
Akebia Saponin D Inhibits the Inflammatory Reaction by Inhibiting the IL-6-STAT3-DNMT3b Axis and Activating the Nrf2 Pathway

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. MTT Assay
2.4. Measurement of the PGE2, NO, IL-6 and TNF-α Levels
2.5. Real-Time PCR Analysis
2.6. Western Blot Analysis
2.7. Flow Cytometric Analysis
2.8. Transfection Assays
2.9. Statistical Analysis
3. Results
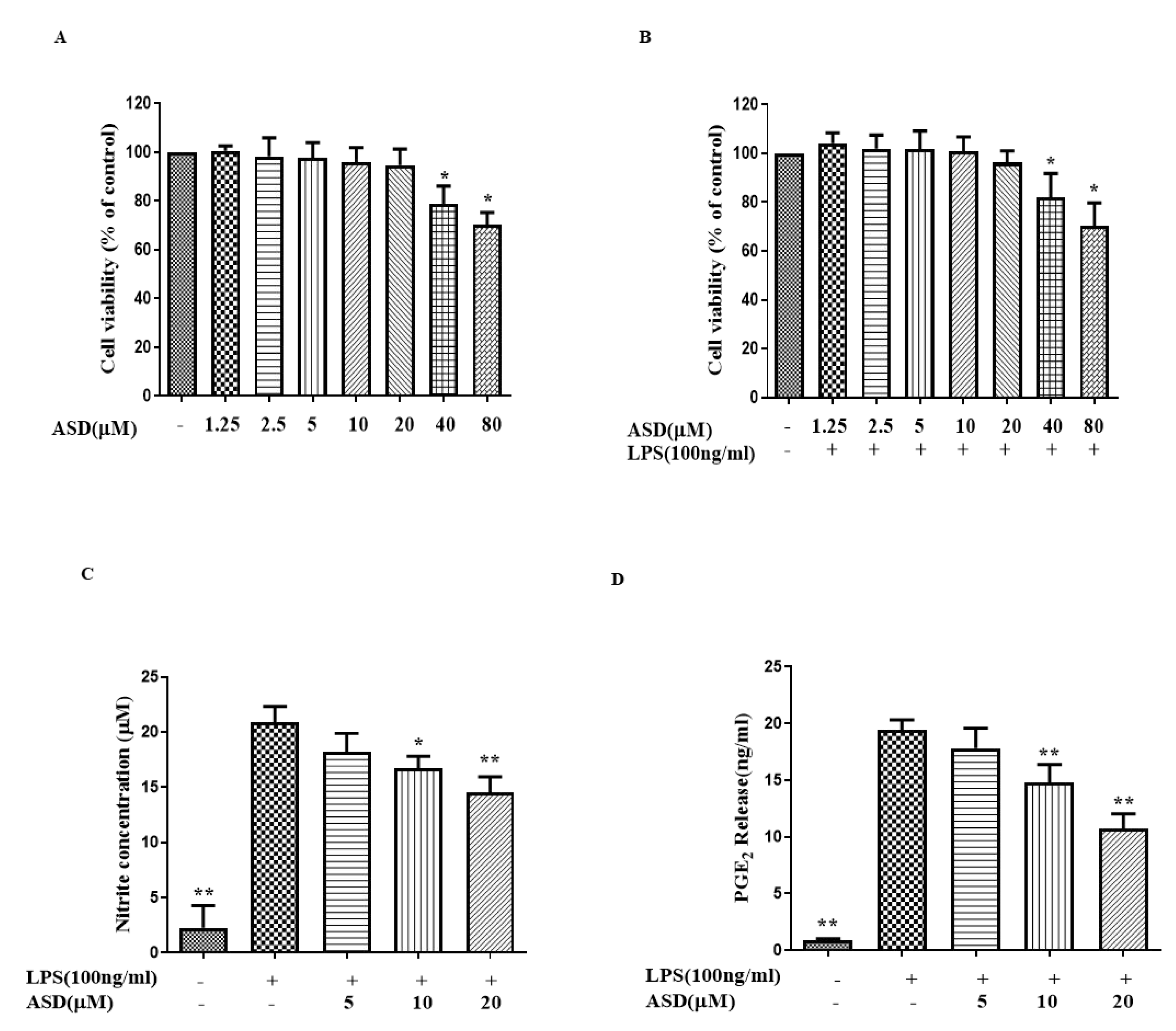
3.1. ASD Inhibited the Expression Levels of NO and PGE2 in LPS-Treated Macrophages
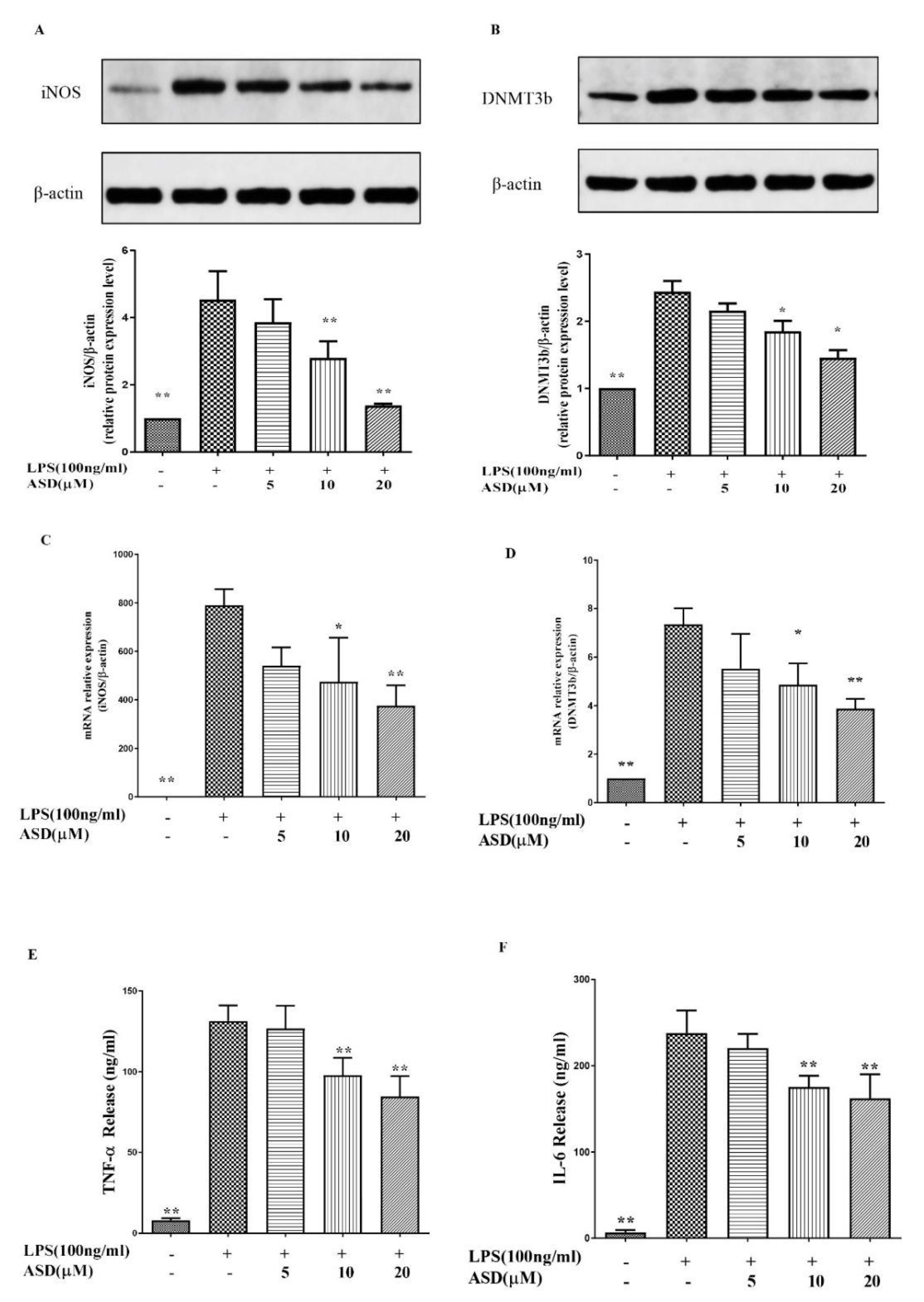
3.2. ASD Restrained the Levels of iNOS, DNMT3b, TNF-α, and IL-6
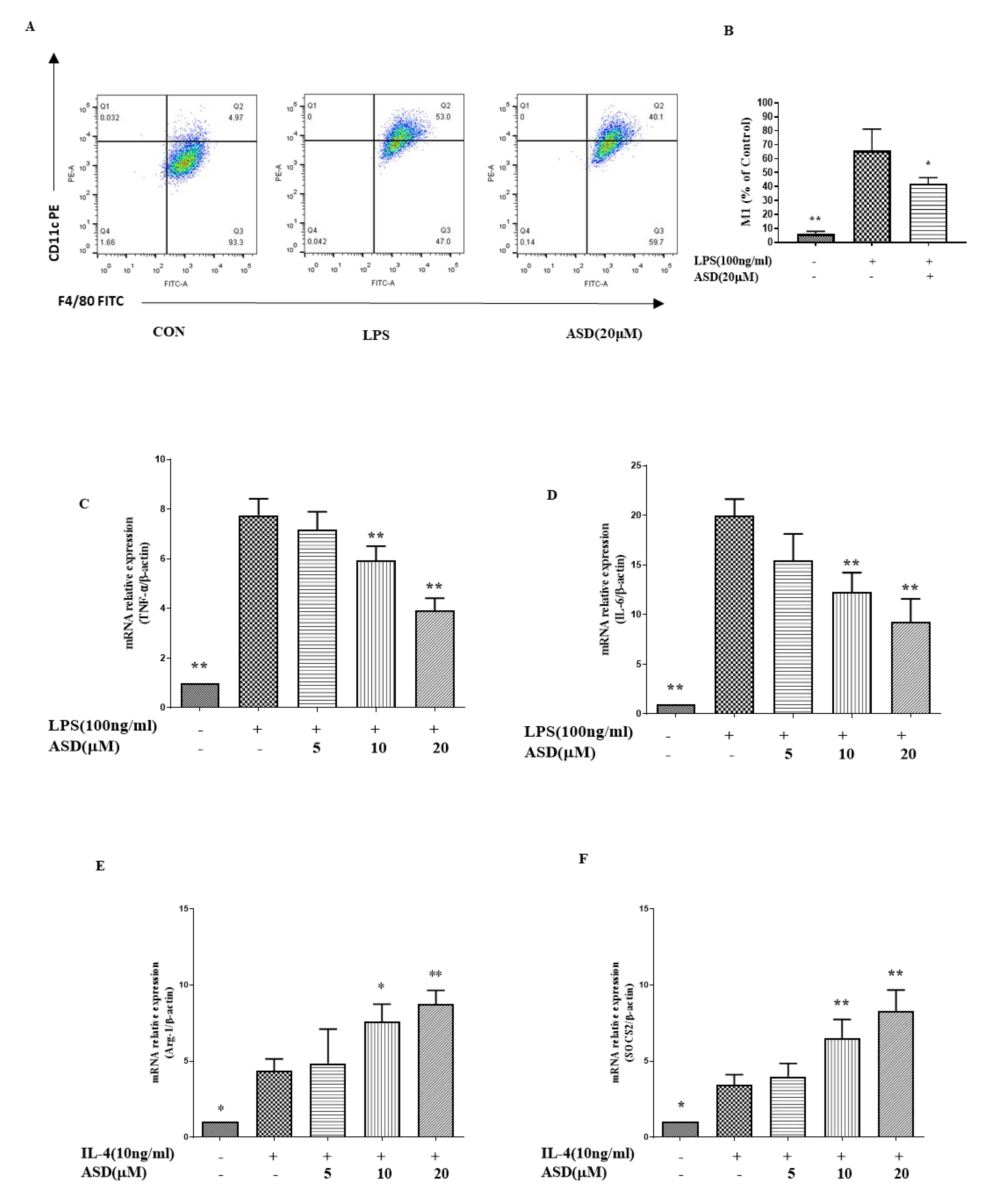
3.3. The Effect of ASD on the Expression Levels of M1, TNF-α, IL-6, Arg-1, and SOCS2
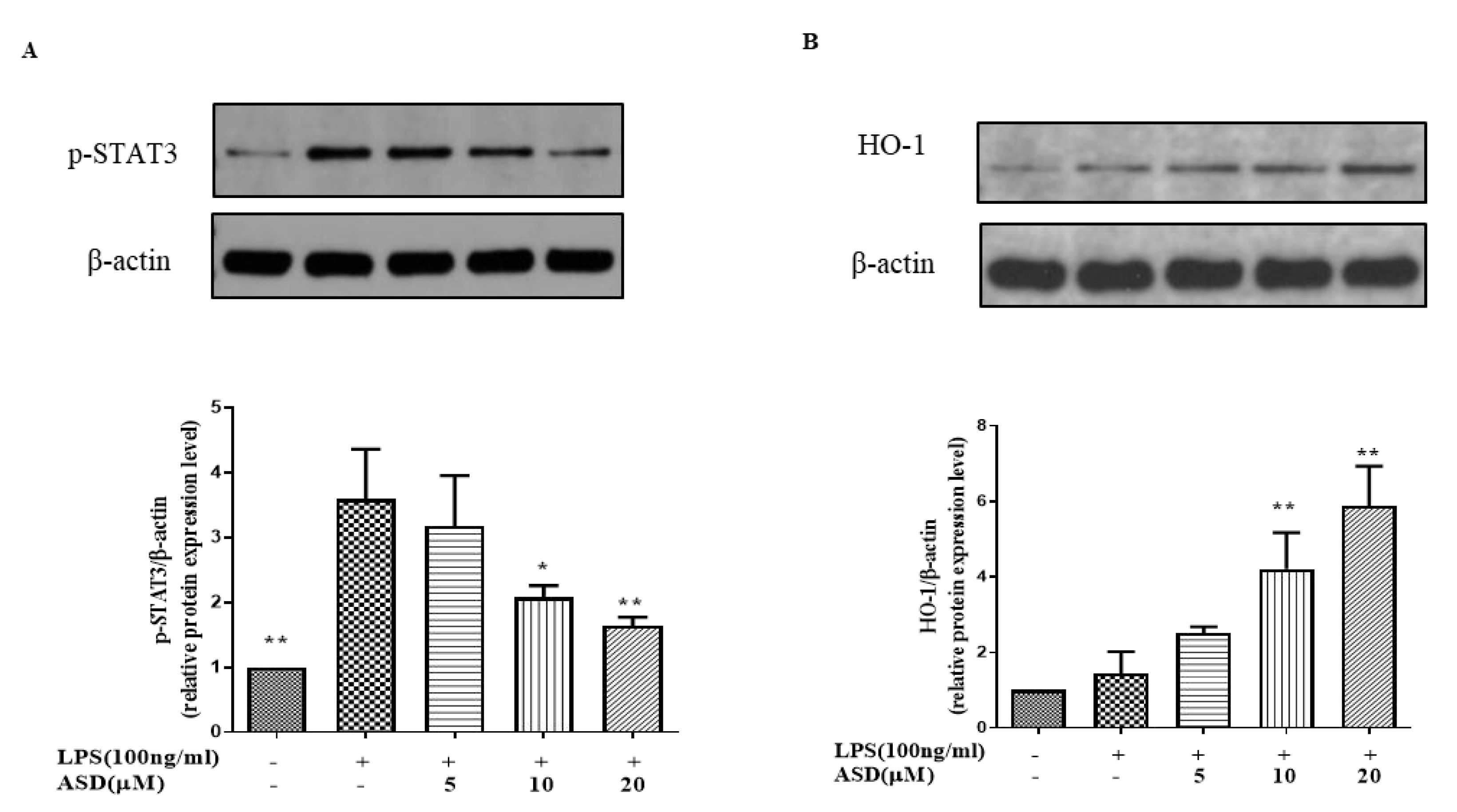
3.4. The Effect of ASD on the Levels of p-STAT3 and HO-1
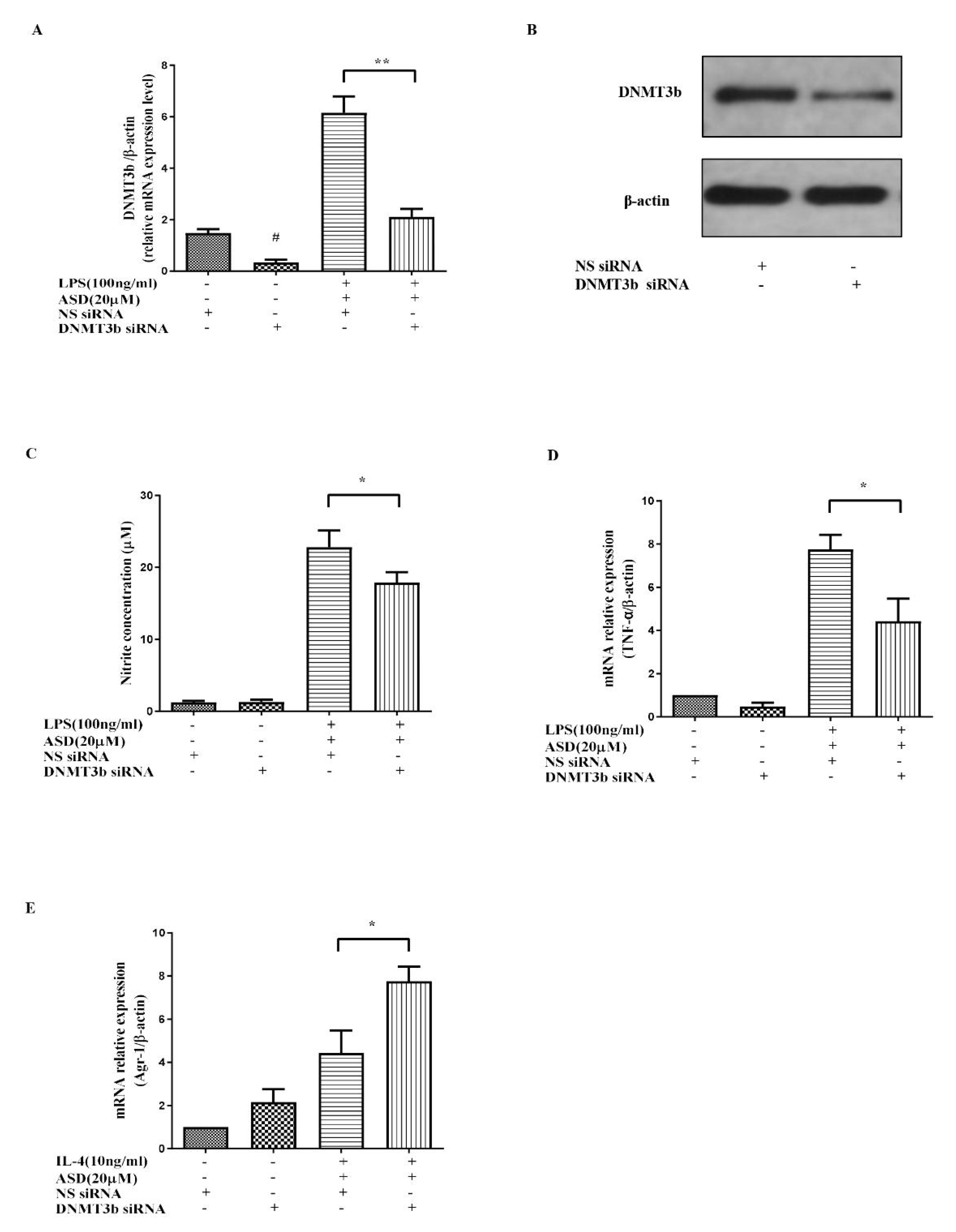
3.5. DNMT3b siRNA Significantly Promoted the Anti-Inflammatory Effect of ASD
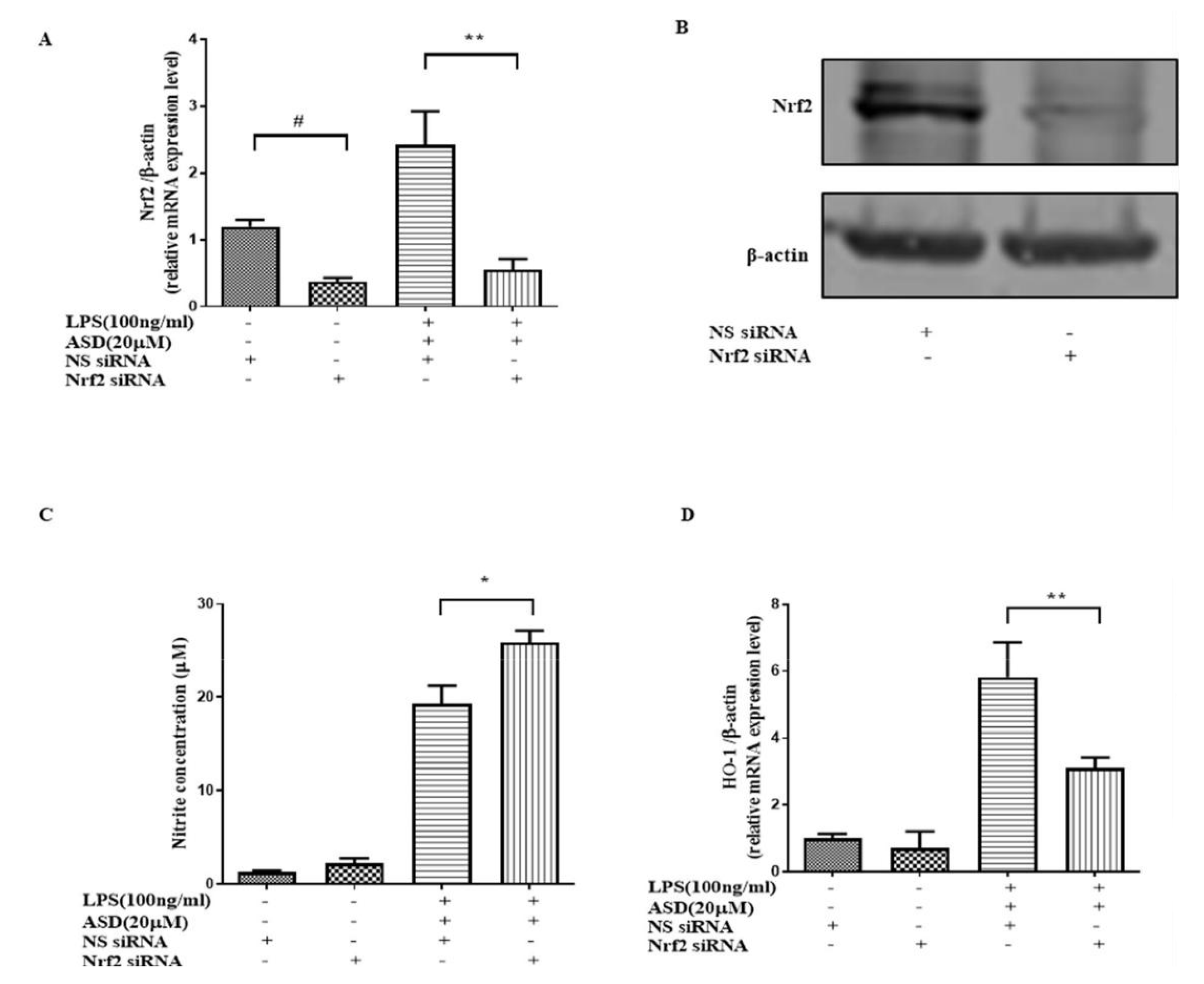
3.6. Nrf2 siRNA Significantly Abolished the Effect of ASD on NO and HO-1
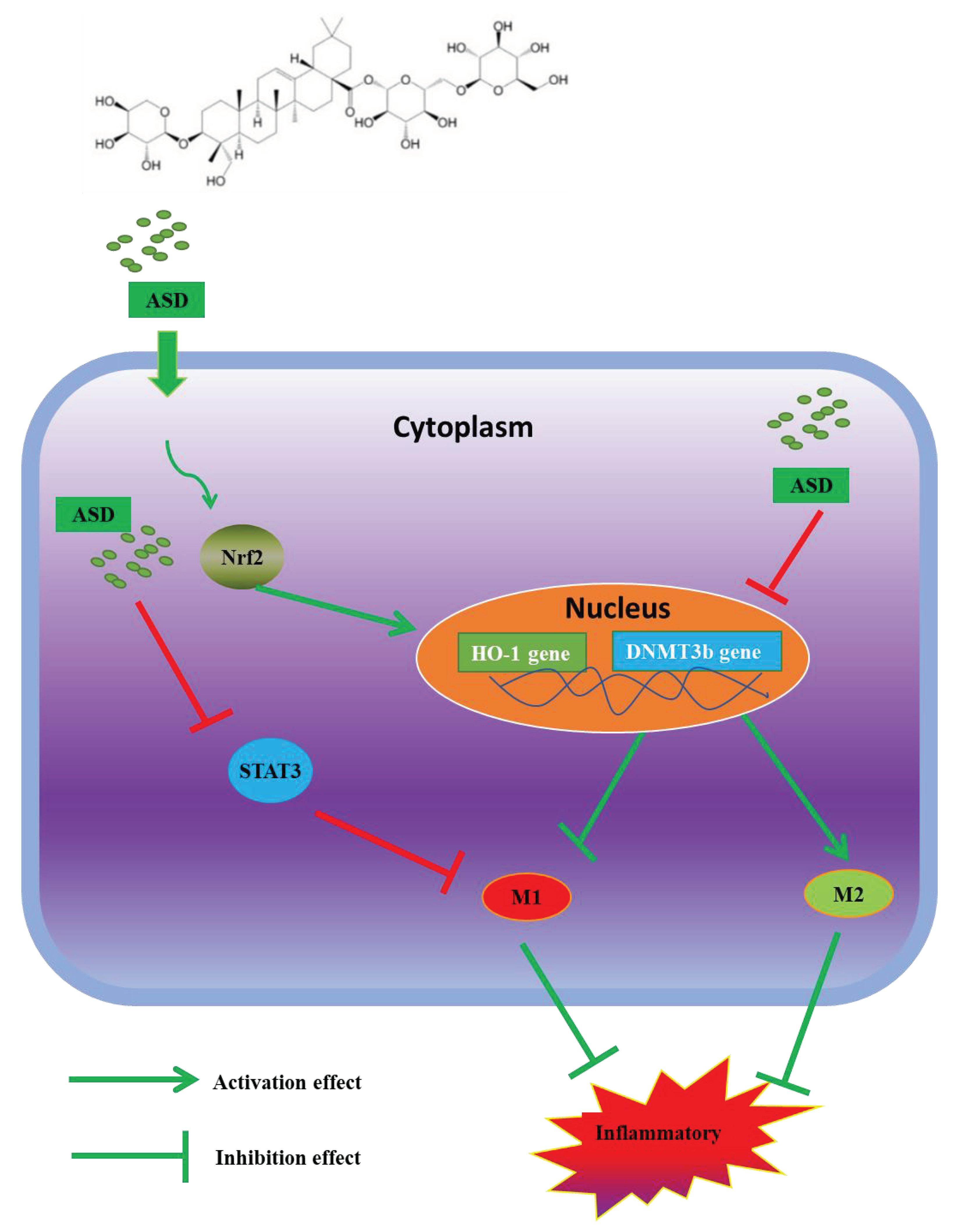
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Yang, Z.-H.; Dang, Y.-Q.; Ji, G. Role of epigenetics in transformation of inflammation into colorectal cancer. World J. Gastroenterol. 2019, 25, 2863. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Ma, H.; Wang, C.-Z.; Wan, J.-Y.; Yao, H.; Yuan, C.-S. Epigenetic Studies of Chinese Herbal Medicine: Pleiotropic Role of DNA Methylation. Front. Pharmacol. 2021, 12, 790321. [Google Scholar] [CrossRef] [PubMed]
- Man, X.; Li, Q.; Wang, B.; Zhang, H.; Zhang, S.; Li, Z. DNMT3A and DNMT3B in Breast Tumorigenesis and Potential Therapy. Front. Cell Dev. Biol. 2022, 1020, 916725. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.-F.; Yao, Y.-D.; Cheng, C.-S.; Lio, C.-K.; Liu, J.-X.; Huang, Y.-F.; He, F.; Xie, Y.; Liu, L.; Liu, Z.-Q. Sinomenine increases the methylation level at specific GCG site in mPGES-1 promoter to facilitate its specific inhibitory effect on mPGES-1. Biochimica Biophys. Acta-Gene Regul. Mech. 2022, 1865, 194813. [Google Scholar] [CrossRef] [PubMed]
- Stenvinkel, P.; Karimi, M.; Johansson, S.; Axelsson, J.; Suliman, M.; Lindholm, B.; Heimbürger, O.; Barany, P.; Alvestrand, A.; Nordfors, L. Impact of inflammation on epigenetic DNA methylation—A novel risk factor for cardiovascular disease? J. Intern. Med. 2007, 261, 488–499. [Google Scholar] [CrossRef]
- Yang, X.; Wang, X.; Liu, D.; Yu, L.; Xue, B.; Shi, H. Epigenetic regulation of macrophage polarization by DNA methyltransferase 3b. Mol. Endocrinol. 2014, 28, 565–574. [Google Scholar] [CrossRef]
- Niu, Y.; Chen, J.; Qiao, Y. Epigenetic Modifications in Tumor-Associated Macrophages: A New Perspective for an Old Foe. Front. Immunol. 2022, 13, 836223. [Google Scholar] [CrossRef]
- Moreira Lopes, T.C.; Mosser, D.M.; Gonçalves, R. Macrophage polarization in intestinal inflammation and gut homeostasis. Inflamm. Res. 2020, 69, 1163–1172. [Google Scholar] [CrossRef]
- Zhao, C.; Chen, H.; Liang, H.; Zhao, X.; Tang, W.; Wei, M.; Li, Y.; Zhang, J.; Yu, X.; Chen, G. Lactobacillus plantarum RS-09 Induces M1-Type Macrophage Immunity Against Salmonella Typhimurium Challenge via the TLR2/NF-κB Signalling Pathway. Front. Pharmacol. 2022, 13, 832245. [Google Scholar] [CrossRef]
- Naidoo, V.; Naidoo, M.; Ghai, M. Cell-and tissue-specific epigenetic changes associated with chronic inflammation in insulin resistance and type 2 diabetes mellitus. Scand. J. Immunol. 2018, 88, e12723. [Google Scholar] [CrossRef]
- Alkreathy, H.M.; Esmat, A. Lycorine Ameliorates Thioacetamide-Induced Hepatic Fibrosis in Rats: Emphasis on Antioxidant, Anti-Inflammatory, and STAT3 Inhibition Effects. Pharmaceuticals 2022, 15, 369. [Google Scholar] [CrossRef] [PubMed]
- Lai, S.-C.; Su, Y.-T.; Chi, C.-C.; Kuo, Y.-C.; Lee, K.-F.; Wu, Y.-C.; Lan, P.-C.; Yang, M.-H.; Chang, T.-S.; Huang, Y.-H. DNMT3b/OCT4 expression confers sorafenib resistance and poor prognosis of hepatocellular carcinoma through IL-6/STAT3 regulation. J. Exp. Clin. Cancer Res. 2019, 38, 474. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Barnes, G.T.; Yang, Q.; Tan, G.; Yang, D.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Investig. 2003, 112, 1821–1830. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, E.H.; Suzuki, T.; Funayama, R.; Nagashima, T.; Hayashi, M.; Sekine, H.; Tanaka, N.; Moriguchi, T.; Motohashi, H.; Nakayama, K. Nrf2 suppresses macrophage inflammatory response by blocking proinflammatory cytokine transcription. Nat. Commun. 2016, 7, 11624. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.-F.; Shen, X.-Y.; Lio, C.K.; Dai, Y.; Cheng, C.-S.; Liu, J.-X.; Yao, Y.-D.; Yu, Y.; Xie, Y.; Luo, P. Activation of Nrf2/HO-1 pathway by nardochinoid C inhibits inflammation and oxidative stress in lipopolysaccharide-stimulated macrophages. Front. Pharmacol. 2018, 9, 911. [Google Scholar] [CrossRef]
- Shen, T.; Chen, X.-M.; Harder, B.; Long, M.; Wang, X.-N.; Lou, H.-X.; Wondrak, G.T.; Ren, D.-M.; Zhang, D.D. Plant extracts of the family Lauraceae: A potential resource for chemopreventive agents that activate the nuclear factor-erythroid 2-related factor 2/antioxidant response element pathway. Planta Med. 2014, 80, 426–434. [Google Scholar] [CrossRef]
- Eräsalo, H.; Hämäläinen, M.; Leppänen, T.; Mäki-Opas, I.; Laavola, M.; Haavikko, R.; Yli-Kauhaluoma, J.; Moilanen, E. Natural stilbenoids have anti-inflammatory properties in vivo and down-regulate the production of inflammatory mediators NO, IL6, and MCP1 possibly in a PI3K/Akt-dependent manner. J. Nat. Prod. 2018, 81, 1131–1142. [Google Scholar] [CrossRef]
- Gong, L.-L.; Yang, S.; Liu, H.; Zhang, W.; Ren, L.-L.; Han, F.-F.; Lv, Y.-L.; Wan, Z.-R.; Liu, L.-H. Anti-nociceptive and anti-inflammatory potentials of Akebia saponin D. Eur. J. Pharmacol. 2019, 845, 85–90. [Google Scholar] [CrossRef]
- Ke, K.; Li, Q.; Yang, X.; Xie, Z.; Wang, Y.; Shi, J.; Chi, L.; Xu, W.; Hu, L.; Shi, H. Asperosaponin VI promotes bone marrow stromal cell osteogenic differentiation through the PI3K/AKT signaling pathway in an osteoporosis model. Sci. Rep. 2016, 6, 35233. [Google Scholar] [CrossRef]
- Ding, X.; Li, W.; Chen, D.; Zhang, C.; Wang, L.; Zhang, H.; Qin, N.; Sun, Y. Asperosaponin VI stimulates osteogenic differentiation of rat adipose-derived stem cells. Regen. Ther. 2019, 11, 17–24. [Google Scholar] [CrossRef]
- Zhen, D.; Xuan, T.-Q.; Hu, B.; Bai, X.; Fu, D.-N.; Wang, Y.; Wu, Y.; Yang, J.; Ma, Q. Pteryxin attenuates LPS-induced inflammatory responses and inhibits NLRP3 inflammasome activation in RAW264. 7 cells. J. Ethnopharmacol. 2022, 284, 114753. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, N.; Kobayashi, K. Macrophages in inflammation. Curr. Drug Targets-Inflamm. Allerg. 2005, 4, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-D.; Chen, X.; Xu, J.-J.; Du, X.-S.; Yang, Y.; Li, J.-J.; Yang, X.-J.; Huang, H.-M.; Li, X.-F.; Wu, M.-F. DNMT3b-mediated methylation of ZSWIM3 enhances inflammation in alcohol-induced liver injury via regulating TRAF2-mediated NF-κB pathway. Clin. Sci. 2020, 134, 1935–1956. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-W.; Sun, Y.-M. The IL-6/JAK/STAT3 pathway: Potential therapeutic strategies in treating colorectal cancer. Int. J. Oncol. 2014, 44, 1032–1040. [Google Scholar] [CrossRef]
- Linton, M.F.; Fazio, S. Macrophages, inflammation, and atherosclerosis. Int. J. Obes. 2003, 27, S35. [Google Scholar] [CrossRef]
- McInnes, I.B.; Schett, G. The pathogenesis of rheumatoid arthritis. N. Engl. J. Med. 2011, 365, 2205–2219. [Google Scholar] [CrossRef]
- Balkwill, F.R.; Mantovani, A. Cancer-related inflammation: Common themes and therapeutic opportunities. Semin. Cancer Biol. 2021, 22, 33–40. [Google Scholar] [CrossRef]
- Chawla, A.; Nguyen, K.D.; Goh, Y.S. Macrophage-mediated inflammation in metabolic disease. Nat. Rev. Immunol. 2011, 11, 738–749. [Google Scholar] [CrossRef]
- Kang, J.-K.; Chung, Y.-C.; Hyun, C.-G. Anti-Inflammatory Effects of 6-Methylcoumarin in LPS-Stimulated RAW 264.7 Macrophages via Regulation of MAPK and NF-κB Signaling Pathways. Molecules 2021, 26, 5351. [Google Scholar] [CrossRef]
- Park, M.Y.; Ha, S.E.; Kim, H.H.; Bhosale, P.B.; Abusaliya, A.; Jeong, S.H.; Park, J.-S.; Heo, J.D.; Kim, G.S. Scutellarein Inhibits LPS-Induced Inflammation through NF-κB/MAPKs Signaling Pathway in RAW264. 7 Cells. Molecules 2022, 27, 3782. [Google Scholar] [CrossRef]
- Thitinarongwate, W.; Nimlamool, W.; Khonsung, P.; Mektrirat, R.; Kunanusorn, P. Anti-Inflammatory Activity of Essential Oil from Zingiber ottensii Valeton in Animal Models. Molecules 2022, 27, 4260. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Kato, J.; Hiraoka, S.; Horii, J.; Suzuki, H.; Higashi, R.; Kaji, E.; Kondo, Y.; Yamamoto, K. DNA methylation of colon mucosa in ulcerative colitis patients: Correlation with inflammatory status. Inflamm. Bowel Dis. 2011, 17, 1955–1965. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Liang, H.; Zen, K. Molecular mechanisms that influence the macrophage M1–M2 polarization balance. Front. Immunol. 2014, 5, 614. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-B.; Lee, W.S.; Shin, J.-S.; Jang, D.S.; Lee, K.T. Xanthotoxin suppresses LPS-induced expression of iNOS, COX-2, TNF-α, and IL-6 via AP-1, NF-κB, and JAK-STAT inactivation in RAW 264.7 macrophages. Int. Immunopharmacol. 2017, 49, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.D.; Lu, C.; Payne, D.; Paschall, A.V.; Klement, J.D.; Redd, P.S.; Ibrahim, M.L.; Yang, D.; Han, Q.; Liu, Z. Autocrine IL6-Mediated Activation of the STAT3–DNMT Axis Silences the TNFα–RIP1 Necroptosis Pathway to Sustain Survival and Accumulation of Myeloid-Derived Suppressor Cells. Cancer Res. 2020, 80, 3145–3156. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.W.; Liu, X.; Macias, A.A.; Baron, R.M.; Perrella, M.A. Heme oxygenase-1–derived carbon monoxide enhances the host defense response to microbial sepsis in mice. J. Clin. Investig. 2008, 118, 239–247. [Google Scholar] [CrossRef]
- Ryter, S.W.; Alam, J.; Choi, A.M. Heme oxygenase-1/carbon monoxide: From basic science to therapeutic applications. Physiol. Rev. 2006, 86, 583–650. [Google Scholar] [CrossRef]
- Lin, H.-Y.; Juan, S.-H.; Shen, S.-C.; Hsu, F.-L.; Chen, Y.-C. Inhibition of lipopolysaccharide-induced nitric oxide production by flavonoids in RAW264. 7 macrophages involves heme oxygenase-1. Biochem. Pharmacol. 2003, 66, 1821–1832. [Google Scholar] [CrossRef]
- Wang, N.; Nie, H.; Zhang, Y.; Han, H.; Wang, S.; Liu, W.; Tian, K. Dexmedetomidine exerts cerebral protective effects against cerebral ischemic injury by promoting the polarization of M2 microglia via the Nrf2/HO-1/NLRP3 pathway. Inflamm. Res. 2022, 71, 93–106. [Google Scholar] [CrossRef]
- Fan, H.; Chen, W.; Zhu, J.; Zhang, J.; Peng, S. Toosendanin alleviates dextran sulfate sodium-induced colitis by inhibiting M1 macrophage polarization and regulating NLRP3 inflammasome and Nrf2/HO-1 signaling. Int. Immunopharmacol. 2019, 76, 105909. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Sequences |
|---|---|
| β-actin_F | 5′-CGGTTCCGATGCCCTGAGGCTCTT-3′ |
| β-actin_R | 5′-CGTCACACTTCATGATGGAATTGA-3′ |
| iNOS_F | 5′-CAGCACAGGAAATGTTTCAGC-3′ |
| iNOS_R | 5′-TAGCCAGCGTACCGGATGA-3′ |
| DNMT3b_F | 5′-TTCAGTGACCAGTCCTCAGACACGAA-3′ |
| DNMT3b_R | 5′-TCAGAAGGCTGGAGACCTCCCTCTT-3′ |
| TNF-α_F | 5′-TATGGCTCAGGGTCCAACTC-3′ |
| TNF-α_R | 5′-CTCCCTTTGCAGAACTCAGG-3′ |
| IL-6_F | 5′-GGTGACAACCACGGCCTTCCC-3′ |
| IL-6_R | 5′-AAGCCTCCGACTTGTGAAGTGGT-3′ |
| Nrf2_F | 5′-AGCAGGACATGGAGCAAGTT-3′ |
| Nrf2_R | 5′-TTCTTTTTCCAGCGAGGAGA-3′ |
| HO-1_F | 5′-CCCACCAAGTTCAAACAGCTC-3′ |
| HO-1_R | 5′-AGGAAGGCGGTCTTAGCCTC-3′ |
| Arg-1_F | 5′-AGCTCTGGGAATCTGCATGG-3′ |
| Arg-1_R | 5′-ATGTACACGATGTCTTTGGCAGATA-3′ |
| SOCS2_F | 5′-CTGCGCGAGCTCAGTCAAAC-3′ |
| SOCS2_R | 5′-CAAGAAAGTTCCTTCTGGAGCCTCT-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, J.-F.; Zhou, H.; Lio, C.-K. Akebia Saponin D Inhibits the Inflammatory Reaction by Inhibiting the IL-6-STAT3-DNMT3b Axis and Activating the Nrf2 Pathway. Molecules 2022, 27, 6236. https://doi.org/10.3390/molecules27196236
Luo J-F, Zhou H, Lio C-K. Akebia Saponin D Inhibits the Inflammatory Reaction by Inhibiting the IL-6-STAT3-DNMT3b Axis and Activating the Nrf2 Pathway. Molecules. 2022; 27(19):6236. https://doi.org/10.3390/molecules27196236
Chicago/Turabian StyleLuo, Jin-Fang, Hua Zhou, and Chon-Kit Lio. 2022. "Akebia Saponin D Inhibits the Inflammatory Reaction by Inhibiting the IL-6-STAT3-DNMT3b Axis and Activating the Nrf2 Pathway" Molecules 27, no. 19: 6236. https://doi.org/10.3390/molecules27196236