
Molecular Mechanistic Pathways Targeted by Natural Compounds in the Prevention and Treatment of Diabetic Kidney Disease

Abstract
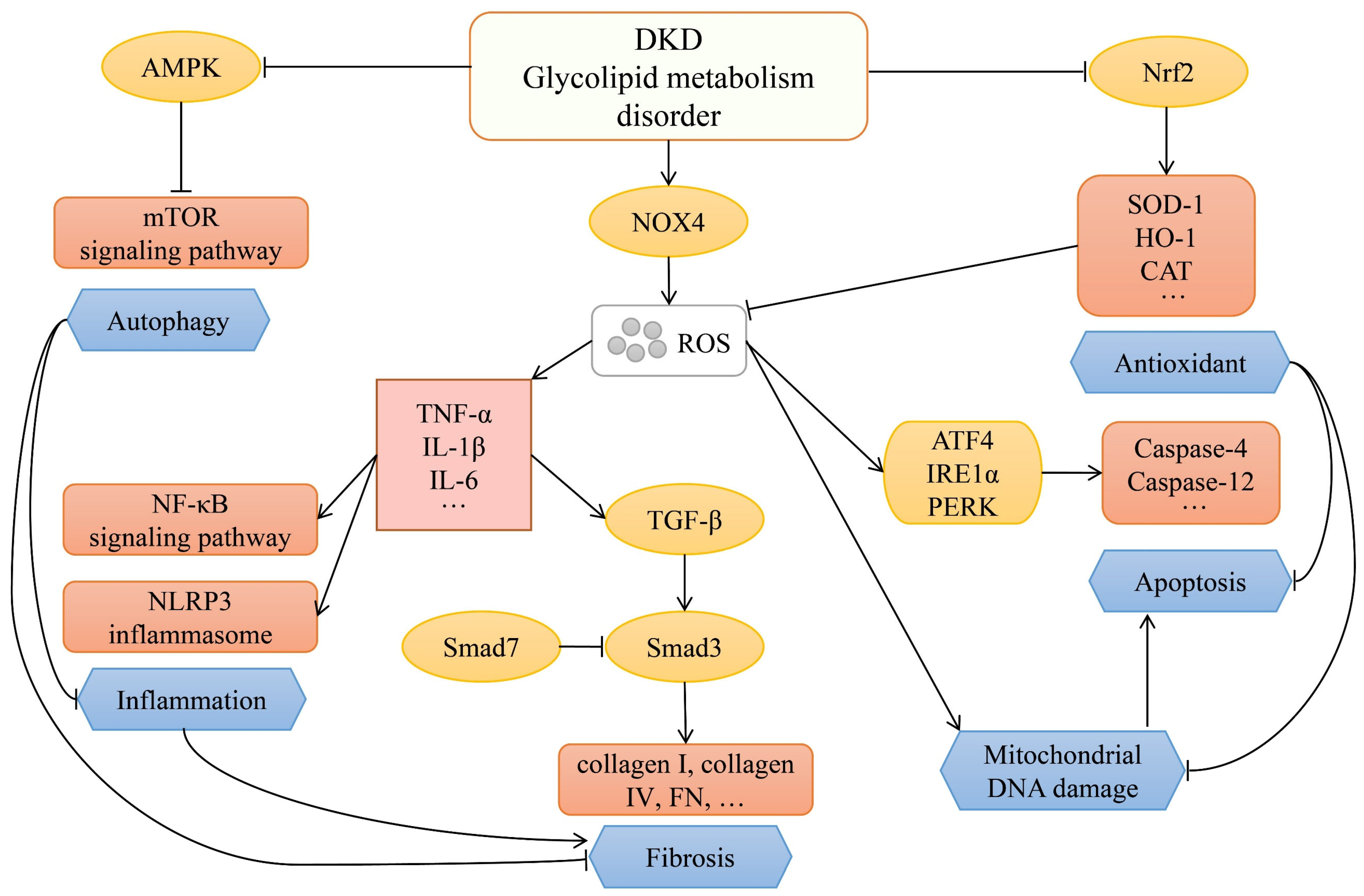
:1. Introduction
2. Signaling Pathways for DKD Progression
3. Natural Compounds
3.1. Phenolics
3.2. Alkaloids
3.3. Flavonoids
3.4. Terpenoids
3.5. Saponins
3.6. Other Compounds
4. Discussion
5. Methodology
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, L.; Han, Q.; Zhou, L.; Bai, L.; Wang, Y.; Wu, Y.; Ren, H.; Zou, Y.; Li, S.; Su, Q.; et al. Addition of glomerular lesion severity improves the value of anemia status for the prediction of renal outcomes in Chinese patients with type 2 diabetes. Ren. Fail. 2022, 44, 346–357. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, L.; Hsiung, J.T.; Arif, Y.; Soohoo, M.; Jackson, N.; Gosmanova, E.O.; Budoff, M.; Kovesdy, C.P.; Kalantar-Zadeh, K.; Streja, E. Serum Low-Density Lipoprotein Cholesterol and Cardiovascular Disease Risk Across Chronic Kidney Disease Stages (Data from 1.9 Million United States Veterans). Am. J. Cardiol. 2022, 170, 47–55. [Google Scholar] [CrossRef]
- Wu, T.H.; Chang, L.H.; Chu, C.H.; Hwu, C.M.; Chen, H.S.; Lin, L.Y. Soluble tumor necrosis factor receptor 2 is associated with progressive diabetic kidney disease in patients with type 2 diabetes mellitus. PLoS ONE 2022, 17, e0266854. [Google Scholar] [CrossRef] [PubMed]
- Chinese Diabetes Society. Guideline for the prevention and treatment of type 2 diabetes mellitus in China (2020 edition). Chin. J. Diabetes Mellit. 2021, 13, 315–409. [Google Scholar]
- Navaneethan, S.D.; Zoungas, S.; Caramori, M.L.; Chan, J.C.N.; Heerspink, H.J.L.; Hurst, C.; Liew, A.; Michos, E.D.; Olowu, W.A.; Sadusky, T.; et al. Diabetes Management in Chronic Kidney Disease: Synopsis of the 2020 KDIGO Clinical Practice Guideline. Ann. Intern. Med. 2021, 174, 385–394. [Google Scholar] [CrossRef]
- Association, A.D. 9. Pharmacologic Approaches to Glycemic Treatment: Standards of Medical Care in Diabetes-2021. Diabetes Care 2021, 44 (Suppl. S1), S111–S124. [Google Scholar] [CrossRef] [PubMed]
- Au, P.C.M.; Tan, K.C.B.; Cheung, B.M.Y.; Wong, I.C.K.; Li, H.L.; Cheung, C.L. Association between SGLT2 Inhibitors vs DPP4 Inhibitors and Renal Outcomes among Patients with Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2022, 107, e2962–e2970. [Google Scholar] [CrossRef] [PubMed]
- de Boer, I.H.; Caramori, M.L.; Chan, J.C.N.; Heerspink, H.J.L.; Hurst, C.; Khunti, K.; Liew, A.; Michos, E.D.; Navaneethan, S.D.; Olowu, W.A.; et al. Executive summary of the 2020 KDIGO Diabetes Management in CKD Guideline: Evidence-based advances in monitoring and treatment. Kidney Int. 2020, 98, 839–848. [Google Scholar] [CrossRef] [PubMed]
- McMurray, J.J.V.; Solomon, S.D.; Inzucchi, S.E.; Køber, L.; Kosiborod, M.N.; Martinez, F.A.; Ponikowski, P.; Sabatine, M.S.; Anand, I.S.; Bělohlávek, J.; et al. Dapagliflozin in Patients with Heart Failure and Reduced Ejection Fraction. N. Engl. J. Med. 2019, 381, 1995–2008. [Google Scholar] [CrossRef]
- Packer, M.; Anker, S.D.; Butler, J.; Filippatos, G.; Pocock, S.J.; Carson, P.; Januzzi, J.; Verma, S.; Tsutsui, H.; Brueckmann, M.; et al. Cardiovascular and Renal Outcomes with Empagliflozin in Heart Failure. N. Engl. J. Med. 2020, 383, 1413–1424. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.T.P.; Tang, S.C.W. Advances in the management of diabetic kidney disease: Beyond sodium-glucose co-transporter 2 inhibitors. Korean J. Nephrol. 2022, 21, 285. [Google Scholar] [CrossRef] [PubMed]
- Thotamgari, S.R.; Grewal, U.S.; Sheth, A.R.; Babbili, A.; Dominic, P. Can glucagon-like peptide-1 receptor agonists and dipeptidyl peptidase-4 inhibitors help in mitigating the risk of atrial fibrillation in patients with diabetes? Cardiovasc. Endocrinol. Metab. 2022, 11, e0265. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Liu, T.; Wang, X.; Sun, J. Research progress of coumarins and their derivatives in the treatment of diabetes. J. Enzym. Inhib. Med. Chem. 2022, 37, 616–628. [Google Scholar] [CrossRef] [PubMed]
- Ajayi, A.M.; Badaki, V.; Adebayo, O.G.; Ben-Azu, B. Plukenetia conophora seed oil ameliorates streptozotocin-induced hyperglycaemia and oxidative stress in rats. Biomark. Biochem. Indic. Expo. Response Susceptibility Chem. 2022, 27, 240–246. [Google Scholar] [CrossRef]
- Molitch, M.E.; Adler, A.I.; Flyvbjerg, A.; Nelson, R.G.; So, W.Y.; Wanner, C.; Kasiske, B.L.; Wheeler, D.C.; de Zeeuw, D.; Mogensen, C.E. Diabetic kidney disease: A clinical update from Kidney Disease: Improving Global Outcomes. Kidney Int. 2015, 87, 20–30. [Google Scholar] [CrossRef]
- Gillard, P.; Schnell, O.; Groop, P.H. The nephrological perspective on SGLT-2 inhibitors in type 1 diabetes. Diabetes Res. Clin. Pract. 2020, 170, 108462. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Wan, C.; Song, A.; Qiu, Y.; Xiong, W.; Zhang, C. Oxidative Stress and Renal Fibrosis: Mechanisms and Therapies. Adv. Exp. Med. Biol. 2019, 1165, 585–604. [Google Scholar] [PubMed]
- Yang, Q.; Wu, F.R.; Wang, J.N.; Gao, L.; Jiang, L.; Li, H.D.; Ma, Q.; Liu, X.Q.; Wei, B.; Zhou, L.; et al. Nox4 in renal diseases: An update. Free Radic. Biol. Med. 2018, 124, 466–472. [Google Scholar] [CrossRef]
- Wan, C.; Su, H.; Zhang, C. Role of NADPH Oxidase in Metabolic Disease-Related Renal Injury: An Update. Oxidative Med. Cell. Longev. 2016, 2016, 7813072. [Google Scholar] [CrossRef] [PubMed]
- Rhee, E.P. NADPH Oxidase 4 at the Nexus of Diabetes, Reactive Oxygen Species, and Renal Metabolism. J. Am. Soc. Nephrol. JASN 2016, 27, 337–339. [Google Scholar] [CrossRef]
- Li, K.; Li, Q. LINC00323 mediates the role of M1 macrophage polarization in diabetic nephropathy through PI3K/AKT signaling pathway. Hum. Immunol. 2021, 82, 960–967. [Google Scholar] [CrossRef] [PubMed]
- Calle, P.; Hotter, G. Macrophage Phenotype and Fibrosis in Diabetic Nephropathy. Int. J. Mol. Sci. 2020, 21, 2806. [Google Scholar] [CrossRef]
- Tang, S.C.W.; Yiu, W.H. Innate immunity in diabetic kidney disease. Nat. Rev. Nephrol. 2020, 16, 206–222. [Google Scholar] [CrossRef] [PubMed]
- Frangogiannis, N.G. Transforming growth factor-β in myocardial disease. Nat. Rev. Cardiol. 2022, 19, 435–455. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.Y.; Liu, X.S.; Huang, X.R.; Yu, X.Q.; Lan, H.Y. Diverse Role of TGF-β in Kidney Disease. Front. Cell Dev. Biol. 2020, 8, 123. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Meng, X.M.; Huang, X.R.; Lan, H.Y. The preventive and therapeutic implication for renal fibrosis by targetting TGF-β/Smad3 signaling. Clin. Sci. 2018, 132, 1403–1415. [Google Scholar] [CrossRef]
- Bhargava, P.; Schnellmann, R.G. Mitochondrial energetics in the kidney. Nat. Rev. Nephrol. 2017, 13, 629–646. [Google Scholar] [CrossRef]
- Victor, P.; Umapathy, D.; George, L.; Juttada, U.; Ganesh, G.V.; Amin, K.N.; Viswanathan, V.; Ramkumar, K.M. Crosstalk between endoplasmic reticulum stress and oxidative stress in the progression of diabetic nephropathy. Cell Stress Chaperones 2021, 26, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, S.; Pergola, P.E.; Zager, R.A.; Vaziri, N.D. Targeting the transcription factor Nrf2 to ameliorate oxidative stress and inflammation in chronic kidney disease. Kidney Int. 2013, 83, 1029–1041. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Isaka, Y.; Yoshimori, T. Autophagy and kidney inflammation. Autophagy 2017, 13, 997–1003. [Google Scholar] [CrossRef]
- Yang, D.; Livingston, M.J.; Liu, Z.; Dong, G.; Zhang, M.; Chen, J.K.; Dong, Z. Autophagy in diabetic kidney disease: Regulation, pathological role and therapeutic potential. Cell. Mol. Life Sci. CMLS 2018, 75, 669–688. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Wu, Y.S.; Li, D.Y.; Tang, J.X.; Liu, H.F. Autophagy and Renal Fibrosis. Aging Dis. 2022, 13, 712–731. [Google Scholar] [CrossRef]
- Tang, C.; Livingston, M.J.; Liu, Z.; Dong, Z. Autophagy in kidney homeostasis and disease. Nat. Rev. Nephrol. 2020, 16, 489–508. [Google Scholar] [CrossRef] [PubMed]
- Neumiller, J.J.; Alicic, R.Z.; Tuttle, K.R. Therapeutic Considerations for Antihyperglycemic Agents in Diabetic Kidney Disease. J. Am. Soc. Nephrol. JASN 2017, 28, 2263–2274. [Google Scholar] [CrossRef]
- Bae, J.H.; Park, E.G.; Kim, S.; Kim, S.G.; Hahn, S.; Kim, N.H. Comparative Renal Effects of Dipeptidyl Peptidase-4 Inhibitors and Sodium-Glucose Cotransporter 2 Inhibitors on Individual Outcomes in Patients with Type 2 Diabetes: A Systematic Review and Network Meta-Analysis. Endocrinol. Metab. 2021, 36, 388–400. [Google Scholar] [CrossRef] [PubMed]
- Hammoud, S.H.; AlZaim, I.; Mougharbil, N.; Koubar, S.; Eid, A.H.; Eid, A.A.; El-Yazbi, A.F. Peri-renal adipose inflammation contributes to renal dysfunction in a non-obese prediabetic rat model: Role of anti-diabetic drugs. Biochem. Pharmacol. 2021, 186, 114491. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.Q.; Tan, H.B.; Zhang, X.Y.; Zhang, Y.Z.; Lin, Q.Y.; Huang, M.Y.; Lin, Z.Y.; Mo, J.Z.; Zhang, Y.; Lan, T.; et al. The Chinese medicine Fufang Zhenzhu Tiaozhi capsule protects against renal injury and inflammation in mice with diabetic kidney disease. J. Ethnopharmacol. 2022, 292, 115165. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Chung, J.O.; Park, S.Y.; Rajamohan, N.; Singh, A.; Kim, J.; Lowe, V.J.; Lee, S. Natural COA water inhibits mitochondrial ROS-mediated apoptosis through Plk3 downregulation under STZ diabetic stress in pancreatic β-cell lines. Biochem. Biophys. Rep. 2022, 30, 101247. [Google Scholar] [CrossRef] [PubMed]
- Fang, D.; Wan, X.; Deng, W.; Guan, H.; Ke, W.; Xiao, H.; Li, Y. Fufang Xue Shuan Tong capsules inhibit renal oxidative stress markers and indices of nephropathy in diabetic rats. Exp. Ther. Med. 2012, 4, 871–876. [Google Scholar] [CrossRef]
- Yao, L.; Liang, X.; Qiao, Y.; Chen, B.; Wang, P.; Liu, Z. Mitochondrial dysfunction in diabetic tubulopathy. Metab. Clin. Exp. 2022, 131, 155195. [Google Scholar] [CrossRef]
- Yuan, T.; Yang, T.; Chen, H.; Fu, D.; Hu, Y.; Wang, J.; Yuan, Q.; Yu, H.; Xu, W.; Xie, X. New insights into oxidative stress and inflammation during diabetes mellitus-accelerated atherosclerosis. Redox Biol. 2019, 20, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Aboolian, A.; Urner, S.; Roden, M.; Jha, J.C.; Jandeleit-Dahm, K. Diabetic Kidney Disease: From Pathogenesis to Novel Treatment Possibilities. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2022; pp. 269–307. [Google Scholar]
- Wu, X.M.; Gao, Y.B.; Xu, L.P.; Zou, D.W.; Zhu, Z.Y.; Wang, X.L.; Yao, W.J.; Luo, L.T.; Tong, Y.; Tian, N.X.; et al. Tongxinluo Inhibits Renal Fibrosis in Diabetic Nephropathy: Involvement of the Suppression of Intercellular Transfer of TGF-β1-Containing Exosomes from GECs to GMCs. Am. J. Chin. Med. 2017, 45, 1075–1092. [Google Scholar] [CrossRef] [PubMed]
- Simón, J.; Casado-Andrés, M.; Goikoetxea-Usandizaga, N.; Serrano-Maciá, M.; Martínez-Chantar, M.L. Nutraceutical Properties of Polyphenols against Liver Diseases. Nutrients 2020, 12, 3517. [Google Scholar] [CrossRef]
- Li, A.N.; Li, S.; Zhang, Y.J.; Xu, X.R.; Chen, Y.M.; Li, H.B. Resources and biological activities of natural polyphenols. Nutrients 2014, 6, 6020–6047. [Google Scholar] [CrossRef] [PubMed]
- Ahamad, J.; Toufeeq, I.; Khan, M.A.; Ameen, M.S.M.; Anwer, E.T.; Uthirapathy, S.; Mir, S.R.; Ahmad, J. Oleuropein: A natural antioxidant molecule in the treatment of metabolic syndrome. Phytother. Res. PTR 2019, 33, 3112–3128. [Google Scholar] [CrossRef]
- Marrano, N.; Spagnuolo, R.; Biondi, G.; Cignarelli, A.; Perrini, S.; Vincenti, L.; Laviola, L.; Giorgino, F.; Natalicchio, A. Effects of Extra Virgin Olive Oil Polyphenols on Beta-Cell Function and Survival. Plants 2021, 10, 286. [Google Scholar] [CrossRef] [PubMed]
- Chaari, A. Inhibition of human islet amyloid polypeptide aggregation and cellular toxicity by oleuropein and derivatives from olive oil. Int. J. Biol. Macromol. 2020, 162, 284–300. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhao, H.; Wang, A. Oleuropein alleviates gestational diabetes mellitus by activating AMPK signaling. Endocr. Connect. 2021, 10, 45–53. [Google Scholar] [CrossRef]
- Liu, Y.; Dai, W.; Ye, S. The olive constituent oleuropein exerts nephritic protective effects on diabetic nephropathy in db/db mice. Arch. Physiol. Biochem. 2022, 128, 455–462. [Google Scholar] [CrossRef]
- Acar-Tek, N.; Ağagündüz, D. Olive Leaf (Olea europaea L. folium): Potential Effects on Glycemia and Lipidemia. Ann. Nutr. Metab. 2020, 76, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Michno, A.; Grużewska, K.; Ronowska, A.; Gul-Hinc, S.; Zyśk, M.; Jankowska-Kulawy, A. Resveratrol Inhibits Metabolism and Affects Blood Platelet Function in Type 2 Diabetes. Nutrients 2022, 14, 1633. [Google Scholar] [CrossRef] [PubMed]
- Szkudelski, T.; Szkudelska, K. Resveratrol and diabetes: From animal to human studies. Biochim. Biophys. Acta 2015, 1852, 1145–1154. [Google Scholar] [CrossRef]
- Choudhury, H.; Pandey, M.; Hua, C.K.; Mun, C.S.; Jing, J.K.; Kong, L.; Ern, L.Y.; Ashraf, N.A.; Kit, S.W.; Yee, T.S.; et al. An update on natural compounds in the remedy of diabetes mellitus: A systematic review. J. Tradit. Complement. Med. 2018, 8, 361–376. [Google Scholar] [CrossRef] [PubMed]
- Mahjabeen, W.; Khan, D.A.; Mirza, S.A. Role of resveratrol supplementation in regulation of glucose hemostasis, inflammation and oxidative stress in patients with diabetes mellitus type 2: A randomized, placebo-controlled trial. Complement. Ther. Med. 2022, 66, 102819. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.; Zhang, Y. Effects of resveratrol therapy on glucose metabolism, insulin resistance, inflammation, and renal function in the elderly patients with type 2 diabetes mellitus: A randomized controlled clinical trial protocol. Medicine 2022, 101, e30049. [Google Scholar] [CrossRef]
- Zhang, J.; Dong, X.J.; Ding, M.R.; You, C.Y.; Lin, X.; Wang, Y.; Wu, M.J.; Xu, G.F.; Wang, G.D. Resveratrol decreases high glucose-induced apoptosis in renal tubular cells via suppressing endoplasmic reticulum stress. Mol. Med. Rep. 2020, 22, 4367–4375. [Google Scholar] [CrossRef]
- Wang, F.; Li, R.; Zhao, L.; Ma, S.; Qin, G. Resveratrol ameliorates renal damage by inhibiting oxidative stress-mediated apoptosis of podocytes in diabetic nephropathy. Eur. J. Pharm. 2020, 885, 173387. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Chi, Y.; Kang, Y.; Lu, H.; Niu, H.; Liu, W.; Li, Y. Resveratrol ameliorates podocyte damage in diabetic mice via SIRT1/PGC-1α mediated attenuation of mitochondrial oxidative stress. J. Cell. Physiol. 2019, 234, 5033–5043. [Google Scholar] [CrossRef]
- Du, L.; Wang, L.; Wang, B.; Wang, J.; Hao, M.; Chen, Y.B.; Li, X.Z.; Li, Y.; Jiang, Y.F.; Li, C.C.; et al. A novel compound AB38b attenuates oxidative stress and ECM protein accumulation in kidneys of diabetic mice through modulation of Keap1/Nrf2 signaling. Acta Pharmacol. Sin. 2020, 41, 358–372. [Google Scholar] [CrossRef]
- Qiao, Y.; Gao, K.; Wang, Y.; Wang, X.; Cui, B. Resveratrol ameliorates diabetic nephropathy in rats through negative regulation of the p38 MAPK/TGF-β1 pathway. Exp. Ther. Med. 2017, 13, 3223–3230. [Google Scholar] [CrossRef]
- Gu, W.; Wang, X.; Zhao, H.; Geng, J.; Li, X.; Zheng, K.; Guan, Y.; Hou, X.; Wang, C.; Song, G. Resveratrol ameliorates diabetic kidney injury by reducing lipotoxicity and modulates expression of components of the junctional adhesion molecule-like/sirtuin 1 lipid metabolism pathway. Eur. J. Pharm. 2022, 918, 174776. [Google Scholar] [CrossRef]
- Zhu, H.; Zhong, S.; Yan, H.; Wang, K.; Chen, L.; Zhou, M.; Li, Y. Resveratrol reverts Streptozotocin-induced diabetic nephropathy. Front. Biosci. 2020, 25, 699–709. [Google Scholar]
- Salehi, B.; Mishra, A.P.; Nigam, M.; Sener, B.; Kilic, M.; Sharifi-Rad, M.; Fokou, P.V.T.; Martins, N.; Sharifi-Rad, J. Resveratrol: A Double-Edged Sword in Health Benefits. Biomedicines 2018, 6, 91. [Google Scholar] [CrossRef]
- Shaito, A.; Posadino, A.M.; Younes, N.; Hasan, H.; Halabi, S.; Alhababi, D.; Al-Mohannadi, A.; Abdel-Rahman, W.M.; Eid, A.H.; Nasrallah, G.K.; et al. Potential Adverse Effects of Resveratrol: A Literature Review. Int. J. Mol. Sci. 2020, 21, 2084. [Google Scholar] [CrossRef]
- Dong, Z.; Bian, L.; Wang, Y.L.; Sun, L.M. Gastrodin protects against high glucose-induced cardiomyocyte toxicity via GSK-3β-mediated nuclear translocation of Nrf2. Hum. Exp. Toxicol. 2021, 40, 1584–1597. [Google Scholar] [CrossRef]
- Bai, Y.; Mo, K.; Wang, G.; Chen, W.; Zhang, W.; Guo, Y.; Sun, Z. Intervention of Gastrodin in Type 2 Diabetes Mellitus and Its Mechanism. Front. Pharmacol. 2021, 12, 710722. [Google Scholar] [CrossRef]
- Deng, C.-K.; Mu, Z.-H.; Miao, Y.-H.; Liu, Y.-D.; Zhou, L.; Huang, Y.-J.; Zhang, F.; Wang, Y.-Y.; Yang, Z.-H.; Qian, Z.-Y.; et al. Gastrodin Ameliorates Motor Learning Deficits Through Preserving Cerebellar Long-Term Depression Pathways in Diabetic Rats. Front. Neurosci. 2019, 13, 1239. [Google Scholar] [CrossRef]
- Xu, G.; Huang, K.; Zhou, J. Hepatic AMP Kinase as a Potential Target for Treating Nonalcoholic Fatty Liver Disease: Evidence from Studies of Natural Products. Curr. Med. Chem. 2018, 25, 889–907. [Google Scholar] [CrossRef] [PubMed]
- Ye, T.; Meng, X.; Zhai, Y.; Xie, W.; Wang, R.; Sun, G.; Sun, X. Gastrodin Ameliorates Cognitive Dysfunction in Diabetes Rat Model via the Suppression of Endoplasmic Reticulum Stress and NLRP3 Inflammasome Activation. Front. Pharmacol. 2018, 9, 1346. [Google Scholar] [CrossRef]
- Huang, L.; Shao, M.; Zhu, Y. Gastrodin inhibits high glucose-induced inflammation, oxidative stress and apoptosis in podocytes by activating the AMPK/Nrf2 signaling pathway. Exp. Ther. Med. 2022, 23, 168. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, J.; Peng, M.; Meng, H.; Ma, H.; Cai, P.; Xu, Y.; Zhao, Q.; Si, G. A Review on Central Nervous System Effects of Gastrodin. Front. Pharmacol. 2018, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Wang, R.; Li, W.; Zhu, H.; Fei, S.; Shi, H.; Lu, N.; Ung, C.O.L.; Hu, H.; Han, S. Clinical and economic analysis of Gastrodin injection for dizziness or vertigo: A retrospective cohort study based on electronic health records in China. Chin. Med. 2022, 17, 6. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, Y.; Zhu, L.; Han, C. Clinical Research Progress on Treatment of Diabetic Peripheral Neuropathy by Acupoint Injection of Traditional Chinese Medicine. Clin. J. Tradit. Chin. Med. 2018, 30, 1574–1577. [Google Scholar]
- Ponikvar-Svet, M.; Zeiger, D.N.; Liebman, J.F. Alkaloids and Selected Topics in Their Thermochemistry. Molecules 2021, 26, 6715. [Google Scholar] [CrossRef]
- Abookleesh, F.L.; Al-Anzi, B.S.; Ullah, A. Potential Antiviral Action of Alkaloids. Molecules 2022, 27, 903. [Google Scholar] [CrossRef] [PubMed]
- Mohamadi, N.; Sharififar, F.; Pournamdari, M.; Ansari, M. A Review on Biosynthesis, Analytical Techniques, and Pharmacological Activities of Trigonelline as a Plant Alkaloid. J. Diet. Suppl. 2018, 15, 207–222. [Google Scholar] [CrossRef]
- Shao, X.; Chen, C.; Miao, C.; Yu, X.; Li, X.; Geng, J.; Fan, D.; Lin, X.; Chen, Z.; Shi, Y. Expression analysis of microRNAs and their target genes during experimental diabetic renal lesions in rats administered with ginsenoside Rb1 and trigonelline. Pharmazie 2019, 74, 492–498. [Google Scholar] [PubMed]
- Chen, C.; Shi, Y.; Ma, J.; Chen, Z.; Zhang, M.; Zhao, Y. Trigonelline reverses high glucose-induced proliferation, fibrosis of mesangial cells via modulation of Wnt signaling pathway. Diabetol. Metab. Syndr. 2022, 14, 28. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, Q.; Wang, C.; Lou, Z.; Li, Q. Trigonelline reduced diabetic nephropathy and insulin resistance in type 2 diabetic rats through peroxisome proliferator-activated receptor-γ. Exp. Ther. Med. 2019, 18, 1331–1337. [Google Scholar] [CrossRef]
- Chen, C.; Ma, J.; Miao, C.S.; Zhang, H.; Zhang, M.; Cao, X.; Shi, Y. Trigonelline induces autophagy to protect mesangial cells in response to high glucose via activating the miR-5189-5p-AMPK pathway. Phytomed. Int. J. Phytother. Phytopharm. 2021, 92, 153614. [Google Scholar] [CrossRef]
- Zhou, J.; Chan, L.; Zhou, S. Trigonelline: A plant alkaloid with therapeutic potential for diabetes and central nervous system disease. Curr. Med. Chem. 2012, 19, 3523–3531. [Google Scholar] [CrossRef]
- Jia, X.; Shao, W.; Tian, S. Berberine alleviates myocardial ischemia-reperfusion injury by inhibiting inflammatory response and oxidative stress: The key function of miR-26b-5p-mediated PTGS2/MAPK signal transduction. Pharm. Biol. 2022, 60, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Akdad, M.; Ameziane, R.; Khallouki, F.; Bakri, Y.; Eddouks, M. Antidiabetic Phytocompounds Acting as Glucose Transport Stimulators. Endocr. Metab. Immune Disord. Drug Targets 2022. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Ma, L.; Wang, G.; Yang, J.; Zhang, M.; Wang, X.; Su, J.; Xie, M. In vitro Antimicrobial Activity and the Mechanism of Berberine against Methicillin-Resistant Staphylococcus aureus Isolated from Bloodstream Infection Patients. Infect. Drug Resist. 2022, 15, 1933–1944. [Google Scholar] [CrossRef] [PubMed]
- Dian, L.; Xu, Z.; Sun, Y.; Li, J.; Lu, H.; Zheng, M.; Wang, J.; Drobot, L.; Horak, I. Berberine alkaloids inhibit the proliferation and metastasis of breast carcinoma cells involving Wnt/β-catenin signaling and EMT. Phytochemistry 2022, 200, 113217. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Chen, H.; Zhang, X.; Lou, W.; Zhang, P.; Qiu, Y.; Zhang, C.; Wang, Y.; Liu, W.J. The Effect of Berberine on Metabolic Profiles in Type 2 Diabetic Patients: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Oxidative Med. Cell. Longev. 2021, 2021, 2074610. [Google Scholar] [CrossRef]
- Ni, W.J.; Zhou, H.; Ding, H.H.; Tang, L.Q. Berberine ameliorates renal impairment and inhibits podocyte dysfunction by targeting the phosphatidylinositol 3-kinase-protein kinase B pathway in diabetic rats. J. Diabetes Investig. 2020, 11, 297–306. [Google Scholar] [CrossRef]
- Ni, W.J.; Guan, X.M.; Zeng, J.; Zhou, H.; Meng, X.M.; Tang, L.Q. Berberine regulates mesangial cell proliferation and cell cycle to attenuate diabetic nephropathy through the PI3K/Akt/AS160/GLUT1 signalling pathway. J. Cell. Mol. Med. 2022, 26, 1144–1155. [Google Scholar] [CrossRef]
- Zhang, X.; Guan, T.; Yang, B.; Chi, Z.; Wan, Q.; Gu, H.F. Protective effect of berberine on high glucose and hypoxia-induced apoptosis via the modulation of HIF-1α in renal tubular epithelial cells. Am. J. Transl. Res. 2019, 11, 669–682. [Google Scholar]
- Sun, J.; Chen, X.; Liu, T.; Jiang, X.; Wu, Y.; Yang, S.; Hua, W.; Li, Z.; Huang, H.; Ruan, X.; et al. Berberine Protects against Palmitate-Induced Apoptosis in Tubular Epithelial Cells by Promoting Fatty Acid Oxidation. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 1484–1492. [Google Scholar] [CrossRef]
- Rong, Q.; Han, B.; Li, Y.; Yin, H.; Li, J.; Hou, Y. Berberine Reduces Lipid Accumulation by Promoting Fatty Acid Oxidation in Renal Tubular Epithelial Cells of the Diabetic Kidney. Front. Pharmacol. 2022, 12, 729384. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Liu, L.; Gan, L.; Hu, Y.; Xiang, P.; Xing, Y.; Zhu, J.; Ye, S. Berberine Acts on C/EBPβ/lncRNA Gas5/miR-18a-5p Loop to Decrease the Mitochondrial ROS Generation in HK-2 Cells. Front. Endocrinol. 2021, 12, 675834. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Jiang, M.; Zhao, Y.; Gong, J.; Su, H.; Yuan, F.; Fang, K.; Yuan, X.; Yu, X.; Dong, H.; et al. Berberine protects against diabetic kidney disease via promoting PGC-1α-regulated mitochondrial energy homeostasis. Br. J. Pharmacol. 2020, 177, 3646–3661. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Zhao, Y.; Gong, J.; Huang, W.; Su, H.; Yuan, F.; Fang, K.; Wang, D.; Li, J.; Zou, X.; et al. Berberine Protects Glomerular Podocytes via Inhibiting Drp1-Mediated Mitochondrial Fission and Dysfunction. Theranostics 2019, 9, 1698–1713. [Google Scholar] [CrossRef]
- Yang, G.; Zhao, Z.; Zhang, X.; Wu, A.; Huang, Y.; Miao, Y.; Yang, M. Effect of berberine on the renal tubular epithelial-to-mesenchymal transition by inhibition of the Notch/snail pathway in diabetic nephropathy model KKAy mice. Drug Des. Dev. Ther. 2017, 11, 1065–1079. [Google Scholar] [CrossRef]
- Zhu, L.; Han, J.; Yuan, R.; Xue, L.; Pang, W. Berberine ameliorates diabetic nephropathy by inhibiting TLR4/NF-κB pathway. Biol. Res. 2018, 51, 9. [Google Scholar] [CrossRef]
- Li, C.; Guan, X.M.; Wang, R.Y.; Xie, Y.S.; Zhou, H.; Ni, W.J.; Tang, L.Q. Berberine mitigates high glucose-induced podocyte apoptosis by modulating autophagy via the mTOR/P70S6K/4EBP1 pathway. Life Sci. 2020, 243, 117277. [Google Scholar] [CrossRef]
- Mohammadzadeh, N.; Mehri, S.; Hosseinzadeh, H. Berberis vulgaris and its constituent berberine as antidotes and protective agents against natural or chemical toxicities. Iran. J. Basic Med. Sci. 2017, 20, 538–551. [Google Scholar]
- Imenshahidi, M.; Hosseinzadeh, H. Berberine and barberry (Berberis vulgaris): A clinical review. Phytother. Res. PTR 2019, 33, 504–523. [Google Scholar] [CrossRef]
- Spinozzi, S.; Colliva, C.; Camborata, C.; Roberti, M.; Ianni, C.; Neri, F.; Calvarese, C.; Lisotti, A.; Mazzella, G.; Roda, A. Berberine and Its Metabolites: Relationship between Physicochemical Properties and Plasma Levels after Administration to Human Subjects. J. Nat. Prod. 2014, 77, 766–772. [Google Scholar] [CrossRef]
- Liu, Y.-T.; Hao, H.-P.; Xie, H.-G.; Lai, L.; Wang, Q.; Liu, C.-X.; Wang, G.-J. Extensive Intestinal First-Pass Elimination and Predominant Hepatic Distribution of Berberine Explain Its Low Plasma Levels in Rats. Drug Metab. Dispos. 2010, 38, 1779–1784. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Sun, B.; Hou, Y.; Liu, L.; Sun, J.; Xu, F.; Li, D.; Hua, H. Anti-breast cancer sinomenine derivatives via mechanisms of apoptosis induction and metastasis reduction. J. Enzym. Inhib. Med. Chem. 2022, 37, 1870–1883. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xie, H.; Zhang, H. Protective effect of sinomenine against inflammation and oxidative stress in gestational diabetes mellitus in female rats via TLR4/MyD88/NF-κB signaling pathway. J. Food Biochem. 2021, 45, e13952. [Google Scholar] [CrossRef] [PubMed]
- Yin, Q.; Xia, Y.; Wang, G. Sinomenine alleviates high glucose-induced renal glomerular endothelial hyperpermeability by inhibiting the activation of RhoA/ROCK signaling pathway. Biochem. Biophys. Res. Commun. 2016, 477, 881–886. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, J. Sinomenine alleviates glomerular endothelial permeability by activating the C/EBP-α/claudin-5 signaling pathway. Hum. Cell 2022, 35, 1453–1463. [Google Scholar] [CrossRef]
- Zhu, M.; Wang, H.; Chen, J.; Zhu, H. Sinomenine improve diabetic nephropathy by inhibiting fibrosis and regulating the JAK2/STAT3/SOCS1 pathway in streptozotocin-induced diabetic rats. Life Sci. 2021, 265, 118855. [Google Scholar] [CrossRef]
- Huang, Z.; Mao, X.; Chen, J.; He, J.; Shi, S.; Gui, M.; Gao, H.; Hong, Z. The Efficacy and Safety of Zhengqing Fengtongning for Knee Osteoarthritis: A Systematic Review and Meta-Analysis of Randomized Clinical Trials. Evid.-Based Complement. Altern. Med. 2022, 2022, 2768444. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Mao, X.; Chen, J.; He, J.; Shi, S.; Gui, M.; Gao, H.; Hong, Z. Sinomenine hydrochloride injection for knee osteoarthritis: A protocol for systematic review and meta-analysis. Medicine 2022, 101, e28503. [Google Scholar] [CrossRef]
- Zhang, Y.S.; Han, J.Y.; Iqbal, O.; Liang, A.H. Research Advances and Prospects on Mechanism of Sinomenin on Histamine Release and the Binding to Histamine Receptors. Int. J. Mol. Sci. 2018, 20, 70. [Google Scholar] [CrossRef]
- Huang, H.; Zhang, E.B.; Yi, O.Y.; Wu, H.; Deng, G.; Huang, Y.M.; Liu, W.L.; Yan, J.Y.; Cai, X. Sex-related differences in safety profiles, pharmacokinetics and tissue distribution of sinomenine hydrochloride in rats. Arch. Toxicol. 2022. [Google Scholar] [CrossRef]
- Middleton, E., Jr.; Kandaswami, C.; Theoharides, T.C. The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacol. Rev. 2000, 52, 673–751. [Google Scholar] [PubMed]
- Dias, M.C.; Pinto, D.; Silva, A.M.S. Plant Flavonoids: Chemical Characteristics and Biological Activity. Molecules 2021, 26, 5377. [Google Scholar] [CrossRef] [PubMed]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić Bok, V.; Salopek-Sondi, B. The Role of Polyphenols in Abiotic Stress Response: The Influence of Molecular Structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Sharma, A.; Monga, V.; Bhatia, R. Compendium of naringenin: Potential sources, analytical aspects, chemistry, nutraceutical potentials and pharmacological profile. Crit. Rev. Food Sci. Nutr. 2022, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Nyane, N.A.; Tlaila, T.B.; Malefane, T.G.; Ndwandwe, D.E.; Owira, P.M.O. Metformin-like antidiabetic, cardio-protective and non-glycemic effects of naringenin: Molecular and pharmacological insights. Eur. J. Pharmacol. 2017, 803, 103–111. [Google Scholar] [CrossRef]
- Khan, M.F.; Mathur, A.; Pandey, V.K.; Kakkar, P. Naringenin alleviates hyperglycemia-induced renal toxicity by regulating activating transcription factor 4-C/EBP homologous protein mediated apoptosis. J. Cell Commun. Signal. 2022, 16, 271–291. [Google Scholar] [CrossRef]
- Ding, S.; Qiu, H.; Huang, J.; Chen, R.; Zhang, J.; Huang, B.; Zou, X.; Cheng, O.; Jiang, Q. Activation of 20-HETE/PPARs involved in reno-therapeutic effect of naringenin on diabetic nephropathy. Chem. Biol. Interact. 2019, 307, 116–124. [Google Scholar] [CrossRef]
- Yan, N.; Wen, L.; Peng, R.; Li, H.; Liu, H.; Peng, H.; Sun, Y.; Wu, T.; Chen, L.; Duan, Q.; et al. Naringenin Ameliorated Kidney Injury through Let-7a/TGFBR1 Signaling in Diabetic Nephropathy. J. Diabetes Res. 2016, 2016, 8738760. [Google Scholar] [CrossRef]
- Rebello, C.J.; Beyl, R.A.; Lertora, J.J.L.; Greenway, F.L.; Ravussin, E.; Ribnicky, D.M.; Poulev, A.; Kennedy, B.J.; Castro, H.F.; Campagna, S.R.; et al. Safety and pharmacokinetics of naringenin: A randomized, controlled, single-ascending-dose clinical trial. Diabetes Obes. Metab. 2020, 22, 91–98. [Google Scholar] [CrossRef]
- Ranawat, P.; Bakshi, N. Naringenin; a bioflavonoid, impairs the reproductive potential of male mice. Toxicol. Mech. Methods 2017, 27, 417–427. [Google Scholar] [CrossRef]
- Yan, L.; Vaghari-Tabari, M.; Malakoti, F.; Moein, S.; Qujeq, D.; Yousefi, B.; Asemi, Z. Quercetin: An effective polyphenol in alleviating diabetes and diabetic complications. Crit. Rev. Food Sci. Nutr. 2022, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Bu, F.; Huang, L.; Xu, W.; Wang, W.; Wu, Q. Preclinical evidence of the effect of quercetin on diabetic nephropathy: A meta-analysis of animal studies. Eur. J. Pharm. 2022, 921, 174868. [Google Scholar] [CrossRef]
- Adeshara, K.A.; Bangar, N.; Diwan, A.G.; Tupe, R.S. Plasma glycation adducts and various RAGE isoforms are intricately associated with oxidative stress and inflammatory markers in type 2 diabetes patients with vascular complications. Diabetes Metab. Syndr. 2022, 16, 102441. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Li, K.; Zhang, Y.; Li, H.; Li, A.; Xu, Y.; Wei, B. Quercetin liposomes ameliorate streptozotocin-induced diabetic nephropathy in diabetic rats. Sci. Rep. 2020, 10, 2440. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Y.; Xu, L.; Shi, J.; Yu, X.; Wang, X.; Li, X.; Jiang, H.; Yang, T.; Yin, X.; et al. Quercetin Attenuates Podocyte Apoptosis of Diabetic Nephropathy Through Targeting EGFR Signaling. Front. Pharmacol. 2021, 12, 792777. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Wang, Y.; Pan, Q.; Chen, X.; Chen, S.; Li, X.; Yao, W. Quercetin attenuates the proliferation, inflammation, and oxidative stress of high glucose-induced human mesangial cells by regulating the miR-485-5p/YAP1 pathway. Int. J. Immunopathol. Pharm. 2022, 36, 20587384211066440. [Google Scholar] [CrossRef]
- Lei, D.; Chengcheng, L.; Xuan, Q.; Yibing, C.; Lei, W.; Hao, Y.; Xizhi, L.; Yuan, L.; Xiaoxing, Y.; Qian, L. Quercetin inhibited mesangial cell proliferation of early diabetic nephropathy through the Hippo pathway. Pharm. Res. 2019, 146, 104320. [Google Scholar] [CrossRef]
- Yuan, Y.; Sun, H.; Sun, Z. Advanced glycation end products (AGEs) increase renal lipid accumulation: A pathogenic factor of diabetic nephropathy (DN). Lipids Health Dis. 2017, 16, 126. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Yu, J.; Wang, X.; Ge, J.; Li, N. Quercetin improves lipid metabolism via SCAP-SREBP2-LDLr signaling pathway in early stage diabetic nephropathy. Diabetes Metab. Syndr. Obes. Targets Ther. 2019, 12, 827–839. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Pan, Y.; Zhang, Q.Y.; Wang, F.M.; Kong, L.D. Quercetin and allopurinol ameliorate kidney injury in STZ-treated rats with regulation of renal NLRP3 inflammasome activation and lipid accumulation. PLoS ONE 2012, 7, e38285. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.; Peng, H.; Wu, X.; Xu, X.; Kuang, T.; Zhang, J.; Du, L.; Fan, G. The Therapeutic Effects and Mechanisms of Quercetin on Metabolic Diseases: Pharmacological Data and Clinical Evidence. Oxidative Med. Cell. Longev. 2021, 2021, 6678662. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.-Y.; Liao, L.-X.; Tu, P.-F.; Li, W.-W.; Zeng, K.-W. Icariin Inhibits AGE-Induced Injury in PC12 Cells by Directly Targeting Apoptosis Regulator Bax. Oxidative Med. Cell. Longev. 2019, 2019, 7940808. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Zhao, H.; Qiao, C. Icariin protects podocytes from NLRP3 activation by Sesn2-induced mitophagy through the Keap1-Nrf2/HO-1 axis in diabetic nephropathy. Phytomed. Int. J. Phytother. Phytopharm. 2022, 99, 154005. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.Y.; He, Y.H.; Cheng, Y.; Fang, Q.; Ma, R.Y.; Zhou, S.J.; Hao, J.Q. Icariin ameliorates streptozocin-induced diabetic nephropathy through suppressing the TLR4/NF-κB signal pathway. Food Funct. 2021, 12, 1241–1251. [Google Scholar] [CrossRef]
- Li, Y.C.; Ding, X.S.; Li, H.M.; Zhang, C. Icariin attenuates high glucose-induced type ΙV collagen and fibronectin accumulation in glomerular mesangial cells by inhibiting transforming growth factor-β production and signalling through G protein-coupled oestrogen receptor 1. Clin. Exp. Pharmacol. Physiol. 2013, 40, 635–643. [Google Scholar] [CrossRef]
- Jia, Z.; Wang, K.; Zhang, Y.; Duan, Y.; Xiao, K.; Liu, S.; Ding, X. Icariin Ameliorates Diabetic Renal Tubulointerstitial Fibrosis by Restoring Autophagy via Regulation of the miR-192-5p/GLP-1R Pathway. Front. Pharmacol. 2021, 12, 720387. [Google Scholar] [CrossRef] [PubMed]
- Zang, L.; Gao, F.; Huang, A.; Zhang, Y.; Luo, Y.; Chen, L.; Mao, N. Icariin inhibits epithelial mesenchymal transition of renal tubular epithelial cells via regulating the miR-122-5p/FOXP2 axis in diabetic nephropathy rats. J. Pharmacol. Sci. 2022, 148, 204–213. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Wang, Z.; Shi, J. Chapter Seven—Pharmacological effects of icariin. In Advances in Pharmacology; Du, G., Ed.; Academic Press: Cambridge, MA, USA, 2020; Volume 87, pp. 179–203. [Google Scholar]
- Zhou, F.M.; Huang, J.J.; Hu, X.J.; Wang, J.; Zhu, B.Q.; Ding, Z.S.; Huang, S.; Fang, J.J. Protective effects of flavonoids from the leaves of Carya cathayensis Sarg. against H2O2-induced oxidative damage and apoptosis in vitro. Exp. Ther. Med. 2021, 22, 1443. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, N.; Jin, Y.; Xu, H. Cardamonin Promotes the Apoptosis and Chemotherapy Sensitivity to Gemcitabine of Pancreatic Cancer Through Modulating the FOXO3a-FOXM1 Axis. Dose-Response Publ. Int. Hormesis Soc. 2021, 19, 15593258211042163. [Google Scholar] [CrossRef] [PubMed]
- Satsu, H.; Shibata, R.; Suzuki, H.; Kimura, S.; Shimizu, M. Inhibitory Effect of Tangeretin and Cardamonin on Human Intestinal SGLT1 Activity In Vitro and Blood Glucose Levels in Mice In Vivo. Nutrients 2021, 13, 3382. [Google Scholar] [CrossRef]
- Zhang, T.; Yamamoto, N.; Ashida, H. Chalcones suppress fatty acid-induced lipid accumulation through a LKB1/AMPK signaling pathway in HepG2 cells. Food Funct. 2014, 5, 1134–1141. [Google Scholar] [CrossRef] [PubMed]
- Schalkwijk, C.G.; Stehouwer, C.D.A. Methylglyoxal, a Highly Reactive Dicarbonyl Compound, in Diabetes, Its Vascular Complications, and Other Age-Related Diseases. Physiol. Rev. 2020, 100, 407–461. [Google Scholar] [CrossRef]
- Cha, S.H.; Hwang, Y.; Heo, S.J.; Jun, H.S. Diphlorethohydroxycarmalol Attenuates Methylglyoxal-Induced Oxidative Stress and Advanced Glycation End Product Formation in Human Kidney Cells. Oxidative Med. Cell. Longev. 2018, 2018, 3654095. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Fei, X.; Wang, M.; Chen, Q.; Zhao, N. Cardamomin protects from diabetes-induced kidney damage through modulating PI3K/AKT and JAK/STAT signaling pathways in rats. Int. Immunopharmacol. 2022, 107, 108610. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, N.; Asle-Rousta, M.; Rahnema, M.; Amini, R. Morin attenuates memory deficits in a rat model of Alzheimer’s disease by ameliorating oxidative stress and neuroinflammation. Eur. J. Pharm. 2021, 910, 174506. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Zhang, C.; Yang, L.; Zeng, X.; Hu, Y.; Xue, X.; Dai, Y.; Wei, Z. The activation of PPARγ enhances Treg responses through up-regulating CD36/CPT1-mediated fatty acid oxidation and subsequent N-glycan branching of TβRII/IL-2Rα. Cell Commun. Signal. CCS 2022, 20, 48. [Google Scholar] [CrossRef]
- Issac, P.K.; Velayutham, M.; Guru, A.; Sudhakaran, G.; Pachaiappan, R.; Arockiaraj, J. Protective effect of morin by targeting mitochondrial reactive oxygen species induced by hydrogen peroxide demonstrated at a molecular level in MDCK epithelial cells. Mol. Biol. Rep. 2022, 49, 4269–4279. [Google Scholar] [CrossRef] [PubMed]
- Mathur, A.; Pandey, V.K.; Khan, M.F.; Kakkar, P. PHLPP1/Nrf2-Mdm2 axis induces renal apoptosis via influencing nucleo-cytoplasmic shuttling of FoxO1 during diabetic nephropathy. Mol. Cell. Biochem. 2021, 476, 3681–3699. [Google Scholar] [CrossRef]
- Ke, Y.Q.; Liu, C.; Hao, J.B.; Lu, L.; Lu, N.N.; Wu, Z.K.; Zhu, S.S.; Chen, X.L. Morin inhibits cell proliferation and fibronectin accumulation in rat glomerular mesangial cells cultured under high glucose condition. Biomed. Pharmacother. 2016, 84, 622–627. [Google Scholar] [CrossRef]
- Cho, Y.M.; Onodera, H.; Ueda, M.; Imai, T.; Hirose, M. A 13-week subchronic toxicity study of dietary administered morin in F344 rats. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2006, 44, 891–897. [Google Scholar] [CrossRef]
- Caselli, A.; Cirri, P.; Santi, A.; Paoli, P. Morin: A Promising Natural Drug. Curr. Med. Chem. 2016, 23, 774–791. [Google Scholar] [CrossRef]
- Mottaghi, S.; Abbaszadeh, H. The anticarcinogenic and anticancer effects of the dietary flavonoid, morin: Current status, challenges, and future perspectives. Phytother. Res. PTR 2021, 35, 6843–6861. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Gu, H.; Gwon, M.H.; Yun, J.M. Hesperetin suppresses LPS/high glucose-induced inflammatory responses via TLR/MyD88/NF-κB signaling pathways in THP-1 cells. Nutr. Res. Pract. 2021, 15, 591–603. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, N.; Xue, M.; Weickert, M.O.; Thornalley, P.J. Reversal of Insulin Resistance in Overweight and Obese Subjects by trans-Resveratrol and Hesperetin Combination-Link to Dysglycemia, Blood Pressure, Dyslipidemia, and Low-Grade Inflammation. Nutrients 2021, 13, 2374. [Google Scholar] [CrossRef]
- Li, J.; Wang, T.; Liu, P.; Yang, F.; Wang, X.; Zheng, W.; Sun, W. Hesperetin ameliorates hepatic oxidative stress and inflammation via the PI3K/AKT-Nrf2-ARE pathway in oleic acid-induced HepG2 cells and a rat model of high-fat diet-induced NAFLD. Food Funct. 2021, 12, 3898–3918. [Google Scholar] [CrossRef]
- Gong, Y.; Qin, X.Y.; Zhai, Y.Y.; Hao, H.; Lee, J.; Park, Y.D. Inhibitory effect of hesperetin on α-glucosidase: Molecular dynamics simulation integrating inhibition kinetics. Int. J. Biol. Macromol. 2017, 101, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Rasouli, H.; Hosseini-Ghazvini, S.M.; Adibi, H.; Khodarahmi, R. Differential α-amylase/α-glucosidase inhibitory activities of plant-derived phenolic compounds: A virtual screening perspective for the treatment of obesity and diabetes. Food Funct. 2017, 8, 1942–1954. [Google Scholar] [CrossRef] [PubMed]
- Abdou, H.M.; Abd Elkader, H.A.E. The potential therapeutic effects of Trifolium alexandrinum extract, hesperetin and quercetin against diabetic nephropathy via attenuation of oxidative stress, inflammation, GSK-3β and apoptosis in male rats. Chem. Biol. Interact. 2022, 352, 109781. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Kong, L.; Tang, Z.Z.; Zhang, Y.M.; Liu, Y.; Wang, T.Y.; Liu, Y.W. Hesperetin ameliorates diabetic nephropathy in rats by activating Nrf2/ARE/glyoxalase 1 pathway. Biomed. Pharmacother. 2019, 111, 1166–1175. [Google Scholar] [CrossRef] [PubMed]
- Hajialyani, M.; Hosein Farzaei, M.; Echeverría, J.; Nabavi, S.M.; Uriarte, E.; Sobarzo-Sánchez, E. Hesperidin as a Neuroprotective Agent: A Review of Animal and Clinical Evidence. Molecules 2019, 24, 648. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, S.H.; Yun, J.M. Fisetin inhibits hyperglycemia-induced proinflammatory cytokine production by epigenetic mechanisms. Evid.-Based Complement. Altern. Med. 2012, 2012, 639469. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.; Shangguan, X.; Yin, Z.; Wu, S.; Zhang, Q.; Peng, W.; Li, J.; Zhang, L.; Chen, J. Inhibitory Effect of Fisetin on α-Glucosidase Activity: Kinetic and Molecular Docking Studies. Molecules 2021, 26, 5306. [Google Scholar] [CrossRef]
- Jia, Y.; Ma, Y.; Cheng, G.; Zhang, Y.; Cai, S. Comparative Study of Dietary Flavonoids with Different Structures as α-Glucosidase Inhibitors and Insulin Sensitizers. J. Agric. Food Chem. 2019, 67, 10521–10533. [Google Scholar] [CrossRef]
- Althunibat, O.Y.; Al Hroob, A.M.; Abukhalil, M.H.; Germoush, M.O.; Bin-Jumah, M.; Mahmoud, A.M. Fisetin ameliorates oxidative stress, inflammation and apoptosis in diabetic cardiomyopathy. Life Sci. 2019, 221, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Jia, Q.; Cao, H.; Chen, C.; Xing, S.; Huang, Y.; Shen, D. Fisetin ameliorates atherosclerosis by regulating PCSK9 and LOX-1 in apoE-/- mice. Exp. Ther. Med. 2021, 21, 25. [Google Scholar] [CrossRef] [PubMed]
- JZ, A.L.; BinMowyna, M.N.; AlFaris, N.A.; Alagal, R.I.; El-Kott, A.F.; Al-Farga, A.M. Fisetin protects against streptozotocin-induced diabetic cardiomyopathy in rats by suppressing fatty acid oxidation and inhibiting protein kinase R. Saudi Pharm. J. SPJ Off. Publ. Saudi Pharm. Soc. 2021, 29, 27–42. [Google Scholar]
- Sandireddy, R.; Yerra, V.G.; Komirishetti, P.; Areti, A.; Kumar, A. Fisetin Imparts Neuroprotection in Experimental Diabetic Neuropathy by Modulating Nrf2 and NF-κB Pathways. Cell. Mol. Neurobiol. 2016, 36, 883–892. [Google Scholar] [CrossRef]
- Zhang, S.; Xue, R.; Geng, Y.; Wang, H.; Li, W. Fisetin Prevents HT22 Cells from High Glucose-Induced Neurotoxicity via PI3K/Akt/CREB Signaling Pathway. Front. Neurosci. 2020, 14, 241. [Google Scholar] [CrossRef]
- Dong, W.; Jia, C.; Li, J.; Zhou, Y.; Luo, Y.; Liu, J.; Zhao, Z.; Zhang, J.; Lin, S.; Chen, Y. Fisetin Attenuates Diabetic Nephropathy-Induced Podocyte Injury by Inhibiting NLRP3 Inflammasome. Front. Pharmacol. 2022, 13, 783706. [Google Scholar] [CrossRef]
- Ge, C.; Xu, M.; Qin, Y.; Gu, T.; Lou, D.; Li, Q.; Hu, L.; Nie, X.; Wang, M.; Tan, J. Fisetin supplementation prevents high fat diet-induced diabetic nephropathy by repressing insulin resistance and RIP3-regulated inflammation. Food Funct. 2019, 10, 2970–2985. [Google Scholar] [CrossRef]
- Couillaud, J.; Leydet, L.; Duquesne, K.; Iacazio, G. The Terpene Mini-Path, a New Promising Alternative for Terpenoids Bio-Production. Genes 2021, 12, 1974. [Google Scholar] [CrossRef] [PubMed]
- Chalvin, C.; Drevensek, S.; Gilard, F.; Mauve, C.; Chollet, C.; Morin, H.; Nicol, E.; Héripré, E.; Kriegshauser, L.; Gakière, B.; et al. Sclareol and linalyl acetate are produced by glandular trichomes through the MEP pathway. Hortic. Res. 2021, 8, 206. [Google Scholar] [CrossRef] [PubMed]
- Cerri, G.C.; Lima, L.C.F.; Lelis, D.F.; Barcelos, L.D.S.; Feltenberger, J.D.; Mussi, S.V.; Monteiro-Junior, R.S.; Santos, R.; Ferreira, L.A.M.; Santos, S.H.S. Sclareol-loaded lipid nanoparticles improved metabolic profile in obese mice. Life Sci. 2019, 218, 292–299. [Google Scholar] [CrossRef]
- Chen, H.L.; Gong, J.Y.; Lin, S.C.; Li, S.; Chiang, Y.C.; Hung, J.H.; Yen, C.C.; Lin, C.C. Effects of Sclareol Against Small Cell Lung Carcinoma and the Related Mechanism: In Vitro and In Vivo Studies. Anticancer Res. 2020, 40, 4947–4960. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.; Chiang, Y.F.; Shih, Y.H.; Chiu, C.H.; Chen, H.Y.; Shieh, T.M.; Wang, K.L.; Huang, T.C.; Hong, Y.H.; Hsia, S.M. Salvia sclarea L. Essential Oil Extract and Its Antioxidative Phytochemical Sclareol Inhibit Oxytocin-Induced Uterine Hypercontraction Dysmenorrhea Model by Inhibiting the Ca2+-MLCK-MLC20 Signaling Cascade: An Ex Vivo and In Vivo Study. Antioxidants 2020, 9, 991. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Zhang, J.; Zhou, L.; Wei, J.; Tu, Y.; Shi, Q.; Zhang, Y.; Ren, J.; Wang, Y.; Ying, H.; et al. Sclareol ameliorates hyperglycemia-induced renal injury through inhibiting the MAPK/NF-κB signaling pathway. Phytother. Res. PTR 2022, 36, 2511–2523. [Google Scholar] [CrossRef]
- Du, J.; Chen, C.; Sun, Y.; Zheng, L.; Wang, W. Ponicidin suppresses HT29 cell growth via the induction of G1 cell cycle arrest and apoptosis. Mol. Med. Rep. 2015, 12, 5816–5820. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, J.; Liu, B.; Chen, F.; Li, J.; Liu, Y.; Zhu, J.; Shen, C. Ponicidin inhibits pro-inflammatory cytokine TNF-α-induced epithelial-mesenchymal transition and metastasis of colorectal cancer cells via suppressing the AKT/GSK-3β/Snail pathway. Inflammopharmacology 2019, 27, 627–638. [Google Scholar] [CrossRef]
- Islam, M.T.; Biswas, S.; Bagchi, R.; Khan, M.R.; Khalipha, A.B.R.; Rouf, R.; Uddin, S.J.; Shilpi, J.A.; Bardaweel, S.K.; Sabbah, D.A.; et al. Ponicidin as a promising anticancer agent: Its biological and biopharmaceutical profile along with a molecular docking study. Biotechnol. Appl. Biochem. 2019, 66, 434–444. [Google Scholar] [CrossRef]
- An, S.; Li, Y.; Jia, X.; Yang, Y.; Jia, X.; Jia, X.; Xue, W. Ponicidin attenuates streptozotocin-induced diabetic nephropathy in rats via modulating hyperlipidemia, oxidative stress, and inflammatory markers. J. Biochem. Mol. Toxicol. 2022, 36, e22988. [Google Scholar] [CrossRef]
- Peng, H.; You, L.; Yang, C.; Wang, K.; Liu, M.; Yin, D.; Xu, Y.; Dong, X.; Yin, X.; Ni, J. Ginsenoside Rb1 Attenuates Triptolide-Induced Cytotoxicity in HL-7702 Cells via the Activation of Keap1/Nrf2/ARE Pathway. Front. Pharmacol. 2021, 12, 723784. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Tang, X.; Ke, X.; Dai, Y.; Shi, J. Triptolide Suppresses NF-κB-Mediated Inflammatory Responses and Activates Expression of Nrf2-Mediated Antioxidant Genes to Alleviate Caerulein-Induced Acute Pancreatitis. Int. J. Mol. Sci. 2022, 23, 1252. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; He, H.; Zhang, Y.; Fan, J.; Wang, L. Mechanisms of action of triptolide against colorectal cancer: Insights from proteomic and phosphoproteomic analyses. Aging 2022, 14, 3084–3104. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Shen, W.; Qin, W.; Zheng, C.; Zhang, M.; Zeng, C.; Wang, S.; Wang, J.; Zhu, X.; Liu, Z. Treatment of db/db diabetic mice with triptolide: A novel therapy for diabetic nephropathy. Nephrol. Dial. Transplant. Off. Publ. Eur. Dial. Transpl. Assoc. Eur. Ren. Assoc. 2010, 25, 3539–3547. [Google Scholar] [CrossRef]
- Liang, D.; Mai, H.; Ruan, F.; Fu, H. The Efficacy of Triptolide in Preventing Diabetic Kidney Diseases: A Systematic Review and Meta-Analysis. Front. Pharmacol. 2021, 12, 728758. [Google Scholar] [CrossRef]
- Iwata, Y.; Furuichi, K.; Hashimoto, S.; Yokota, K.; Yasuda, H.; Sakai, N.; Kitajima, S.; Toyama, T.; Shinozaki, Y.; Sagara, A.; et al. Pro-inflammatory/Th1 gene expression shift in high glucose stimulated mesangial cells and tubular epithelial cells. Biochem. Biophys. Res. Commun. 2014, 443, 969–974. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Pan, C.; Chang, B.; Wu, X.; Guo, J.; Zhou, Y.; Liu, H.; Zhu, Z.; Chang, B.; Chen, L. Triptolide Improves Diabetic Nephropathy by Regulating Th Cell Balance and Macrophage Infiltration in Rat Models of Diabetic Nephropathy. Exp. Clin. Endocrinol. Diabetes Off. J. Ger. Soc. Endocrinol. Ger. Diabetes Assoc. 2016, 124, 389–398. [Google Scholar] [CrossRef]
- Han, F.; Xue, M.; Chang, Y.; Li, X.; Yang, Y.; Sun, B.; Chen, L. Triptolide Suppresses Glomerular Mesangial Cell Proliferation in Diabetic Nephropathy Is Associated with Inhibition of PDK1/Akt/mTOR Pathway. Int. J. Biol. Sci. 2017, 13, 1266–1275. [Google Scholar] [CrossRef]
- Li, X.Y.; Wang, S.S.; Han, Z.; Han, F.; Chang, Y.P.; Yang, Y.; Xue, M.; Sun, B.; Chen, L.M. Triptolide Restores Autophagy to Alleviate Diabetic Renal Fibrosis through the miR-141-3p/PTEN/Akt/mTOR Pathway. Mol. Ther. Nucleic Acids 2017, 9, 48–56. [Google Scholar] [CrossRef]
- Xue, M.; Cheng, Y.; Han, F.; Chang, Y.; Yang, Y.; Li, X.; Chen, L.; Lu, Y.; Sun, B.; Chen, L. Triptolide Attenuates Renal Tubular Epithelial-mesenchymal Transition via the MiR-188-5p-mediated PI3K/AKT Pathway in Diabetic Kidney Disease. Int. J. Biol. Sci. 2018, 14, 1545–1557. [Google Scholar] [CrossRef]
- Shi, G.; Wu, W.; Wan, Y.G.; Hex, H.W.; Tu, Y.; Han, W.B.; Liu, B.H.; Liu, Y.L.; Wan, Z.Y. Low dose of triptolide ameliorates podocyte epithelial-mesenchymal transition induced by high dose of D-glucose via inhibiting Wnt3α/β-catenin signaling pathway activation. China J. Chin. Mater. Med. 2018, 43, 139–146. [Google Scholar]
- Wu, W.; Liu, B.H.; Wan, Y.G.; Sun, W.; Liu, Y.L.; Wang, W.W.; Fang, Q.J.; Tu, Y.; Yee, H.Y.; Yuan, C.C.; et al. Triptolide inhibits NLRP3 inflammasome activation and ameliorates podocyte epithelial-mesenchymal transition induced by high glucose. China J. Chin. Mater. Med. 2019, 44, 5457–5464. [Google Scholar]
- Ren, L.; Wan, R.; Chen, Z.; Huo, L.; Zhu, M.; Yang, Y.; Chen, Q.; Zhang, X.; Wang, X. Triptolide Alleviates Podocyte Epithelial-Mesenchymal Transition via Kindlin-2 and EMT-Related TGF-β/Smad Signaling Pathway in Diabetic Kidney Disease. Appl. Biochem. Biotechnol. 2022, 194, 1000–1012. [Google Scholar] [CrossRef] [PubMed]
- Xi, C.; Peng, S.; Wu, Z.; Zhou, Q.; Zhou, J. Toxicity of triptolide and the molecular mechanisms involved. Biomed. Pharmacother. 2017, 90, 531–541. [Google Scholar] [CrossRef]
- Singh, D.; Chaudhuri, P.K. Structural characteristics, bioavailability and cardioprotective potential of saponins. Integr. Med. Res. 2018, 7, 33–43. [Google Scholar] [CrossRef]
- Yang, W.Z.; Hu, Y.; Wu, W.Y.; Ye, M.; Guo, D.A. Saponins in the genus Panax L. (Araliaceae): A systematic review of their chemical diversity. Phytochemistry 2014, 106, 7–24. [Google Scholar] [CrossRef]
- Cai, J.; Liu, J.; Fan, P.; Dong, X.; Zhu, K.; Liu, X.; Zhang, N.; Cao, Y. Dioscin prevents DSS-induced colitis in mice with enhancing intestinal barrier function and reducing colon inflammation. Int. Immunopharmacol. 2021, 99, 108015. [Google Scholar] [CrossRef]
- Kang, K.B.; Ryu, J.; Cho, Y.; Choi, S.Z.; Son, M.; Sung, S.H. Combined Application of UHPLC-QTOF/MS, HPLC-ELSD and (1) H-NMR Spectroscopy for Quality Assessment of DA-9801, A Standardised Dioscorea Extract. Phytochem. Anal. PCA 2017, 28, 185–194. [Google Scholar] [CrossRef]
- Wang, J.; Yang, G.Y.; Sun, H.Y.; Meng, T.; Cheng, C.C.; Zhao, H.P.; Luo, X.L.; Yang, M.M. Dioscin Reduces Vascular Damage in the Retina of db/db Mice by Inhibiting the VEGFA Signaling Pathway. Front. Pharmacol. 2022, 12, 811897. [Google Scholar] [CrossRef]
- Xu, L.N.; Yin, L.H.; Jin, Y.; Qi, Y.; Han, X.; Xu, Y.W.; Liu, K.X.; Zhao, Y.Y.; Peng, J.Y. Effect and possible mechanisms of dioscin on ameliorating metabolic glycolipid metabolic disorder in type-2-diabetes. Phytomed. Int. J. Phytother. Phytopharm. 2020, 67, 153139. [Google Scholar] [CrossRef]
- Cai, S.; Chen, J.; Li, Y. Dioscin protects against diabetic nephropathy by inhibiting renal inflammation through TLR4/NF-κB pathway in mice. Immunobiology 2020, 225, 151941. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Liu, J.; Sun, D.; Guo, T.; Yao, Y.; Xia, X.; Shi, C.; Peng, X. Dioscin relieves diabetic nephropathy via suppressing oxidative stress and apoptosis, and improving mitochondrial quality and quantity control. Food Funct. 2022, 13, 3660–3673. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Li, D.; Gao, X.; Li, Y.; Wu, J.; Zou, W. Di’ao Xinxuekang Capsule, a Chinese Medicinal Product, Decreases Serum Lipids Levels in High-Fat Diet-Fed ApoE(-/-) Mice by Downregulating PCSK9. Front. Pharmacol. 2018, 9, 1170. [Google Scholar] [CrossRef]
- Li, K.; Tang, Y.; Fawcett, J.P.; Gu, J.; Zhong, D. Characterization of the pharmacokinetics of dioscin in rat. Steroids 2005, 70, 525–530. [Google Scholar] [CrossRef]
- Li, X.; Liu, S.; Qu, L.; Chen, Y.; Yuan, C.; Qin, A.; Liang, J.; Huang, Q.; Jiang, M.; Zou, W. Dioscin and diosgenin: Insights into their potential protective effects in cardiac diseases. J. Ethnopharmacol. 2021, 274, 114018. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Zhang, S.; Zheng, L.; Yin, L.; Xu, L.; Peng, J. A 90-day subchronic toxicological assessment of dioscin, a natural steroid saponin, in Sprague–Dawley rats. Food Chem. Toxicol. 2012, 50, 1279–1287. [Google Scholar] [CrossRef]
- Zhou, P.; Xie, W.; He, S.; Sun, Y.; Meng, X.; Sun, G.; Sun, X. Ginsenoside Rb1 as an Anti-Diabetic Agent and Its Underlying Mechanism Analysis. Cells 2019, 8, 204. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Liu, P.; Wang, H.; Dong, M.; Li, G.; Li, Y. Ginsenoside Rb1 attenuates diabetic retinopathy in streptozotocin-induced diabetic rats1. Acta Cir. Bras. 2019, 34, e201900201. [Google Scholar] [CrossRef] [PubMed]
- Tabandeh, M.R.; Hosseini, S.A.; Hosseini, M. Ginsenoside Rb1 exerts antidiabetic action on C2C12 muscle cells by leptin receptor signaling pathway. J. Recept. Signal Transduct. Res. 2017, 37, 370–378. [Google Scholar] [CrossRef]
- He, J.Y.; Hong, Q.; Chen, B.X.; Cui, S.Y.; Liu, R.; Cai, G.Y.; Guo, J.; Chen, X.M. Ginsenoside Rb1 alleviates diabetic kidney podocyte injury by inhibiting aldose reductase activity. Acta Pharmacol. Sin. 2022, 43, 342–353. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Oh, T.K.; Kim, Y.H.; Lee, J.; Moon, J.M.; Park, Y.S.; Sung, C.M. Pharmacokinetics of Ginsenoside Rb1, Rg3, Rk1, Rg5, F2, and Compound K from Red Ginseng Extract in Healthy Korean Volunteers. Evid.-Based Complement. Altern. Med. 2022, 2022, 8427519. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Jeon, J.-H.; Lee, S.; Kang, W.Y.; Seong, S.J.; Yoon, Y.-R.; Choi, M.-K.; Song, I.-S. Detection of 13 Ginsenosides (Rb1, Rb2, Rc, Rd, Re, Rf, Rg1, Rg3, Rh2, F1, Compound K, 20(S)-Protopanaxadiol, and 20(S)-Protopanaxatriol) in Human Plasma and Application of the Analytical Method to Human Pharmacokinetic Studies Following Two Week-Repeated Administration of Red Ginseng Extract. Molecules 2019, 24, 2618. [Google Scholar]
- Yang, R.; Pei, T.; Huang, R.; Xiao, Y.; Yan, J.; Zhu, J.; Zheng, C.; Xiao, W.; Huang, C. Platycodon grandiflorum Triggers Antitumor Immunity by Restricting PD-1 Expression of CD8+ T Cells in Local Tumor Microenvironment. Front. Pharmacol. 2022, 13, 774440. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Huang, D.; Wang, X.; Pei, C.; Xiao, W.; Wang, F.; Wang, Z. Suppression of NLRP3 inflammasome by Platycodin D via the TLR4/MyD88/NF-κB pathway contributes to attenuation of lipopolysaccharide induced acute lung injury in rats. Int. Immunopharmacol. 2021, 96, 107621. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.L.; Park, J.; Jung, Y.; Ahn, K.S.; Um, J.Y. Platycodin D, a novel activator of AMP-activated protein kinase, attenuates obesity in db/db mice via regulation of adipogenesis and thermogenesis. Phytomed. Int. J. Phytother. Phytopharm. 2019, 52, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.M.; Cong, S.; Cheng, Z.; Hu, Y.X.; Lei, Y.; Zhu, L.L.; Zhao, X.K.; Mu, M.; Zhang, B.F.; Fan, L.D.; et al. Platycodin D alleviates liver fibrosis and activation of hepatic stellate cells by regulating JNK/c-JUN signal pathway. Eur. J. Pharm. 2020, 876, 172946. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Fu, L.; Zhao, Y.; Ke, W.; Zhuang, Y. Platycodin D improves renal injury in diabetic nephropathy model rats by regulating oxidative stress mediated PI3K/Akt/mTOR signaling pathway. Chin. J. Pharmacol. Toxicol. 2022, 36, 170–176. [Google Scholar]
- Shen, S.; Ji, C.; Wei, K. Cellular Senescence and Regulated Cell Death of Tubular Epithelial Cells in Diabetic Kidney Disease. Front. Endocrinol. 2022, 13, 924299. [Google Scholar] [CrossRef]
- Huang, J.; Chen, G.; Wang, J.; Liu, S.; Su, J. Platycodin D regulates high glucose-induced ferroptosis of HK-2 cells through glutathione peroxidase 4 (GPX4). Bioengineered 2022, 13, 6627–6637. [Google Scholar] [CrossRef]
- Lee, W.H.; Gam, C.O.; Ku, S.K.; Choi, S.H. Single oral dose toxicity test of platycodin d, a saponin from platycodin radix in mice. Toxicol. Res. 2011, 27, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.Y.; Jiang, X.M.; Xu, Y.L.; Yuan, L.W.; Chen, Y.C.; Cui, G.; Huang, R.Y.; Liu, B.; Wang, Y.; Chen, X.; et al. Platycodin D triggers the extracellular release of programed death Ligand-1 in lung cancer cells. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2019, 131, 110537. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Yang, T.; Zhao, S.; Zheng, Q.; Li, Y.; Zhang, Z.; Sun, X.; Liu, Y.; Zhang, Y.; Xie, J. Distribution, Biotransformation, Pharmacological Effects, Metabolic Mechanism and Safety Evaluation of Platycodin D: A Comprehensive Review. Curr. Drug Metab. 2022, 23, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Dieter, S.; Paula, L.; Maria, B.; Victor, W.; Lutz, G. Hydroxycinnamic acid esters of isocitric acid: Accumulation and enzymatic synthesis in Amaranthus cruentus. Phytochemistry 1987, 26, 2919–2922. [Google Scholar] [CrossRef]
- Yao, H.; Zhang, W.; Yang, F.; Ai, F.; Du, D.; Li, Y. Discovery of caffeoylisocitric acid as a Keap1-dependent Nrf2 activator and its effects in mesangial cells under high glucose. J. Enzym. Inhib. Med. Chem. 2022, 37, 178–188. [Google Scholar] [CrossRef]
- Bastani, S.; Vahedian, V.; Rashidi, M.; Mir, A.; Mirzaei, S.; Alipourfard, I.; Pouremamali, F.; Nejabati, H.; Kadkhoda, J.; Maroufi, N.F.; et al. An evaluation on potential anti-oxidant and anti-inflammatory effects of Crocin. Biomed. Pharmacother. 2022, 153, 113297. [Google Scholar] [CrossRef]
- Roshanravan, B.; Samarghandian, S.; Ashrafizadeh, M.; Amirabadizadeh, A.; Saeedi, F.; Farkhondeh, T. Metabolic impact of saffron and crocin: An updated systematic and meta-analysis of randomised clinical trials. Arch. Physiol. Biochem. 2022, 128, 666–678. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, X.; Zhu, H.; Wang, J.; Ma, J.; Gu, M. Crocin protects the renal tubular epithelial cells against high glucose-induced injury and oxidative stress via regulation of the SIRT1/Nrf2 pathway. Iran. J. Basic Med. Sci. 2022, 25, 193–197. [Google Scholar]
- Qiu, Y.; Jiang, X.; Liu, D.; Deng, Z.; Hu, W.; Li, Z.; Li, Y. The Hypoglycemic and Renal Protection Properties of Crocin via Oxidative Stress-Regulated NF-κB Signaling in db/db Mice. Front. Pharmacol. 2020, 11, 541. [Google Scholar] [CrossRef]
- Zhang, L.; Jing, M.; Liu, Q. Crocin alleviates the inflammation and oxidative stress responses associated with diabetic nephropathy in rats via NLRP3 inflammasomes. Life Sci. 2021, 278, 119542. [Google Scholar] [CrossRef] [PubMed]
- Sepahi, S.; Golfakhrabadi, M.; Bonakdaran, S.; Lotfi, H.; Mohajeri, S.A. Effect of crocin on diabetic patients: A placebo-controlled, triple-blinded clinical trial. Clin. Nutr. ESPEN 2022, 50, 255–263. [Google Scholar] [CrossRef]
- Hosseinzadeh, H.; Shariaty, V.; Sameni, A.; Vahabzadeh, M. Acute and sub-acute toxicity of crocin, a constituent of Crocus sativus L. (saffron), in mice and rats. Pharmacologyonline 2010, 2, 943–951. [Google Scholar]
- Nam, P.C.; Thong, N.M.; Hoa, N.T.; Quang, D.T.; Hoang, L.P.; Mechler, A.; Vo, Q.V. Is natural fraxin an overlooked radical scavenger? RSC Adv. 2021, 11, 14269–14275. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Zeng, J.; Li, C.; Xiao, H.; Li, S.; Lin, Z.; Huang, K.; Shen, J.; Huang, H. Fraxin Promotes the Activation of Nrf2/ARE Pathway via Increasing the Expression of Connexin43 to Ameliorate Diabetic Renal Fibrosis. Front. Pharmacol. 2022, 13, 853383. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Natural Compound | Model | Function | Mechanism/Target | Reference |
|---|---|---|---|---|
| Oleuropein | In vivo: db/db mice DKD | Antioxidant Anti-inflammatory Reduce apoptosis | MAPK signaling pathway, caspase-3, Bcl-2, Bax | [50] |
| Resveratrol | In vitro: HG stimulated NRK 52E cells | Anti-ER stress Reduce apoptosis | GRP78, CHOP | [57] |
| In vitro: HG stimulated mouse podocytes In vivo: db/db mice DKD | Antioxidant Reduce apoptosis | AMPK signaling pathway | [58] | |
| In vitro: HG stimulated mouse podocytes In vivo: db/db mice DKD | Antioxidant Reduce apoptosis | SIRT1, PGC-1α, NRF1, TFAM | [59] | |
| In vitro: HG stimulated SV40-MES-13 In vivo: STZ-induced mice DKD | Antioxidant Inhibit proliferation Anti-fibrotic | Keap1/Nrf2 signaling pathway | [60] | |
| In vitro: HG stimulated CRL-2573 In vivo: STZ-induced rats DKD | Anti-fibrotic | MAPK/TGF-β1 signaling pathway | [61] | |
| In vivo: STZ-induced mice DKD | Reduce lipid accumulation | SIRT1 signaling pathway | [62] | |
| In vivo: STZ-induced rats DKD | Reduce lipid accumulation Improve autophagy | AMPK/mTOR signaling pathway PPARα, CPT1, SREBP-1c, ACS | [63] | |
| Gastrodin | In vitro: HG stimulated MPC-5 cells | Antioxidant Anti-inflammatory Reduce apoptosis | AMPK/Nrf2 signaling pathway NLRP3 inflammasome | [71] |
| Natural Compound | Model | Function | Mechanism/Target | Reference |
|---|---|---|---|---|
| Trigonelline | In vitro: HG stimulated HMCs In vivo: STZ-induced rats DKD | Antioxidant Reduce apoptosis Anti-fibrotic | Wnt/β-catenin signaling pathway FN, collagen IV | [78,79] |
| In vivo: STZ-induced rats DKD | Antioxidant Anti-inflammatory Reduce apoptosis | PPARγ/GLUT4-leptin/TNF-α signaling pathway | [80] | |
| In vitro: HG stimulated HMCs | Improve autophagy | miR-5189-5p, HIF1AN AMPK signaling pathway | [81] | |
| Berberine | In vitro: HG stimulated rat podocytes In vivo: STZ-induced rats DKD | Alleviate podocyte injury | PI3K/Akt signaling pathway nephrin, podocin, α3β1 | [88] |
| In vitro: HG stimulated GMCs In vivo: STZ-induced mice DKD | Reduce glucose uptake Inhibit proliferation | PI3K/Akt/AS160/GLUT1 signaling pathway | [89] | |
| In vitro: hypoxia/HG-stimulated NRK 52E and HK-2 cells | Reduce apoptosis | PI3K/Akt signaling pathway HIF-1α | [90] | |
| In vitro: PA stimulated HK-2 cells | Reduce apoptosis Reduce lipid accumulation | CPT1A, PPARα, PGC1α | [91] | |
| In vitro: HG stimulated HK-2 cells In vivo: db/db mice DKD | Anti-fibrotic Reduce lipid accumulation Improve mitochondrial function | α-SMA, collagen I, collagen IV, FN, TGF-β1, CPT1, ACOX1, PPARα, AMPK, PGC-1α | [92] | |
| In vitro: HG stimulated HK-2 cells In vivo: STZ-induced rats DKD | Antioxidant Reduce apoptosis Improve mitochondrial function | C/EBPβ/Gas5/miR-18a-5p signaling pathway C/EBPβ/PGC-1α signaling pathway | [93] | |
| In vitro: PA stimulated mouse podocytes In vivo: db/db mice DKD | Antioxidant Reduce lipid accumulation Improve mitochondrial function | AMPK/PGC-1α signaling pathway CPT1, ACC, CD36 | [94] | |
| In vitro: PA stimulated mouse podocytes In vivo: db/db mice DKD | Antioxidant Reduce apoptosis Improve mitochondrial function | Drp1, MFF, Fis1, Mid49, Mid51 | [95] | |
| In vitro: HG stimulated mouse podocytes In vivo: STZ-induced rats DKD | Anti-inflammatory Reduce apoptosis | TLR4/NF-κB signaling pathway | [97] | |
| In vitro: HG stimulated mRTEC In vivo: KKAy mice DKD | Anti-fibrotic | Notch/snail signaling pathway α-SMA, E-cadherin | [96] | |
| In vitro: HG stimulated mouse podocytes | Reduce apoptosis Improve autophagy | mTOR/P70S6K/4EBP1 signaling pathway | [98] | |
| Sinomenine | In vitro: HG stimulated HRGEs | Antioxidant Decrease cell permeability | Nrf2 signaling pathway ROCK signaling pathway ZO-1, occludin | [105] |
| In vitro: HG stimulated HRGEs In vivo: STZ-induced rats DKD | Anti-inflammatory Decrease cell permeability | C/EBPα/claudin-5 signaling pathway | [106] | |
| In vitro: H2O2 stimulated HK-2 cells In vivo: STZ-induced rats DKD | Antioxidant Reduce apoptosis Anti-inflammatory Anti-fibrotic | JAK/STAT signaling pathway | [107] |
| Natural Compound | Model | Function | Mechanism/Target | Reference |
|---|---|---|---|---|
| Naringenin | In vitro: HG stimulated NRK 52E cells In vivo: STZ-induced rats DKD | Antioxidant Anti-ER stress Reduce apoptosis | p-PERK, eIF2α, XBP1s, ATF4, CHOP | [117] |
| In vitro: HG stimulated NRK 52E cells In vivo: STZ-induced mice DKD | Reduce renal tissue injury | PPARs, CYP4A, 20-HETE | [118] | |
| In vitro: HG stimulated 293T cells In vivo: STZ-induced rats DKD | Anti-fibrotic | MicroRNA let-7a TGF-β1/Smad signaling pathway | [119] | |
| Quercetin | In vivo: STZ-induced rats DKD | Antioxidant Anti-inflammatory | AGEs, TNF-α, IL-6 | [125] |
| In vitro: HG stimulated mouse podocytes In vivo: db/db mice DKD | Reduce apoptosis | EGFR signaling pathway | [126] | |
| In vitro: HG stimulated HMCs In vivo: DKD patients | Inhibit proliferation Antioxidant Anti-inflammatory | miR-485-5p, YAP1 | [127] | |
| In vitro: HG stimulated SV40-MES-13 In vivo: db/db mice DKD | Inhibit proliferation Antioxidant Anti-inflammatory | Hippo signaling pathway | [128] | |
| In vivo: db/db mice DKD | Reduce lipid accumulation | SCAP-SREBP2-LDLr signaling pathway | [130] | |
| In vivo: STZ-induced rats DKD | Reduce lipid accumulation Anti-inflammatory | PPARα, CPT1, OCTN2, ACC2 NLRP3 inflammasome/caspase-1/IL-1β/IL-18 signaling pathway | [131] | |
| Icariin | In vitro: HG stimulated MPC-5 cells In vivo: STZ-induced rats DKD | Anti-inflammatory Improve autophagy Improve mitochondrial function | Sesn2, NLRP3 Nrf2 signaling pathway | [134] |
| In vivo: STZ-induced mice DKD | Anti-inflammatory Anti-fibrotic | TLR4/NF-κB signaling pathway | [135] | |
| In vitro: HG stimulated SV40-MES-13 | Anti-fibrotic | TGF-β1/Smad signaling pathway ERK, GPER | [136] | |
| In vitro: HG stimulated HK-2 and NRK 49F cells In vivo: STZ-induced rats DKD | Improve autophagy Anti-fibrotic | miR-192-5p/GLP-1R signaling pathway p-mTOR, collagen I, α-SMA, FN | [137] | |
| In vitro: HG stimulated NRK 52E cells In vivo: STZ-induced rats DKD | Reduce apoptosis Anti-fibrotic | miR-122-5p, FOXP2, E-cadherin, α-SMA | [138] | |
| Cardamonin | In vitro: MGO stimulated NRK 52E cells In vivo: STZ-induced rats DKD | Antioxidant Anti-inflammatory Reduce apoptosis Anti-fibrotic | PI3K/AKT signaling pathway JAK/STAT signaling pathway caspase-3, Bcl-2, Bax, NF-κB, FN, α-SMA, TGF-β1, Vimentin | [146] |
| Morin | In vitro: HG stimulated NRK 52E cells In vivo: STZ-induced rats DKD | Antioxidant Reduce apoptosis | PHLPP1/FoxO1-Mdm2 signaling pathway Nrf2 | [150] |
| In vitro: HG stimulated primary rat GMCs | Antioxidant Anti-fibrotic | MAPK signaling pathway JNK signaling pathway NOX4 | [151] | |
| Hesperetin | In vivo: STZ-induced rats DKD | Antioxidant Anti-inflammatory Anti-fibrotic | TBARS, GSH-Px, CAT, TNF-α, IL-6, TGF-β, GSK-3β | [160] |
| In vivo: STZ-induced rats DKD | Anti-inflammatory Anti-fibrotic | Nrf2 signaling pathway Glo-1, collagen ΙV, FN | [161] | |
| Fisetin | In vitro: HG stimulated mouse podocytes In vivo: STZ-induced mice DKD | Anti-inflammatory Anti-fibrotic Improve autophagy | CDKN1B/P70S6K signaling pathway NLRP3 inflammasome | [171] |
| In vitro: PA stimulated HK-2 cells In vivo: HFD-induced mice kidney injury | Improve insulin sensitivity Anti-inflammatory | Insulin receptor signaling pathway NF-κB signaling pathway RIP3/NLRP3 signaling pathway | [172] |
| Natural Compound | Model | Function | Mechanism/Target | Reference |
|---|---|---|---|---|
| Sclareol | In vitro: HG stimulated SV40-MES-13 In vivo: STZ-induced mice DKD | Antioxidant Anti-inflammatory Anti-fibrotic | MAPK/NF-κB signaling pathway | [178] |
| Ponicidin | In vivo: STZ-induced rats DKD | Antioxidant Anti-inflammatory Improve lipid metabolism Anti-fibrotic | TNF-α, IL-1β, IL-6, NF-κB | [182] |
| Triptolide | In vivo: STZ-induced rats DKD | Anti-inflammatory Regulate Th1/Th2 cells balance | Interferon-γ, IL-12, TNF-α, IL-4, IL-10 | [189] |
| In vitro: HG stimulated HMCs and HK-2 cells In vivo: STZ-induced rats DKD | Improve autophagy Inhibit proliferation Anti-fibrotic | miR-141-3p, miR-188-5p, PTEN, PDK1, Akt, mTOR | [190,191,192] | |
| In vitro: HG stimulated mouse podocytes | Anti-inflammatory Anti-fibrotic | Wnt3α/β-catenin signaling pathway NLRP3, ASC, caspase-1, IL-1β, IL-18 | [193,194] | |
| In vitro: HG stimulated mouse podocytes In vivo: db/db mice DKD | Anti-fibrotic | TGF-β/Smad signaling pathway kindlin-2 | [195] |
| Natural Compound | Model | Function | Mechanism/Target | Reference |
|---|---|---|---|---|
| Dioscin | In vivo: STZ-induced mice DKD | Anti-inflammatory | TLR4/NF-κB signaling pathway | [203] |
| In vivo: STZ-induced rats DKD | Antioxidant Anti-inflammatory Anti-ER stress Reduce apoptosis Improve autophagy Improve mitochondrial function | IL-1β, IL-6, TNF-α, NF-κB, p-PERK, IRE1, p-IRE1, ATF4, CHOP, Caspase-12, PINK1, Drp1, p-Drp1, MFN2 AMPK/mTOR signaling pathway | [204] | |
| Ginsenoside Rb1 | In vivo: STZ-induced rats DKD | Antioxidant Anti-fibrotic | Wnt/β-catenin signaling pathway miR-3550 | [78] |
| In vitro: HG stimulated mouse podocytes In vivo: STZ-induced mice DKD | Antioxidant Protect mitochondria Reduce apoptosis | AR, NOX4, caspase-9 | [212] | |
| Platycodin D | In vivo: STZ-induced rats DKD | Antioxidant Anti-inflammatory Anti-fibrotic | PI3K/Akt signaling pathway | [219] |
| In vitro: HG stimulated HMCs and HK-2 cells | Antioxidant Reduce ferroptosis | GPX4, FTH1, SLC7A11, ACSL4, TFR1 | [221] |
| Natural Compound | Model | Function | Mechanism/Target | Reference |
|---|---|---|---|---|
| Caffeoylisocitric acid | In vitro: HG stimulated HMCs | Antioxidant Anti-inflammatory Anti-fibrotic | Nrf2 signaling pathway MAPK signaling pathway | [226] |
| Crocin | In vitro: HG stimulated HK-2 cells | Antioxidant Reduce apoptosis | SIRT1/Nrf2 signaling pathway | [229] |
| In vivo: db/db mice DKD | Antioxidant Anti-inflammatory | Nrf2, SOD-1, HO-1, CAT NF-κB signaling pathway | [230] | |
| In vivo: STZ-induced rats DKD | Antioxidant Anti-inflammatory Anti-fibrotic | NLRP3 inflammasome TNF-α, IL-1β, IL-18, TGF-β, collagen I, collagen IV | [231] | |
| Fraxin | In vitro: HG stimulated primary GMCs In vivo: db/db mice DKD | Antioxidant Anti-inflammatory Anti-fibrotic | Nrf2 signaling pathway Cx43, Akt, FN, ICAM-1 | [235] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, K.; Zi, X.; Song, J.; Zhao, Q.; Liu, J.; Bao, H.; Li, L. Molecular Mechanistic Pathways Targeted by Natural Compounds in the Prevention and Treatment of Diabetic Kidney Disease. Molecules 2022, 27, 6221. https://doi.org/10.3390/molecules27196221
Zhou K, Zi X, Song J, Zhao Q, Liu J, Bao H, Li L. Molecular Mechanistic Pathways Targeted by Natural Compounds in the Prevention and Treatment of Diabetic Kidney Disease. Molecules. 2022; 27(19):6221. https://doi.org/10.3390/molecules27196221
Chicago/Turabian StyleZhou, Kaixuan, Xue Zi, Jiayu Song, Qiulu Zhao, Jia Liu, Huiwei Bao, and Lijing Li. 2022. "Molecular Mechanistic Pathways Targeted by Natural Compounds in the Prevention and Treatment of Diabetic Kidney Disease" Molecules 27, no. 19: 6221. https://doi.org/10.3390/molecules27196221