
Assessment of the Effects of Triticonazole on Soil and Human Health

 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Fungicides
2.2. Soil Sampling
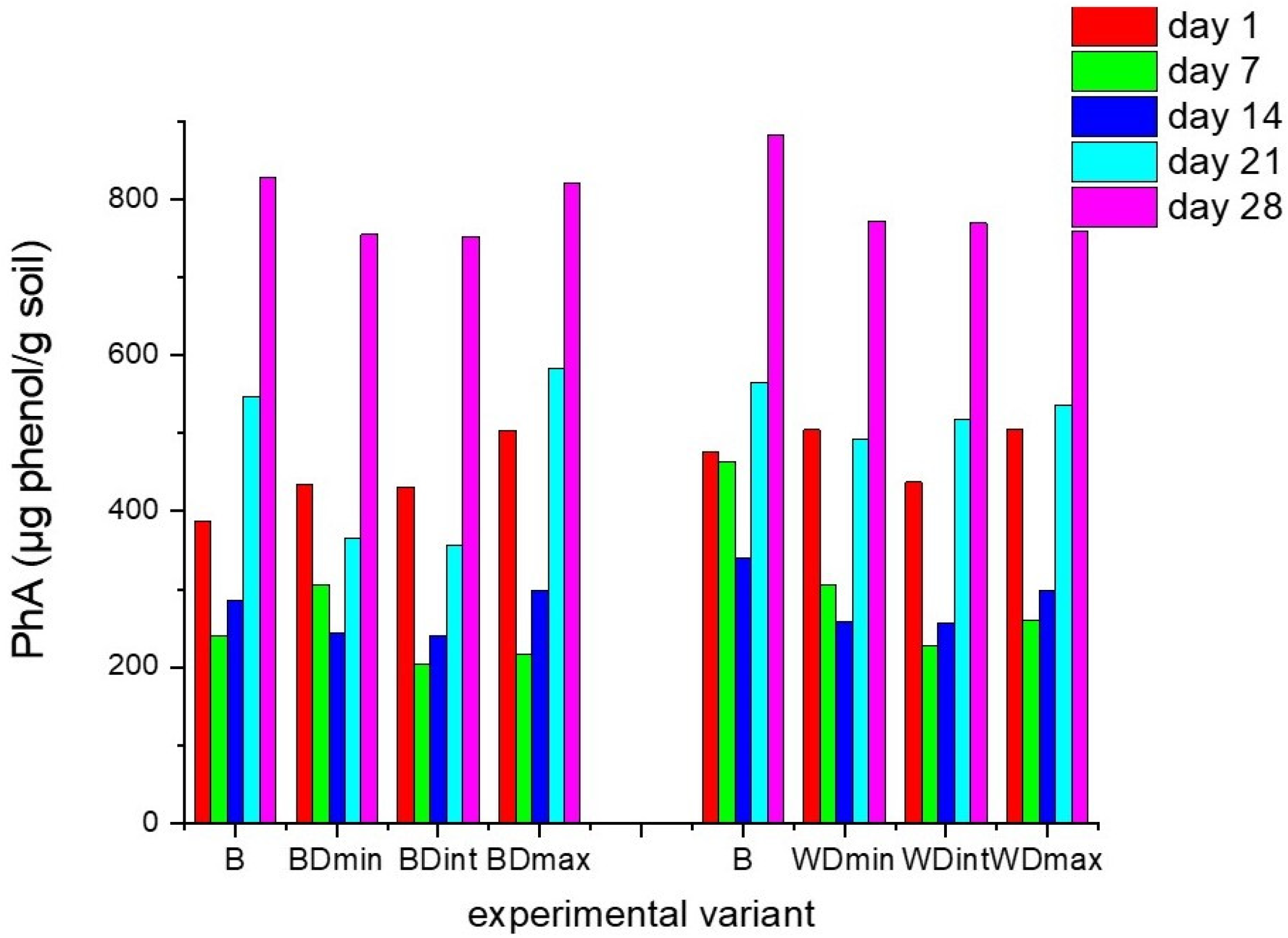
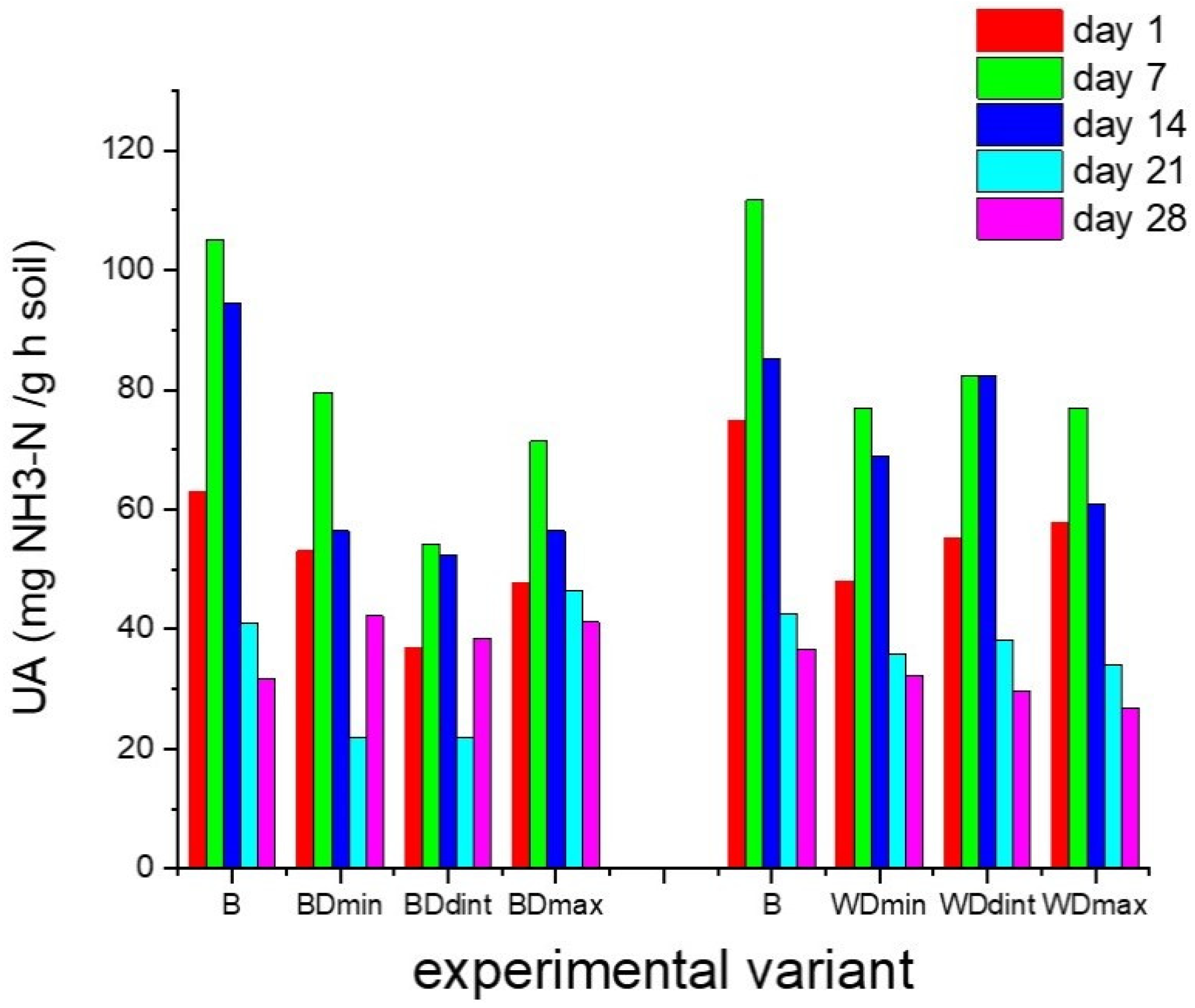
2.3. Enzymatic Activity Analyses
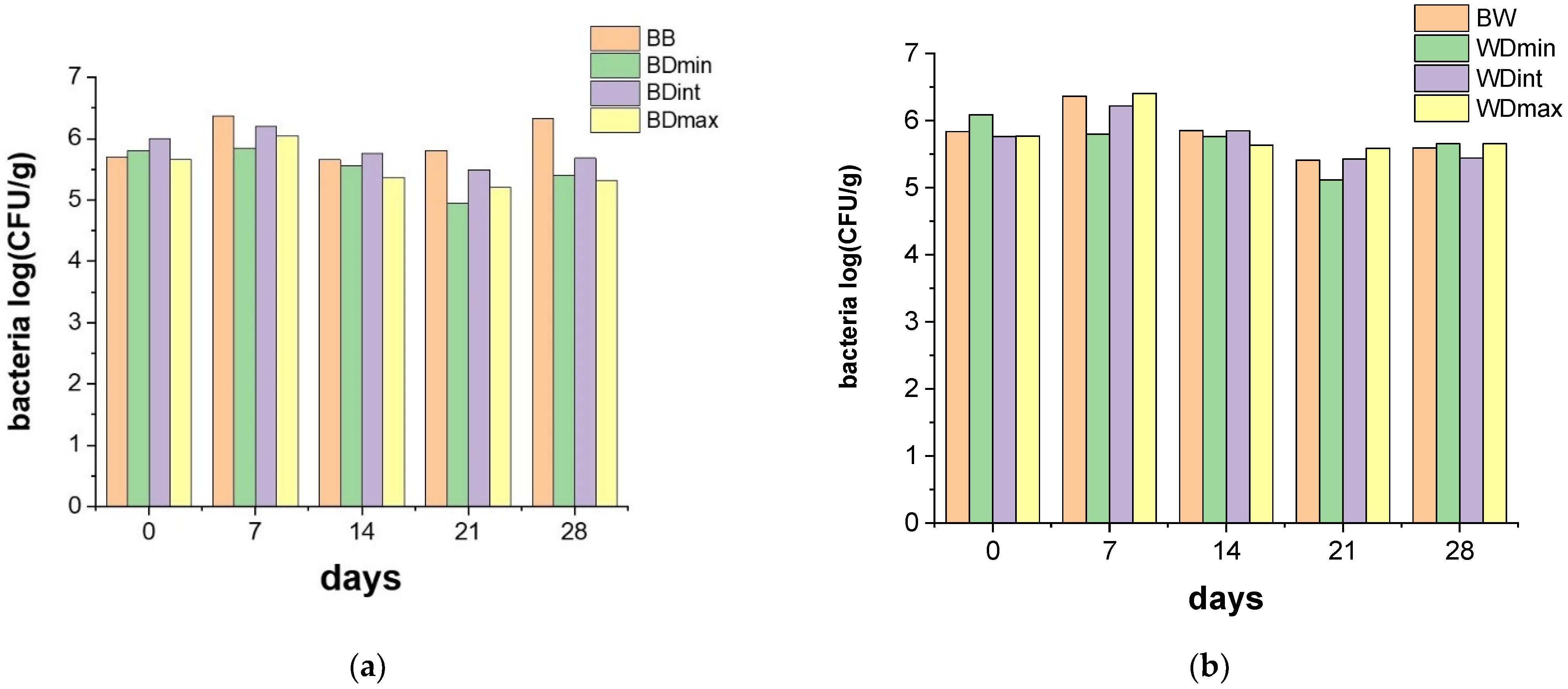
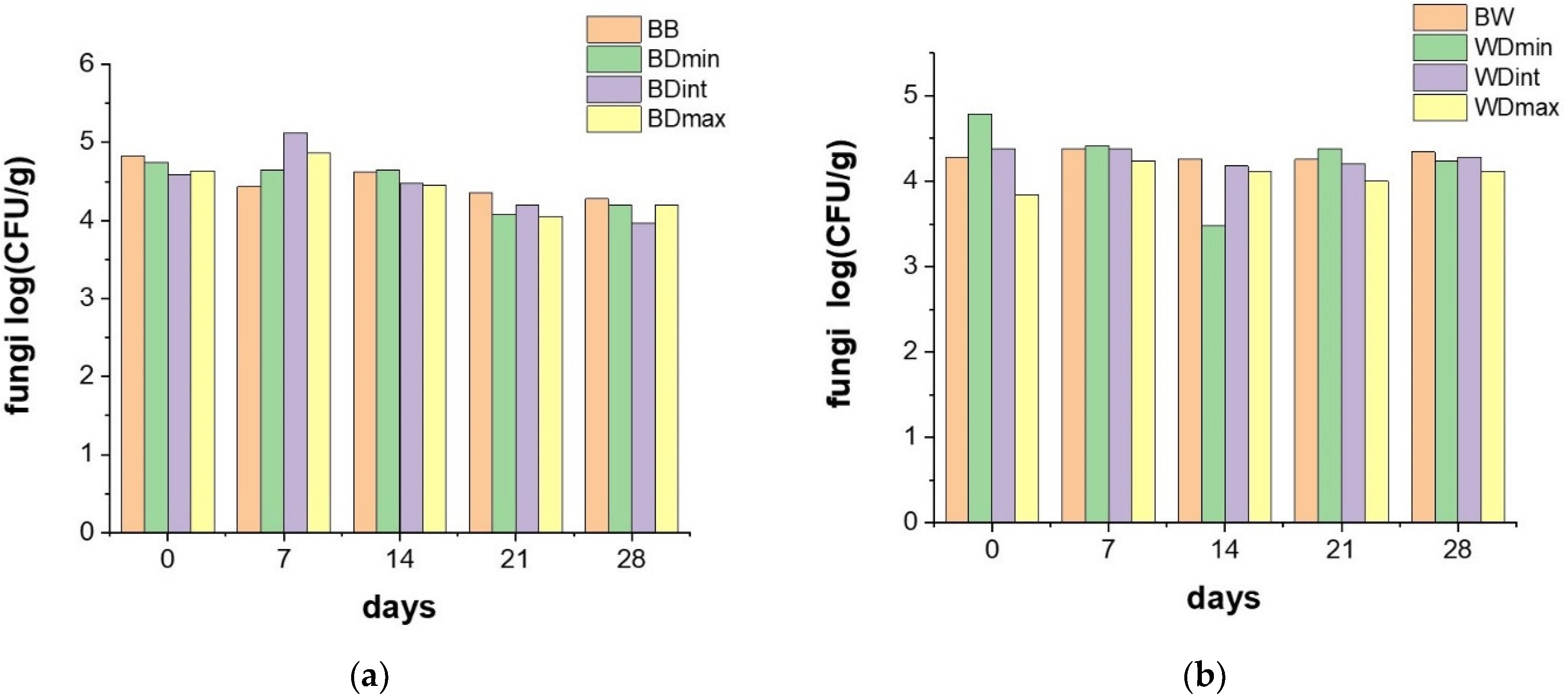
2.4. Microbiological Analysis
2.5. Prediction of the Effects on Human Health of the Stereoisomers of Triticonazole
2.6. Molecular Docking Study
2.7. Statistical Analysis
3. Results
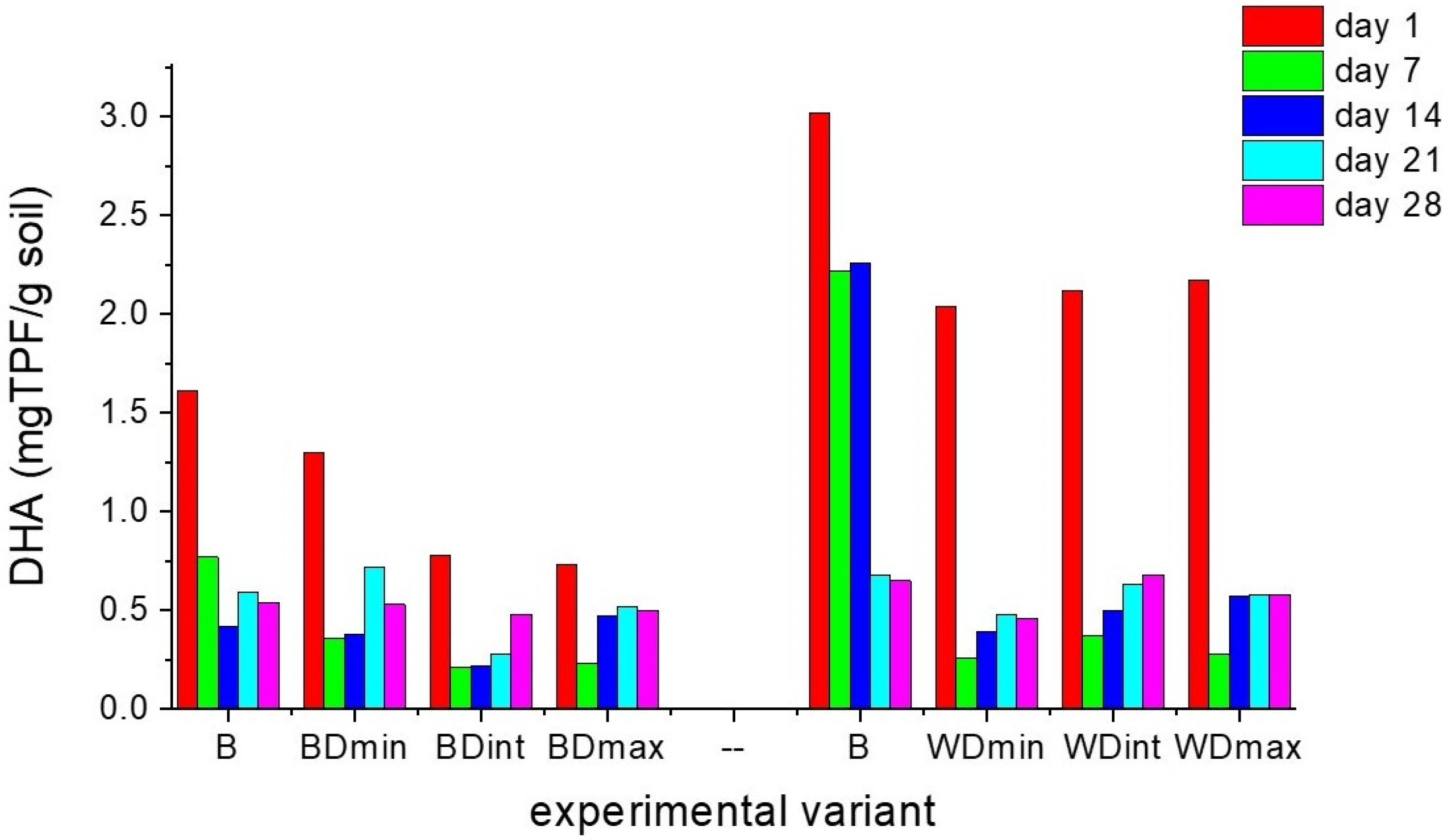
3.1. Effects of Triticonazole on the Activity of Enzymes Found in Soil
3.2. Effects of Triticonazole on the Soil Microorganisms
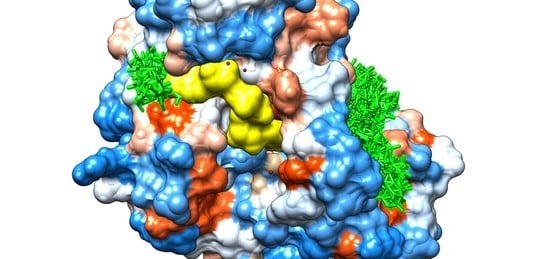
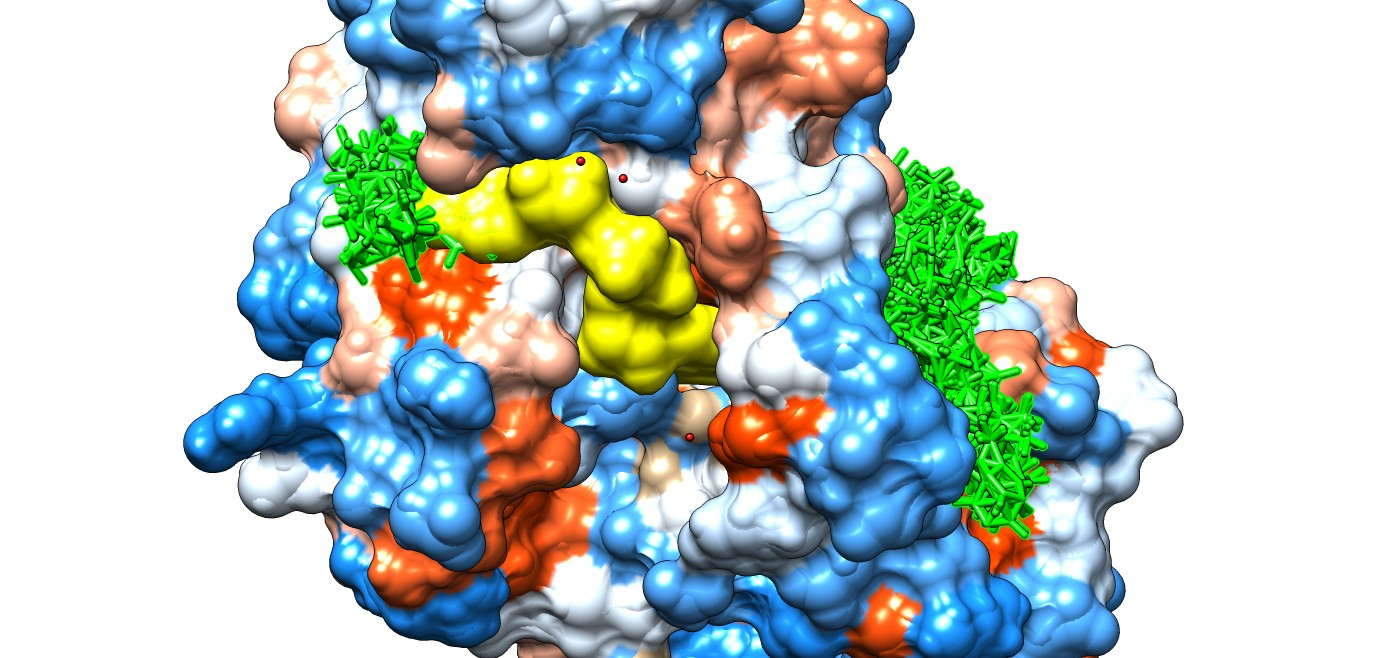
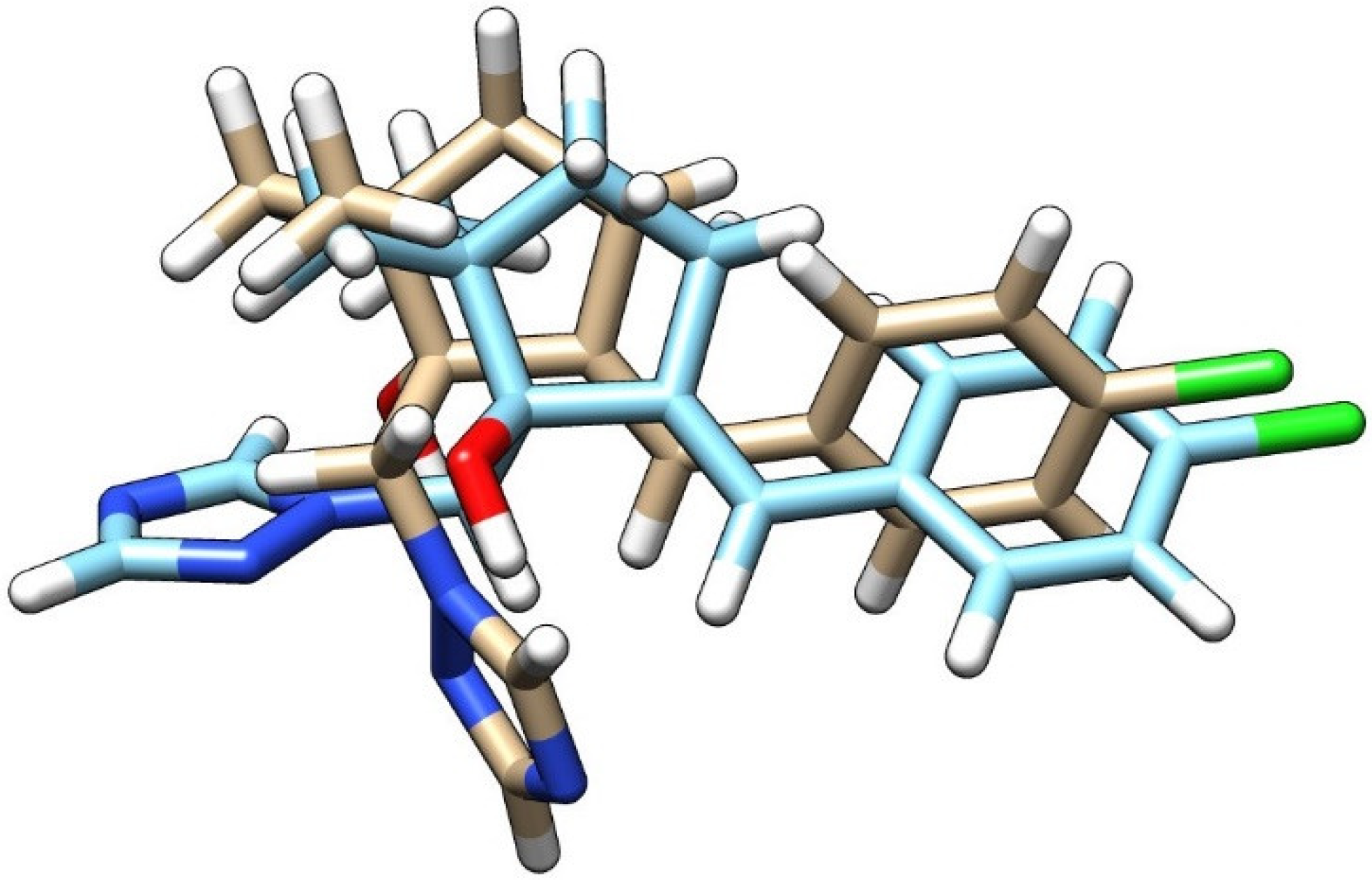
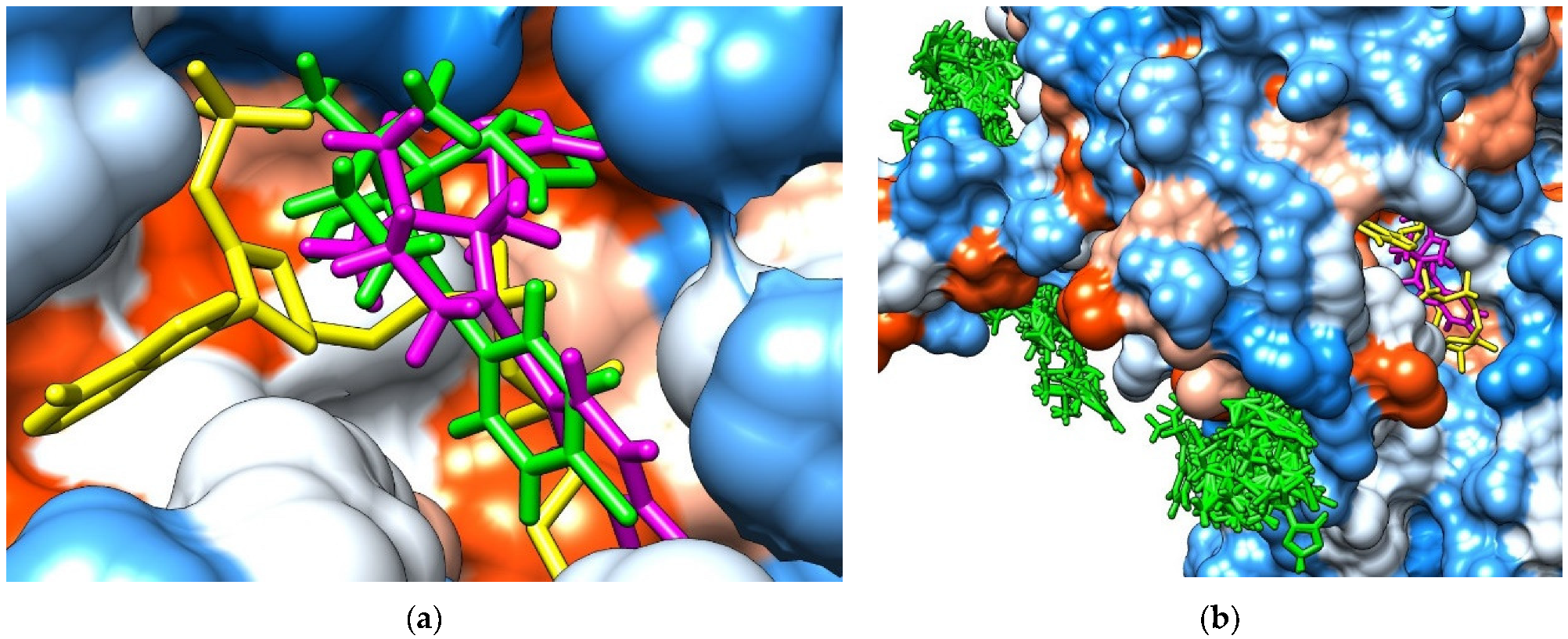
3.3. Molecular Docking Study Regarding the Interactions of the Enantiomers of Triticonazole with Soil Enzymes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Enzyme | PDB ID | Ligand in Crystallographic Structure | (R)-TTZ | (S)-TTZ | ||||
|---|---|---|---|---|---|---|---|---|
| Ligand | ΔG (kcal/mol) | Mean Value of ΔG (kcal/mol) | ΔG (kcal/mol) | Mean Value of ΔG (kcal/mol) | ΔG (kcal/mol) | Mean Value of ΔG (kcal/mol) | ||
| Dehydrogenases | 1KEV chain A | NADPH | −16.70 | −18.67 | −10.06 | −10.36 | −10.04 | −10.22 |
| 1KEV chain B | NADPH | −16.84 | −10.60 | −10.09 | ||||
| 1KEV chain C | NADPH | −20.98 | −9.99 | −10.38 | ||||
| 1KEV chain D | NADPH | −20.16 | −10.81 | −10.38 | ||||
| 3AUT chain A | NAI * | −10.32 | −10.42 | −7.21 | −7.31 | −7.70 | −7.58 | |
| 3AUT chain B | NAI | −10.51 | −7.41 | −7.46 | ||||
| 3NT5 chain A | ISE | −6.51 | −6.50 | −7.72 | −7.29 | −7.94 | −7.52 | |
| 3NT5 chain B | ISE | −6.59 | −6.86 | −7.09 | ||||
| 5GTL chain A | NADPH | −14.88 | −15.78 | it did not bind in the active site | −9.37 | −9.16 | −9.46 | |
| 5GTL chain B | NADPH | −17.11 | −9.46 | −9.94 | ||||
| 5GTL chain C | NADPH | −13.54 | −9.72 | −9.16 | ||||
| 5GTL chain D | NADPH | −14.57 | −8.93 | −9.58 | ||||
| Phophatases | 1H2F | AMP * | −8.89 | −8.89 | −8.19 | −8.19 | −8.37 | −8.37 |
| 1QFX chain A | phytate * | −10.22 | −10.94 | it did not bind in the active site | ||||
| 1QFX chain B | −11.65 | |||||||
| 2YEQ chain A | pNPP * | it did not bind in the active site | ||||||
| 2YEQ chain B | ||||||||
| Urease | 4AC7 chain C | urea * | −6.47 | −6.47 | it did not bind in the active site | |||
3.4. Predictions of the ADMET Profiles of the Enantiomers of Triticonazole
3.5. Molecular Docking Study Regarding the Interactions of the Enantiomers of Triticonazole with the Human Cytochromes and Plasma Proteins
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Jørgensen, L.N.; Heick, T.M. Azole use in agriculture, horticulture, and wood preservation—Is it indispensable? Front. Cell. Infect. Microbiol. 2021, 11, 730297. [Google Scholar] [CrossRef] [PubMed]
- Querou, R.; Euvrard, M.; Gauvrit, C. Uptake and fate of triticonazole applied as seed treatment to spring wheat (Triticum aestivum L.). Pestic. Sci. 1998, 53, 324–332. [Google Scholar] [CrossRef]
- Börjesson, E.; Stenström, J.; Johnsson, L.; Torstensson, L. Comparison of triticonazole dissipation after seed or soil treatment. J. Environ. Qual. 2003, 32, 1258–1261. [Google Scholar] [CrossRef] [PubMed]
- He, J.F.; Fan, J.; Yan, Y.L.; Chen, X.D.; Wang, T.; Zhang, Y.M.; Zhang, W.G. Triticonazole enantiomers: Separation by supercritical fluid chromatography and the effect of the chromatographic conditions. J. Sep. Sci. 2016, 39, 4251–4257. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, Z.X.; Tang, B.W.; Gao, B.B.; Tian, M.M.; Sanganyado, E.; Shi, H.Y.; Wang, M.H. Mechanistic insights into stereospecific bioactivity and dissipation of chiral fungicide triticonazole in agricultural management. J. Agric. Food Chem. 2018, 66, 7286–7293. [Google Scholar]
- Liu, R.; Zhang, H.; Deng, Y.; Zhou, Z.; Liu, X.; Diao, J. Enantioselective fungicidal activity and toxicity to early wheat growth of the chiral pesticide triticonazole. J. Agric. Food Chem. 2021, 69, 11154–11162. [Google Scholar] [CrossRef]
- Liu, R.; Deng, Y.; Zhang, W.; He, R.; Fan, J.; Zhu, W.; Zhiqiang, Z.; Jinling, D. Risk assessment of the chiral fungicide triticonazole: Enantioselective effects, toxicity, and fate. J. Agric. Food Chem. 2022, 70, 2712–2721. [Google Scholar] [CrossRef]
- Roman, D.L.; Ostafe, V.; Isvoran, A. Computational assessment of chito-oligosaccharides interactions with plasma proteins. Mar. Drugs 2021, 19, 120. [Google Scholar] [CrossRef]
- Roman, D.L.; Voiculescu, D.I.; Ostafe, V.; Ciorsac, A.; Isvoran, A. A review of the toxicity of triazole fungicides approved to be used in European Union to the soil and aqueous environment. Ovidius Univ. Ann. Chem. 2022, 33, 113–120. [Google Scholar] [CrossRef]
- Shen, Y.; Yao, X.; Jin, S.; Yang, F. Enantiomer/stereoisomer-specific residues of metalaxyl, napropamide, triticonazole, and metconazole in agricultural soils across China. Environ. Monit. Assess. 2021, 193, 773. [Google Scholar] [CrossRef]
- Lewis, K.A.; Tzilivakis, J.; Warner, D.J.; Green, A. An international database for pesticide risk assessments and management. Hum. Ecol. Risk. Assess. Int. J. 2016, 22, 1050–1064. [Google Scholar] [CrossRef]
- Charnay, M.P.; Tarabelli, L.; Beigel, C.; Barriuso, E. Modifications of soil microbial activity and triticonazole biodegradation by pesticide formulation additives. J. Environ. Qual. 2000, 29, 1618–1624. [Google Scholar] [CrossRef]
- Dakubo, C.Y. Exploring the Linkages Between Ecosystems and Human Health. In Ecosystems and Human Health: A Critical Approach to Ecohealth Research and Practice; Dabuko, C.Y., Ed.; Springer: New York, NY, USA, 2011; pp. 3–19. [Google Scholar]
- Mostafalou, S.; Abdollahi, M. Pesticides and human chronic diseases: Evidences, mechanisms, and perspectives. Toxicol. Appl. Pharmacol. 2013, 263, 157–177. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Wang, Z.; Gao, X.; Guo, X.; Zhao, L. Simultaneous enantioselective determination of 22 chiral pesticides in fruits and vegetables using chiral liquid chromatography coupled with tandem mass spectrometry. Food Chem. 2019, 277, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Tröger, R.; Klöckner, P.; Ahrens, L.; Wiberg, K. Micropollutants in drinking water from source to tap—Method development and application of a multiresidue screening method. Sci. Total Environ. 2018, 627, 1404–1432. [Google Scholar] [CrossRef] [PubMed]
- Draskau, M.K.; Boberg, J.; Taxvig, C.; Pedersen, M.; Frandsen, H.L.; Christiansen, S.; Svingen, T. In vitro and in vivo endocrine disrupting effects of the azole fungicides triticonazole and flusilazole. Environ. Pollut. 2019, 255, 113309. [Google Scholar] [CrossRef]
- Gridan, I.M.; Ciorsac, A.A.; Isvoran, A. Prediction of ADME-Tox properties and toxicological endpoints of triazole fungicides used for cereals protection. ADMET DMPK 2019, 7, 161–173. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.; Di, S.; Chen, X.; Wang, T.; Qi, P.; Wang, Z.; Zhao, H.; Gu, Y.; Xu, H.; Lu, Y.; et al. Evaluation of chiral triticonazole in three kinds of fruits: Enantioseparation, degradation, and dietary risk assessment. Environ. Sci. Pollut. Res. 2022, 29, 32855–32866. [Google Scholar] [CrossRef]
- Voiculescu, D.I.; Roman, D.L.; Ostafe, V.; Isvoran, A. A cheminformatics study regarding the human health risks assessment of the stereoisomers of difenoconazole. Molecules 2022, 27, 4682. [Google Scholar] [CrossRef]
- Isvoran, A.; Ciorsac, A.; Ostafe, V. ADME-Tox profiling of some low molecular weight water soluble chitosan derivatives. ADMET DMPK 2017, 5, 192–200. [Google Scholar] [CrossRef] [Green Version]
- Roman, M.; Roman, D.L.; Ostafe, V.; Ciorsac, A.; Isvoran, A. Computational assessment of pharmacokinetics and biological effects of some anabolic and androgen steroids. Pharm. Res. 2018, 35, 41. [Google Scholar] [CrossRef] [PubMed]
- Roman, D.L.; Roman, M.; Som, C.; Schmutz, M.; Hernandez, E.; Wick, P.; Casalini, T.; Perale, G.; Ostafe, V.; Isvoran, A. Computational assessment of the pharmacological profiles of degradation products of chitosan. Front. Bioeng. Biotechnol. 2019, 7, 214. [Google Scholar] [CrossRef] [PubMed]
- Roman, D.L.; Isvoran, A.; Filip, M.; Ostafe, V.; Zinn, M. In silico assessment of pharmacological profile of low molecular weight oligo-hydroxyalkanoates. Front. Bioeng. Biotechnol. 2020, 8, 584010. [Google Scholar] [CrossRef] [PubMed]
- Boros, B.V.; Dascalu, D.; Ostafe, V.; Isvoran, A. Assessment of the effects of chitosan, chitooligosaccharides and their derivatives on Lemna minor. Molecules 2022, 27, 6123, in press. [Google Scholar] [CrossRef]
- Craciun, D.; Modra, D.; Isvoran, A. ADME-Tox profiles of some food additives and pesticides. AIP Conf. Proc. 2015, 1694, 040007. [Google Scholar]
- Wu, F.; Zhou, Y.; Li, L.; Shen, X.; Chen, G.; Wang, X.; Liang, X.; Tan, M.; Huang, Z. Computational approaches in preclinical studies on drug discovery and development. Front. Chem. 2020, 8, 726. [Google Scholar] [CrossRef]
- Voiculescu, D.I.; Ostafe, V.; Isvoran, A. Computational assessment of the pharmacokinetics and toxicity of the intensive sweeteners. Farmacia 2021, 69, 1032–1041. [Google Scholar] [CrossRef]
- Craciun, D.; Dascalu, D.; Isvoran, A. Computational assessment of the ADME-Tox profiles and harmful effects of the most common used phthalates on the human health. Studia UBB Chemia 2019, 64, 71–92. [Google Scholar] [CrossRef]
- Alves, V.M.; Muratov, E.N.; Zakharov, A.; Muratov, N.N.; Andrade, C.H.; Tropsha, A. Chemical toxicity prediction for major classes of industrial chemicals: Is it possible to develop universal models covering cosmetics, drugs, and pesticides? Food Chem. Toxicol. 2018, 112, 526–534. [Google Scholar] [CrossRef]
- IUPAC Home—Triticonazole (Ref: RPA 400727). Available online: http://sitem.herts.ac.uk/aeru/iupac/Reports/673.htm (accessed on 7 May 2022).
- Sterling, T.; Irwin, J.J. ZINC 15—Ligand discovery for everyone. J. Chem. Inf. Model. 2015, 55, 2324–2337. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schinner, F.; Öhlinger, R.; Kandeler, E.; Margesin, R. Methods in Soil Biology; Springer: Berlin/Heidelberg, Germany, 1996; pp. 241–243. [Google Scholar]
- Alef, K.; Nannipieri, P. Methods in Applied Soil Microbiology and Biochemistry; Academic Press: London, UK, 1995; p. 316. [Google Scholar]
- Dick, R.P. Soil enzyme activities as integrative indicators of soil health. In Biological Indicators of Soil Health; Pankhurst, C.E., Doube, B.M., Gupta, V.V.S.R., Eds.; CAB International: Wallingford, UK, 1997; pp. 121–156. [Google Scholar]
- Filimon, M.N.; Roman, D.L.; Caraba, I.V.; Isvoran, A. Assessment of the effect of application of the herbicide S-metolachlor on the activity of some enzymes found in soil. Agriculture 2021, 11, 469. [Google Scholar] [CrossRef]
- Wollum, A.G. Chapter 37—Cultural methods for soil microorganisms. In Methods of Soil Analysis; Page, A.L., Ed.; John Willey&Sons: Hoboken, NJ, USA, 1983; pp. 781–802. [Google Scholar]
- Muñoz-Leoz, B.; Garbisu, C.; Charcosset, J.Y.; Sánchez-Pérez, J.M.; Antigüedad, I.; Ruiz-Romera, E. Non-target effects of three formulated pesticides on microbially-mediated processes in a clay-loam soil. Sci. Total. Environ. 2013, 449, 345–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filimon, M.N.; Voia, S.O.; Vlădoiu, D.L.; Isvoran, A.; Ostafe, V. Temperature dependent effect of difenoconazole on enzymatic activity from the soil. J. Serb. Chem. Soc. 2015, 80, 1127–1137. [Google Scholar] [CrossRef]
- Taxvig, C.; Hass, U.; Axelstad, M.; Dalgaard, M.; Boberg, J.; Andeasen, H.R.; Vinggaard, A.M. Endocrine-disrupting activities in vivo of the fungicides tebuconazole and epoxiconazole. Toxicol. Sci. 2007, 100, 464–473. [Google Scholar] [CrossRef] [Green Version]
- Kjaerstad, M.B.; Taxvig, C.; Nellemann, C.; Vinggaard, A.M.; Andersen, H.R. Endocrine disrupting effects in vitro of conazole antifungals used as pesticides and pharmaceuticals. Reprod. Toxicol. 2010, 30, 573–582. [Google Scholar] [CrossRef]
- Kolšek, K.; Mavri, J.; Sollner Dolenc, M.; Gobec, S.; Turk, S. Endocrine Disruptome—An open source prediction tool for assessing endocrine disruption potential through nuclear receptor binding. J. Chem. Inf. Model. 2014, 54, 1254–1267. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Moeskops, B.; Buchan, D.; Sleute, S.; Herawaty, L.; Husen, E.; Saraswati, R.; Setyorini, D.; De Neve, S. Soil microbial communities and activities under intensive organic and conventional vegetable farming in west java, Indonesia. Appl. Soil Ecol. 2010, 45, 112–120. [Google Scholar] [CrossRef]
- Dotaniya, M.L.; Aparna, K.; Dotaniya, C.K.; Singh, M.; Regar, K.L. Chapter 33—Role of soil enzymes in sustainable crop production. In Enzymes in Food Biotechnology; Kuddus, M., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 569–589. [Google Scholar]
- Skujip, S. Extracellular enzymes in soil. Crit. Rev. Microbiol. 1976, 4, 383–421. [Google Scholar]
- Korkhin, Y.; Frolow, F.; Bogin, O.; Peretz, M.; Kalb, A.J.; Burstein, Y. Crystalline alcohol dehydrogenases from the mesophilic bacterium Clostridium beijerinckii and the thermophilic bacterium Thermoanaerobium brockii: Preparation, characterization and molecular symmetry. Acta Crystallogr. D Biol. Crystallogr. 1996, 52, 882–886. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, T.; Yasutake, Y.; Nishiya, Y.; Tamura, T. Structure-guided mutagenesis for the improvement of substrate specificity of Bacillus megaterium glucose 1-dehydrogenase IV. FEBS J. 2012, 279, 3264–3275. [Google Scholar] [CrossRef] [PubMed]
- van Straaten, K.E.; Zheng, H.; Palmer, D.R.; Sanders, D.A. Structural investigation of myo-inositol dehydrogenase from Bacillus subtilis: Implications for catalytic mechanism and inositol dehydrogenase subfamily classification. Biochem. J. 2010, 432, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Benini, S.; Kosikowska, P.; Cianci, M.; Mazzei, L.; Gonzalez Vara, A.; Berlicki, Ł.; Ciurli, S. The crystal structure of Sporosarcina pasteurii urease in a complex with citrate provides new hints for inhibitor design. J. Biol. Inorg. Chem. 2013, 18, 391–399. [Google Scholar] [CrossRef]
- Rigden, D.J.; Littlejohn, J.E.; Henderson, K.; Jedrzejas, M.J. Structures of phosphate and trivanadate complexes of Bacillus stearothermophilus phosphatase phoe: Structural and functional analysis in the cofactor-dependent phosphoglycerate mutase superfamily. J. Mol. Biol. 2003, 325, 411. [Google Scholar] [CrossRef]
- Kostrewa, D.; Wyss, M.; D’Arcy, A.; van Loon, A.P. Crystal structure of Aspergillus niger pH 2. 5 acid phosphatase at 2. 4 A resolution. J. Mol. Biol. 1999, 288, 965–974. [Google Scholar]
- Rodriguez, F.; Lillington, J.; Johnson, S.; Timmel, C.R.; Lea, S.M.; Berks, B.C. Crystal structure of the Bacillus subtilis phosphodiesterase PhoD reveals an iron and calcium-containing active site. J. Biol. Chem. 2014, 289, 30889–30899. [Google Scholar] [CrossRef] [Green Version]
- Schönfeld, D.L.; Ravelli, R.B.G.; Mueller, U.; Skerra, A. The 1.8-Å crystal structure of α1-acid glycoprotein (orosomucoid) solved by UV RIP reveals the broad drug-binding activity of this human plasma lipocalin. J. Mol. Biol. 2008, 384, 393–405. [Google Scholar] [CrossRef]
- Zhang, Y.; Lee, P.; Liang, S.; Zhou, Z.; Wu, X.; Yang, F. Structural basis of non-steroidal anti-inflammatory drug diclofenac binding to human serum albumin. Chem. Biol. Drug Des. 2015, 86, 1178–1184. [Google Scholar] [CrossRef]
- Sansen, S.; Yano, J.K.; Reynald, R.L.; Schoch, G.A.; Griffin, K.J.; Stout, C.D.; Johnson, E.F. Adaptations for the oxidation of polycyclic aromatic hydrocarbons exhibited by the structure of human P450 1A2. J. Biol. Chem. 2007, 282, 14348–14355. [Google Scholar] [CrossRef] [Green Version]
- Williams, P.; Cosme, J.; Ward, A.; Angove, H.C.; Matak Vinkovic, D.; Jhoti, H. Crystal structure of human cytochrome P450 2C9 with bound warfarin. Nature 2003, 424, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Reynald, R.L.; Sansen, S.; Stout, C.D.; Johnson, E.F. Structural characterization of human cytochrome P450 2C19. Active site differences between P450s 2C8, 2C9, and 2C19. J. Biol. Chem. 2012, 287, 44581–44591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, A.; Stout, C.D.; Zhang, Q.; Johnson, E.F. Contributions of ionic interactions and protein dynamics to cytochrome P450 2D6 (CYP2D6) substrate and inhibitor binding. J. Biol. Chem. 2015, 290, 5092–5104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, P.A.; Cosme, J.; Vinkovic, D.M.; Ward, A.; Angove, H.C.; Day, P.J.; Vonrhein, C.; Tickle, I.J.; Jhoti, H. Crystal structures of human cytochrome P450 3A4 bound to metyrapone and progesterone. Science 2004, 305, 683–686. [Google Scholar] [CrossRef] [PubMed]
- Marín-Benito, J.M.; Herrero-Hernández, E.; Soledad Andrades, M.; Sánchez-Martín, M.J.; Rodríguez-Cruz, M.S. Effect of different organic amendments on the dissipation of linuron, diazinon and myclobutanil in an agricultural soil incubated for different time periods. Sci. Total Environ. 2014, 476–477, 611–621. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Wu, Y.; Zhang, X.; Zhu, Y. Persistence of myclobutanil and its impact on soil microbial biomass C and dehydrogenase enzyme activity in tea orchard soils, Eurasian. J. Soil Sci. 2017, 6, 106–113. [Google Scholar]
- Singh, N. Factors affecting triadimefon degradation in soils. J. Agric. Food Chem. 2005, 53, 70–75. [Google Scholar] [CrossRef]
- Madhuri, R.J.; Rangaswamy, V. Influence of selected fungicides on microbial population in groundnut (Arachis hypogeae L.) soils. Pollut. Res. 2003, 22, 205–212. [Google Scholar]
- Satapute, P.; Kamble, M.V.; Adhikari, S.S.; Jogaiah, S. Influence of triazole pesticides on tillage soil microbial populations and metabolic changes. Sci. Total. Environ. 2019, 651, 2334–2344. [Google Scholar] [CrossRef]
- Muñoz-Leoz, B.; Ruiz-Romera, E.; Antigüedad, I.; Garbisu, C. Tebuconazole application decreases soil microbial biomass and activity. Soil Biol. Biochem. 2011, 43, 2176–2183. [Google Scholar] [CrossRef]
- Saha, A.; Piparya, A.; Bhadury, D. Enzymatic activities and microbial biomass in peanut field soil as affected by the foliar application of tebuconazole. Environ. Earth. Sci. 2016, 75, 558. [Google Scholar] [CrossRef]
- Baćmaga, M.; Wyszkowska, J.; Kucharski, J. Biostimulation as a process aiding tebuconazole degradation in soil. J. Soils Sediments 2019, 19, 3728–3741. [Google Scholar] [CrossRef] [Green Version]
- Baćmaga, M.; Wyszkowska, J.; Kucharski, J. Response of soil microorganisms and enzymes to the foliar application of Helicur 250 EW fungicide on Horderum vulgare L. Chemosphere 2020, 242, 125163. [Google Scholar] [CrossRef] [PubMed]
- Vladoiu, D.L.; Filimon, M.N.; Ostafe, V.; Isvoran, A. Effects of herbicides and fungicides on the soil chitinolytic activity. A molecular docking approach. Ecol. Chem. Eng. S. 2015, 22, 439–450. [Google Scholar] [CrossRef] [Green Version]
- Rigden, D.J.; Bagyan, I.; Lamani, E.; Setlow, P.; Jedrzejas, M.J. A cofactor-dependent phosphoglycerate mutase homolog from Bacillus stearothermophilus is actually a broad specificity phosphatase. Protein Sci. 2001, 10, 1835–1846. [Google Scholar] [CrossRef] [Green Version]
- Wyss, M.; Brugger, R.; Kronenberger, A.; ReÂmy, R.; Fimbel, R.; Oesterhelt, G.; Lehmann, M.; van Loon, A.P.G.M. Biochemical characterization of fungal phytases (myo-inositol hexakisphosphate phosphoshydrolases): Catalytic properties. Appl. Environ. Microbiol. 1999, 65, 367–373. [Google Scholar] [CrossRef]
- Adasme, M.F.; Linnemann, K.L.; Bolz, S.N.; Kaiser, F.; Salentin, S.; Haupt, V.J.; Schroeder, M. PLIP 2021: Expanding the scope of the protein–ligand interaction profiler to DNA and RNA. Nucleic Acids Res. 2021, 49, W530–W534. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- Braga, R.C.; Alves, V.M.; Muratov, E.N.; Strickland, J.; Kleinstreuer, N.; Trospsha, A.; Andrade, C.H. Pred-Skin: A fast and reliable web application to assess skin sensitization effect of chemicals. J. Chem. Inf. Model. 2017, 57, 1013–1017. [Google Scholar] [CrossRef]
- Zhang, L.; Ai, H.; Chen, W.; Yin, Z.; Hu, H.; Zhu, J.; Zhao, J.; Zhao, Q.; Liu, H. CarcinoPred-EL: Novel models for predicting the carcinogenicity of chemicals using molecular fingerprints and ensemble learning methods. Sci. Rep. 2017, 7, 2118. [Google Scholar] [CrossRef] [Green Version]
- Gadaleta, D.; Mangiatordi, G.F.; Catto, M.; Carotti, A.; Nicolotti, O. Applicability domain for QSAR models: Where theory meets reality. Int. J. Quant. Struct. Prop. Relatsh. 2016, 1, 45–63. [Google Scholar] [CrossRef]
- Sanganyado, E. Regulatory Perspectives and Challenges in Risk Assessment of Chiral Pollutants. In Chiral Organic Pollutants: Monitoring and Characterization in Food and the Environment; Sanganyado, E., Munjanja, B.K., Nollet, L.M.L., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 301–320. [Google Scholar]
- EFSA (European Food Safety Authority); Bura, L.; Friel, A.; Magrans, J.O.; Parra-Morte, J.M.; Szentes, C. Guidance of EFSA on risk assessments for active substances of plant protection products that have stereoisomers as components or impurities and for transformation products of active substances that may have stereoisomers. EFSA J. 2019, 17, 5804. [Google Scholar]
- Basheer, A.A. Chemical chiral pollution: Impact on the society and science and need of the regulations in the 21st century. Chirality 2018, 30, 402–406. [Google Scholar] [CrossRef] [PubMed]







| Experimental Variant/Dose | Seeds Untreated with Triticonazole | Minimum Dose 0.3 L/1000 kg | Intermediate Dose 0.6 L/1000 kg | Maximum Dose 0.9 L/1000 kg |
|---|---|---|---|---|
| Soil sown with seeds of barley | BB | BDmin | BDint | BDmax |
| Soil sown with seeds of wheat | BW | WDmin | WDint | WDmax |
| Stereoisomer | HIA < 30% | Pgp Substrate | Pgp Inhibitor | BBB Permeation | PPB | CL (mL/min/kg) |
|---|---|---|---|---|---|---|
| (R)-TTZ | 0.004 | 0.008 | 0.056 | 0.333 | 94.66% | 8.518 |
| (S)-TTZ | 0.003 | 0.008 | 0.011 | 0.514 | 89.39% | 10.572 |
| Stereoisomer | CYP 1A2 s | CYP 1A2 i | CYP 2C19 s | CYP 2C19 i | CYP 2C9 s | CYP 2C9 i | CYP 2D6 s | CYP2 CD6 i | CYP 3A4 s | CYP 3A4 i |
|---|---|---|---|---|---|---|---|---|---|---|
| (R)-TTZ | 0.392 | 0.975 | 0.452 | 0.922 | 0.281 | 0.795 | 0.122 | 0.516 | 0.816 | 0.873 |
| (S)-TTZ | 0.481 | 0.976 | 0.484 | 0.894 | 0.104 | 0.564 | 0.066 | 0.414 | 0.718 | 0.790 |
| Stereoisomer | hERG | HT | AMES Mutagenicity | Skin Sensitization | Carcinogenicity | Eye Corrosion | Eye Irritation | Respiratory Toxicity |
|---|---|---|---|---|---|---|---|---|
| (R)-TTZ | 0.878 | 0.412 | 0.488 | 0.779 | 0.641 | 0.004 | 0.273 | 0.273 |
| (S)-TTZ | 0.894 | 0.522 | 0.232 | 0.635 | 0.882 | 0.004 | 0.165 | 0.796 |
| Nuclear Receptor | (R)-TTZ | (S)-TTZ |
|---|---|---|
| AR | ||
| AR an | ||
| ER α | ||
| ER α an | ||
| ER β | ||
| ER β an | ||
| GR | ||
| GR an | ||
| LXR α | ||
| LXR β | ||
| MR | ||
| PPAR α | ||
| PPAR β | ||
| PPAR γ | ||
| PR | ||
| RXR α | ||
| TR α | ||
| TR β |
| Human Protein | PDB ID | Ligand in Crystallographic Structure | (R)-TTZ | (S)-TTZ | |||
|---|---|---|---|---|---|---|---|
| ΔG (kcal/mol) | Mean Value of ΔG (kcal/mol) | ΔG (kcal/mol) | Mean Value of ΔG (kcal/mol) | ΔG (kcal/mol) | Mean Value of ΔG (kcal/mol) | ||
| Plasma protein | AGP | −6.14 | −6.14 | −7.36 | −7.36 | −7.78 | −7.78 |
| HSA chain A | −7.37 | −7.44 | −9.25 | −8.86 | −8.52 | −9.37 | |
| HSA chain I | −7.50 | −8.47 | −10.21 | ||||
| Cytochrome | CYP1A2 chain A | −8.62 | −8,62 | −8.51 | −8.51 | −8.83 | −8.83 |
| CYP2C9 chain A | −7.73 | −7.92 | −7.78 | −7.50 | −7.32 | 7.60 | |
| CYP2C9 chain B | −8.11 | −7.22 | −7.88 | ||||
| CYP2C19 chain A | −7.47 | −7.50 | −7.35 | −7.24 | −7.38 | −7.49 | |
| CYP2C19 chain B | −7.40 | −7.16 | −8.73 | ||||
| CYP2C19 chain C | −7.63 | −6.96 | −6.52 | ||||
| CYP2C19 chain D | −7.51 | −7.47 | −7.34 | ||||
| CYP2D6 chain A | −8.71 | −8.71 | It did not bind in the active site | ||||
| CYP3A4 chain A | −6.67 | −6.67 | −8.97 | −8.79 | −7.63 | −7.63 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roman, D.L.; Voiculescu, D.I.; Matica, M.A.; Baerle, V.; Filimon, M.N.; Ostafe, V.; Isvoran, A. Assessment of the Effects of Triticonazole on Soil and Human Health. Molecules 2022, 27, 6554. https://doi.org/10.3390/molecules27196554
Roman DL, Voiculescu DI, Matica MA, Baerle V, Filimon MN, Ostafe V, Isvoran A. Assessment of the Effects of Triticonazole on Soil and Human Health. Molecules. 2022; 27(19):6554. https://doi.org/10.3390/molecules27196554
Chicago/Turabian StyleRoman, Diana Larisa, Denisa Ioana Voiculescu, Mariana Adina Matica, Victor Baerle, Marioara Nicoleta Filimon, Vasile Ostafe, and Adriana Isvoran. 2022. "Assessment of the Effects of Triticonazole on Soil and Human Health" Molecules 27, no. 19: 6554. https://doi.org/10.3390/molecules27196554