and False Negative Compound Identifications from Mass Spectrometry Metabolomics Data: New Algorithms for Constructing Extracted Ion Chromatograms and Detecting Chromatographic Peaks. Anal. Chem. 2017, 89, 8696–8703. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
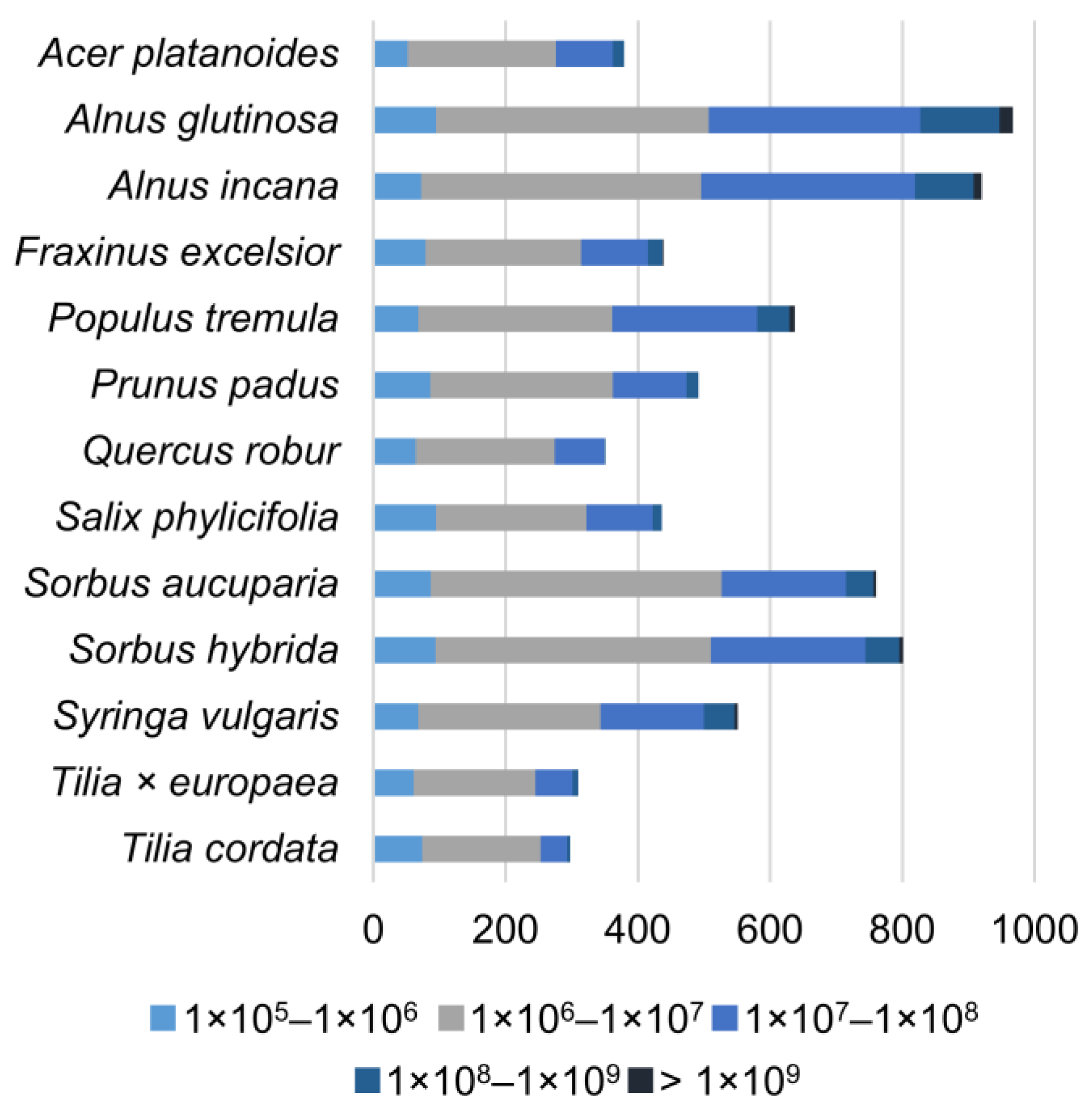{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method I (Species-Specific Ions) | Method II (Main Ions) | ||||||
|---|---|---|---|---|---|---|---|
| Species | Repeatability | Specificity | Repeatability | Specificity | Final Method | ||
| Intensity-Based | Profile-Based | Intensity-Based | Profile-Based | ||||
| Acer platanoides | 8/8 | OK | good | 8/8 | poor | poor | I |
| Alnus glutinosa | 6/8 * | good | good | 8/8 | good | good | II |
| Alnus incana | 8/8 | good | good | 8/8 | poor | poor | I |
| Fraxinus excelsior | 9/9 | good | good | 9/9 | poor | poor | I |
| Populus tremula | 10/10 | good | good | 10/10 | good | good | I |
| Prunus padus | 5/5 | good | good | 5/5 | poor | poor | I |
| Quercus robur | 9/9 | OK | good | 9/9 | poor | poor | I |
| Salix phylicifolia | 3/4 * | good | good | 4/4 | poor | poor | I |
| Sorbus aucuparia † | 9/9 | poor | good | 9/9 | good | good | II |
| Sorbus hybrida | 8/10 | good | good | 10/10 | good | good | II |
| Syringa vulgaris | 5/5 | good | good | 5/5 | poor | poor | I |
| Tilia cordata and Tilia × europaea | 15/15 | poor | good | 15/15 | good | poor | I |
| No. | Species | UV Max (nm) | Retention Time (min) | Molecular Formula | [M−H]− | m/z Values of Main Fragment Ions | Exact Mass, Calculated | Exact Mass, Measured | Error (ppm) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Acer platanoides | 212, 257, 300 (sh) | 0.40 | C15H20O10 | 359.09864 | 123, 182, 166, 197 | 360.10565 | 360.10594 | 0.29 |
| 2 | Fraxinus excelsior | nd. | 0.52 | * | 792.08522 | 191, 248, 263, 298, 354, 422, 438, 453, 468, 630 | |||
| 3 | Salix phylicifolia | 205, 290 | 0.66 | C15H12O8 | 319.04573 | 109, 125, 151, 193 | 320.05322 | 320.05303 | −0.19 |
| 4 | Tilia sp. | 230, 291 | 0.67 | C21H22O12 | 465.10378 | 107, 151, 285, 303 | 466.11113 | 466.11108 | −0.05 |
| 5 | Fraxinus excelsior | 205, 225 (sh), 329 | 0.67 | C18H22O12 | 429.10425 | 163, 191, 206, 221 | 430.11113 | 430.11155 | 0.42 |
| 6 | Acer platanoides | 258 | 0.70 | C22H24O14 | 511.1103 | 125, 169, 182, 197, 313, 345 | 512.11661 | 512.11760 | 0.99 |
| 7 | Acer platanoides | 258 | 0.79 | C22H24O14 | 511.11011 | 125, 169, 182, 197, 313, 345 | 512.11661 | 512.11741 | 0.8 |
| 8 | Acer platanoides | 258 | 0.88 | C22H24O14 | 511.11000 | 125, 169, 182, 197, 313, 345 | 512.11661 | 512.11730 | 0.69 |
| 9 | Fraxinus excelsior | 198, 230, 290 (sh), 328 | 0.94 | C23H26O11 | 477.14055 | 133, 161, 179, 315 | 478.14752 | 478.14785 | 0.335 |
| 10 | Syringa vulgaris | 195, 237, 282 (sh), 333 (sh) | 0.96 | C24H30O13 | 525.16106 | 121, 139, 165, 183, 209, 227, 249, 363, 389 | 526.16865 | 526.16836 | −0.285 |
| 11 | Acer platanoides | 225, 278 | 0.98 | C29H28O18 | 663.12130 | 125, 169, 313, 465, 511 | 664.12757 | 664.12860 | 1.03 |
| 12 | Fraxinus excelsior | 203, 231, 295 (sh), 330 | 1.00 | C23H26O11 | 477.14055 | 133, 161, 179, 315 | 478.14752 | 478.14785 | 0.335 |
| 13 | Sorbus sp. | nd. | 1.04 | C16H18O9 | 353.08783 | 135, 161, 179, 191 | 354.09509 | 354.09513 | 0.045 |
| 14 | Syringa vulgaris | 193, 237, 278 (sh), 333 (sh) | 1.10 | C24H30O12 | 509.16640 | 121, 139, 165, 233, 277, 347 | 510.17373 | 510.17370 | −0.03 |
| 15 | Acer platanoides | 221, 278 | 1.15 | C55H40O34 | 1243.13081 a | 125, 169, 393 b, 469 b, 617, 769 | 1244.14011 | 1244.13990 | −0.21 |
| 16 | Syringa vulgaris | nd. | 1.31 | C27H38O15 | 601.21324 | 153, 197 | 602.22108 | 602.22054 | −0.535 |
| 17 | Syringa vulgaris | 243, 278 (sh) | 1.44 | C32H38O14 | 645.21930 | 121, 139, 149, 165, 209, 275, 329, 413 | 646.22616 | 646.22660 | 0.44 |
| 18 | Prunus padus | 315 | 1.56 | C31H38O18 | 697.19783 | 117, 145, 163 | 698.20582 | 698.20513 | −0.69 |
| 19 | Populus tremula | 193, 221, 243, 297 (sh), 328 | 1.60 | C23H22O10 | 457.11453 | 133, 135, 161, 179, 235, 295, 397 | 458.12130 | 458.12183 | 0.53 |
| 20 | Salix phylicifolia | 240, 270 | 1.60 | C27H28O11 | 527.15605 | 93, 109, 121, 137, 155, 405 | 528.16317 | 528.16335 | 0.185 |
| 21 | Prunus padus | nd. | 1.75 | C28H26O11 | 537.14196 | 119, 151, 177, 271 | 538.14752 | 538.14926 | 1.745 |
| 22 | Populus tremula | 233, 290 (sh), 314 | 1.89 | C23H22O8 | 425.12500 | 93, 117, 119, 145, 163, 219, 365 | 426.13147 | 426.13230 | 0.83 |
| 23 | Populus tremula | 236, 296 (sh), 319 | 1.93 | C24H24O9 | 455.13543 | 117, 119, 134, 145, 160, 163, 175, 193, 395 | 456.14204 | 456.14273 | 0.695 |
| 24 | Populus tremula | 240, 293 (sh), 325 | 1.97 | C25H26O10 | 485.14578 | 134, 149, 160, 175, 193, 207, 425 | 486.15260 | 486.15308 | 0.48 |
| 25 | Alnus glutinosa | nd. | 2.35 | C35H58O10 | 637.39620 | 389, 477, 521, 605 | 638.40300 | 638.40350 | 0.5 |
| 26 | Alnus glutinosa | nd. | 2.41 | C35H58O10 | 637.39641 | 389, 477, 521, 605 | 638.40300 | 638.40371 | 0.71 |
| 27 | Sorbus sp. | 309 | 2.49 | C39H54O7 | 633.38042 | 117, 119, 145, 163, 571, 589, 615 | 634.38696 | 634.38772 | 0.765 |
| 28 | Tilia sp. | nd. | 2.64 | C13H28O8 | 311.16894 | 119, 183 | 312.17842 | 312.17624 | −2.18 |
| 29 | Sorbus sp. | 322 | 2.66 | C39H54O7 | 633.38031 | 133, 135, 161, 179 | 634.38696 | 634.38761 | 0.655 |
| 30 | Sorbus sp. | nd. | 2.67 | C30H46O4 | 469.33235 | 423 | 470.33961 | 470.33965 | 0.04 |
| 31 | Alnus glutinosa | nd. | 2.72 | C35H58O8 | 605.40623 | 283, 345, 455, 473 | 606.41317 | 606.41353 | 0.36 |
| 32 | Alnus glutinosa | nd. | 2.79 | C36H60O8 | 619.42097 | 343, 345, 371, 389, 487 | 620.42882 | 620.42827 | −0.55 |
| 33 | Alnus glutinosa | nd. | 2.82 | C35H58O8 | 605.40664 | 283, 345, 455, 473 | 606.41317 | 606.41394 | 0.77 |
| 34 | Alnus glutinosa | nd. | 2.88 | C36H60O8 | 619.42214 | 343, 345, 371, 389, 487 | 620.42882 | 620.42944 | 0.62 |
| 35 | Sorbus sp. | nd. | 2.93 | C32H48O5 | 511.34265 | 451 | 512.35018 | 512.34995 | −0.225 |
| 36 | Sorbus sp. | nd. | 3.00 | C30H46O3 | 453.33786 | nd. | 454.34470 | 454.34516 | 0.465 |
| 37 | Tilia sp. | nd. | 3.04 | C18H32O4 | 311.22300 | 111, 155, 171 | 312.23006 | 312.23030 | 0.24 |
| 38 | Sorbus sp. | nd. | 3.15 | C32H50O4 | 497.36354 | 437 | 498.37091 | 498.37084 | −0.07 |
| 39 | Quercus robur | nd. | 3.55 | C27H44O4 | 431.31652 | 133, 161, 179 | 432.32396 | 432.32382 | −0.14 |
| 40 | Alnus incana | nd. | 3.81 | C53H80O11 | 891.56326 | 117, 145, 163, 271, 331, 399, 417, 491, 745 | 892.57007 | 892.57056 | 0.495 |
| 41 | Alnus incana | nd. | 3.85 | C57H83O8 | 895.60993 | 117, 119, 145, 163, 749 | 896.61662 | 896.61723 | 0.61 |
| 42 | Alnus incana | nd. | 3.89 | C46H76O6 | 723.56173 | 117, 145, 223, 267, 285, 297, 455, 705 | 724.56419 | 724.56903 | 4.84 |
| Method I | Method II | |||||
|---|---|---|---|---|---|---|
| Species | m/z Value | Cone Voltage (V) | RT Range (min) | m/z Value | Cone Voltage (V) | RT Range (min) |
| Acer platanoides | 359.0 | 30 | 0.25–1.50 | 359.0 | 30 | 0.25–4.00 |
| 511.0 | 50 | 469.0 | 30 | |||
| 621.0 | 30 | 545.0 | 50 | |||
| 663.0 | 50 | 712.6 | 50 | |||
| Alnus glutinosa | 605.4 | 50 | 2.00–3.40 | 605.4 | 50 | 2.00–3.40 |
| 637.4 | 50 | 619.0 | 50 | |||
| 661.4 | 50 | 637.4 | 50 | |||
| Alnus incana | 723.5 | 70 | 3.40–4.00 | 439.4 | 30 | 2.40–4.00 |
| 891.5 | 70 | 453.4 | 30 | |||
| 895.6 | 70 | 471.4 | 30 | |||
| Fraxinus excelsior | 0.30–1.30 | 369.0 | 30 | 0.20–3.50 | ||
| 429.0 | 10 | 455.0 | 30 | |||
| 477.0 | 50 | 471.0 | 30 | |||
| 792.0 | 50 | 477.0 | 30 | |||
| 623.0 | 50 | |||||
| Populus tremula | 425.0 | 30 | 1.00–2.25 | 441.0 | 30 | 1.20–2.25 |
| 455.0 | 30 | 457.0 | 30 | |||
| 457.0 | 30 | 471.0 | 30 | |||
| 485.0 | 30 | |||||
| Prunus padus | 1.00–2.00 | 455.0 | 30 | 1.00–3.50 | ||
| 537.0 | 30 | 471.0 | 30 | |||
| 697.0 | 50 | 497.0 | 30 | |||
| 697.0 | 50 | |||||
| Quercus robur | 431.0 | 50 | 0.25–3.80 | 217.0 | 30 | 0.20–4.50 |
| 289.0 | 30 | |||||
| 431.0 | 50 | |||||
| 439.0 | 30 | |||||
| Salix phylicifolia | 0.40–1.90 | 217.0 | 30 | 0.30–4.50 | ||
| 319.0 | 30 | 301.0 | 30 | |||
| 527.0 | 30 | 503.4 | 30 | |||
| 712.5 | 30 | |||||
| Sorbus hybrida | 653.0 | 50 | 0.40–3.20 | |||
| 1045.6 | 50 | |||||
| Sorbusaucuparia and Sorbus hybrida | 0.60–2.80 | 353.0 | 30 | 0.80–3.50 | ||
| 453.4 | 30 | |||||
| 425.0 | 30 | 469.4 | 30 | |||
| 569.0 | 50 | 497.4 | 30 | |||
| 511.5 | 30 | |||||
| 633.5 | 30 | |||||
| Syringa vulgaris | 509.0 | 30 | 0.60–1.70 | 455.4 | 30 | 0.80–3.20 |
| 525.0 | 30 | 471.0 | 30 | |||
| 601.0 | 30 | 539.0 | 30 | |||
| 645.0 | 30 | |||||
| Tilia sp. | 0.50–3.25 | 217.0 | 30 | 0.25–4.50 | ||
| 311.0 | 30 | 289.0 | 30 | |||
| 465.0 | 30 | 475.4 | 30 | |||
| 503.6 | 30 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manninen, M.; Karonen, M.; Salminen, J.-P. Chemotaxonomic Markers for the Leaf Buds of Common Finnish Trees and Shrubs: A Rapid UHPLC MS Fingerprinting Tool for Species Identification. Molecules 2022, 27, 6810. https://doi.org/10.3390/molecules27206810
Manninen M, Karonen M, Salminen J-P. Chemotaxonomic Markers for the Leaf Buds of Common Finnish Trees and Shrubs: A Rapid UHPLC MS Fingerprinting Tool for Species Identification. Molecules. 2022; 27(20):6810. https://doi.org/10.3390/molecules27206810
Chicago/Turabian StyleManninen, Marianna, Maarit Karonen, and Juha-Pekka Salminen. 2022. "Chemotaxonomic Markers for the Leaf Buds of Common Finnish Trees and Shrubs: A Rapid UHPLC MS Fingerprinting Tool for Species Identification" Molecules 27, no. 20: 6810. https://doi.org/10.3390/molecules27206810