
Heterologous Expression and Catalytic Properties of Codon-Optimized Small-Sized Bromelain from MD2 Pineapple

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Gene Optimization, Synthesis and Expression System Construction
2.2. Protein Expression
2.3. Solubilization and Refolding of Insoluble Protein
2.4. Protein Purification
2.5. SDS-PAGE
2.6. Enzymatic Activity and Kinetic Parameters
2.7. Optimum Temperature and pH
2.8. Determination of EDTA and Metal Ions Effect
2.9. Antioxidant Activity
3. Results
3.1. Gene Optimization
3.2. Protein Expression and Solubilization
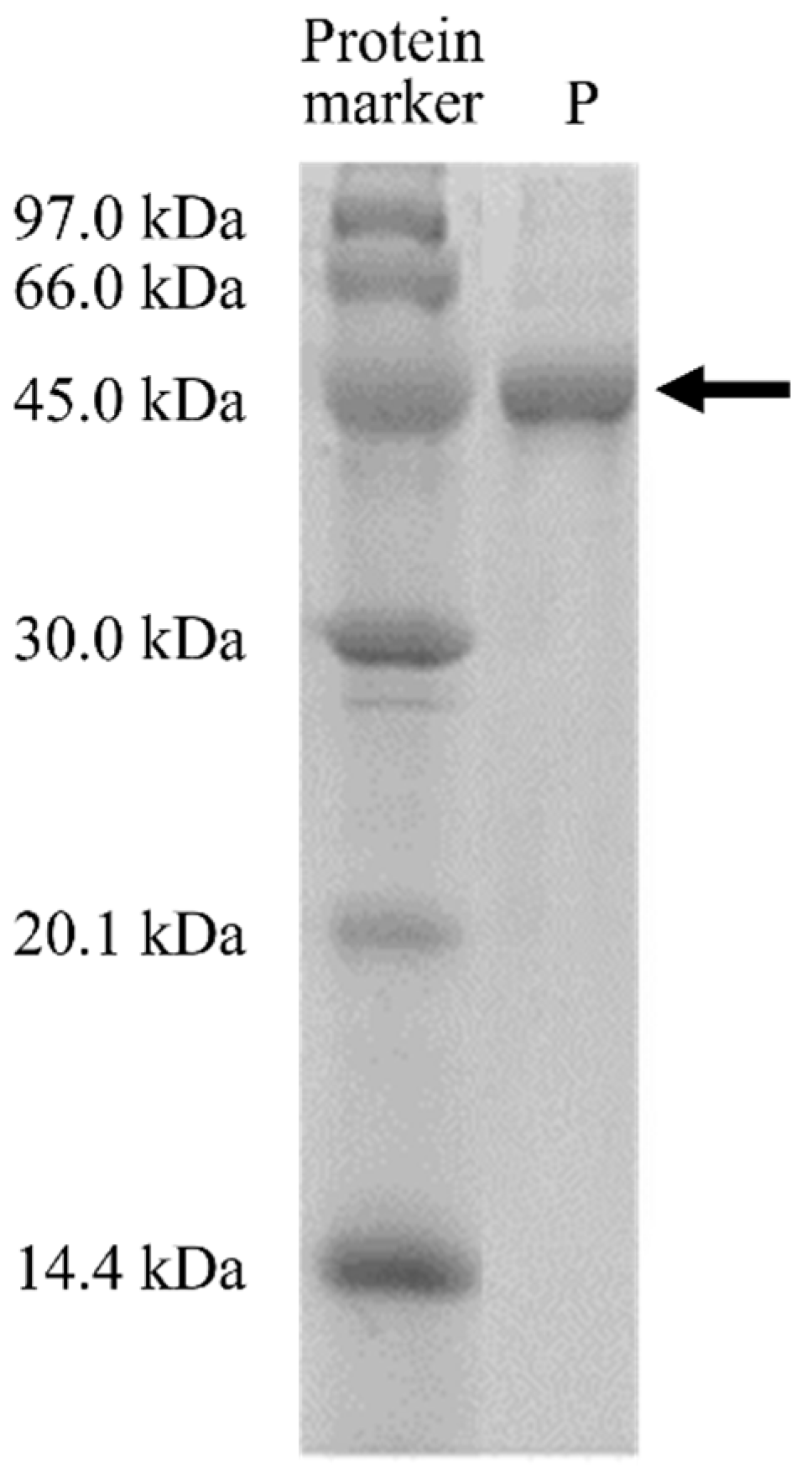
3.3. Protein Purification
3.4. Enzymatic Activity and Kinetic Parameters
3.5. Optimum Temperature and pH
3.6. Effect of EDTA and Metal Ions
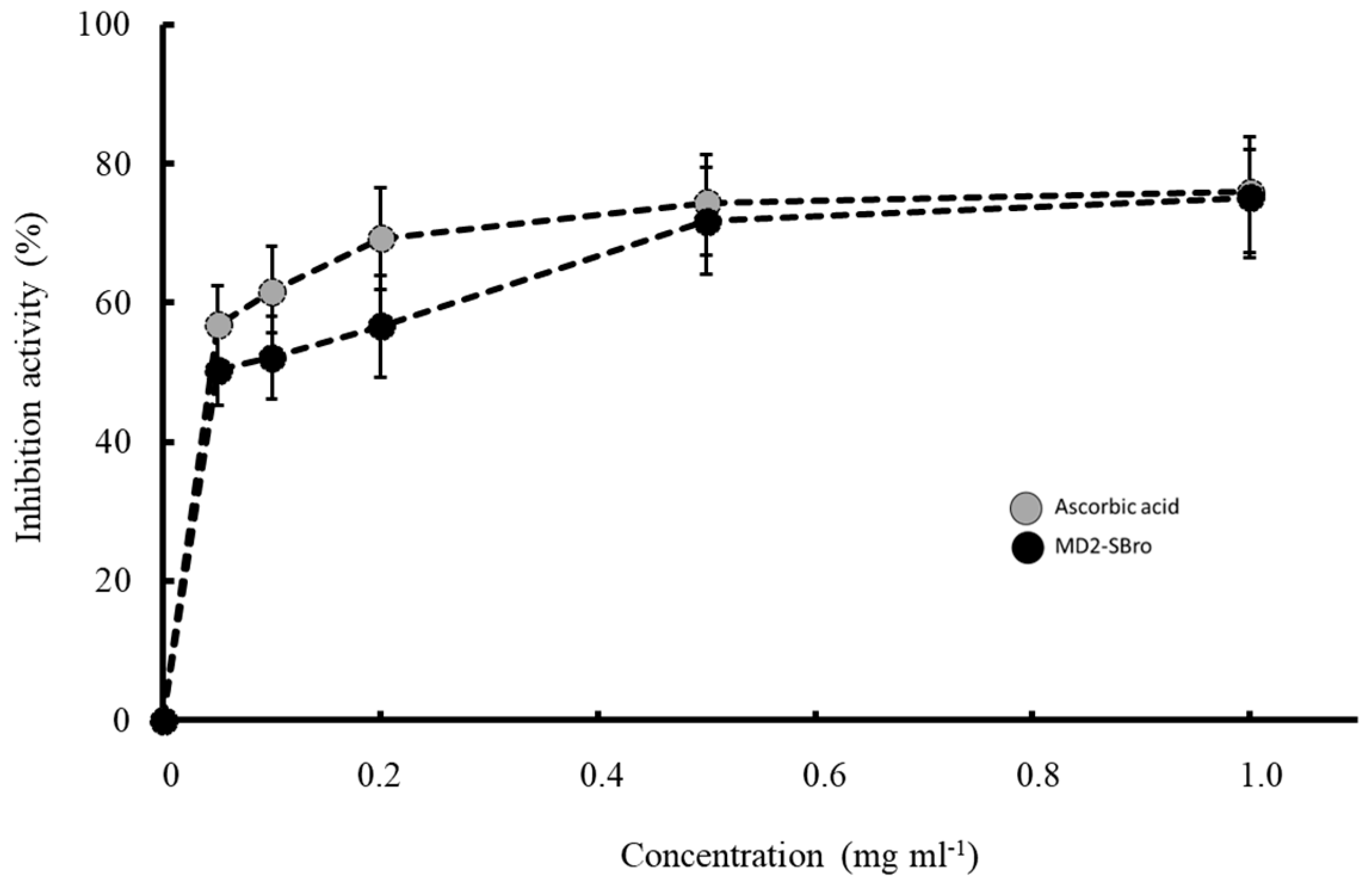
3.7. Antioxidant Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Razali, R.; Budiman, C.; Kamaruzaman, K.A.; Subbiah, V.K. Soluble expression and catalytic properties of codon-optimized recombinant bromelain form MD2 pineapple in Escherichia coli. Protein J. 2021, 40, 406–418. [Google Scholar] [CrossRef] [PubMed]
- Menard, R.; Carrière, J.; Laflamme, P.; Plouffe, C.; Khouri, H.E.; Vernet, T.; Tessier, D.C.; Thomas, D.Y.; Storer, D.C. Contribution of the glutamine 19 side chain to transition-state stabilization in the oxyanion hole of papain. Biochemistry 1991, 30, 8924–8928. [Google Scholar] [CrossRef] [PubMed]
- Otto, H.H.; Schirmeister, T. Cysteine proteases and their inhibitors. Chem. Rev. 1997, 97, 133–171. [Google Scholar] [CrossRef] [PubMed]
- Ketnawa, S.; Chaiwut, P.; Rawdkuen, S. Pineapple wastes: A potential source for bromelain extraction. Food Bioprod. Process. 2012, 90, 385–391. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J. Families of cysteine peptidases. Methods Enzymol. 1994, 244, 461–486. [Google Scholar]
- Arribére, M.P.; Caffin, O.; Priolo, S. Proteolytic enzymes from the latex of Ficus pumila L. (Moraceae). Acta Farm. Bonaer. 2000, 19, 257–262. [Google Scholar]
- Razali, R.; Asis, H.; Budiman, C. Structure-function characteristics of SARS-CoV-2 proteases and their potential inhibitors from microbial sources. Microorganisms 2021, 9, 2481. [Google Scholar] [CrossRef]
- Rathnavelu, V.; Alitheen, N.B.; Sohila, S.; Kanagesan, S.; Ramesh, R. Potential role of bromelain in clinical and therapeutic applications (Review). Biomed. Rep. 2016, 5, 283–288. [Google Scholar] [CrossRef]
- Maurer, H.R. Bromelain: Biochemistry, pharmacology, and medical use. Cell. Mol. Life Sci. 2001, 58, 1234–1245. [Google Scholar] [CrossRef]
- Gomes, H.A.R.; Moreira, L.R.S.; Filho, E.X.F. Chapter 3—Enzymes and food industry: A consolidated marriage. In Advances in Biotechnology for Food Industry; Academic Press: Cambridge, MA, USA, 2018; pp. 55–89. [Google Scholar]
- Pavan, R.; Jain, S.; Shraddha; Kumar, A. Properties and therapeutic application of bromelain: A review. Biotechnol. Res. Int. 2012, 2012, 976203. [Google Scholar] [CrossRef]
- Chobotova, K.; Vernallis, A.B.; Majid, F.A.A. Bromelain’s activity and potential as an anti-cancer agent: Current evidence and perspectives. Cancer Lett. 2010, 290, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Castell, J.V.; Friedrich, G.; Kuhn, C.S.; Poppe, G.E. Intestinal absorption of undegraded proteins in men: Presence of bromelain in plasma after oral intake. Am. J. Physiol. Cell Physiol. 1997, 273, G139–G146. [Google Scholar] [CrossRef]
- Ketnawa, S.; Rawdkuen, S. Application of bromelain extract for muscle foods tenderization. Food Nutr. Sci. 2011, 2, 393–401. [Google Scholar] [CrossRef]
- Redwan, R.M.; Saidin, A.; Kumar, S.V. The draft genome of MD-2 pineapple using hybrid error correction of long reads. DNA Res. 2016, 23, 427–439. [Google Scholar] [CrossRef]
- Razali, R.; Kumar, V.; Budiman, C. Structural insights into the enzymatic activity of cysteine protease bromelain of MD2 pineapple. Pak. J. Biol. Sci. 2020, 23, 829–838. [Google Scholar] [CrossRef]
- Bala, M.; Mel, M.; Jami, M.S.; Amid, A.; Salleh, H.M. Kinetic studies on recombinant stem bromelain. Adv. Enzym. Res. 2013, 1, 52–60. [Google Scholar] [CrossRef]
- Muntari, B.; Amid, A.; Mel, M.; Jami, M.S.; Salleh, H.M. Recombinant bromelain production in Escherichia coli: Process optimization in shake flask culture by response surface methodology. AMB Expr. 2012, 2, 12. [Google Scholar] [CrossRef]
- Amid, A.; Ismail, N.A.; Yusof, F.; Salleh, H.M. Expression, purification, and characterization of a recombinant stem bromelain from Ananas comosus. Process Biochem. 2011, 46, 2232–2239. [Google Scholar] [CrossRef]
- Froger, A.; Hall, J.E. Transformation of plasmid DNA into E. coli using the heat shock method. J. Vis. Exp. 2007, 6, 253. [Google Scholar] [CrossRef]
- Kannan, Y.; Koga, Y.; Inoue, Y.; Haruki, M.; Takagi, M.; Imanaka, T.; Morikawa, M.; Kanaya, S. Active subtilisin-like protease from a hyperthermophilic archaeon in a form with a putative prosequence. Appl. Environ. Microbiol. 2001, 67, 2445–2452. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Miyazaki, M. Refolding techniques for recovering biologically active recombinant proteins from inclusion bodies. Biomolecules 2014, 4, 235–251. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, M.; Ali, L.; Islam, W.; Noman, A.; Zhou, C.; Shen, L.; Zhu, T.; Can, L.; Nasif, O.; Gasparovic, K.; et al. Heterologous expression and characterization of glycoside hydrolase with its potential applications in hyperthermic environment. Saudi J. Biol. Sci. 2022, 29, 751–757. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, T.W.; Morton, R.A. The spectrophotometric determination of tyrosine and tryptophan in proteins. Biochem. J. 1956, 40, 628–632. [Google Scholar] [CrossRef] [PubMed]
- LaemmLi, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Silverstein, R.M. The assay of the bromelains using N alpha-CBZ-L-lysine p-nitrophenyl ester and N-CBZ-glycine p-nitrophenyl ester as substrates. Anal. Biochem. 1975, 62, 478–484. [Google Scholar] [CrossRef]
- Nelson, D.L.; Cox, M.M. Lehnigher Principles of Biochemistry, 5th ed.; Freeman and Company: New York, NY, USA, 2008. [Google Scholar]
- Hidayani, W.A.; Setiasih, S.; Hudiyono, S. Determination of the effect of EDTA and PCMB on purified bromelain activity from pineapple core and in vitro antiplatelet activity. IOP Conf. Ser. Mater. Sci. Eng. 2020, 763, 012054. [Google Scholar] [CrossRef]
- Singh, A.N.; Shukla, A.K.; Jagannadham, M.V.; Dubey, V.K. Purification of a novel cysteine protease, procerain B, from Calotropis procera with distinct characteristics compared to Procerain. Process Biochem. 2010, 45, 399–406. [Google Scholar] [CrossRef]
- Lestari, P.; Suyata. Antibacterial activity of hydrolysate protein from Etawa goat milk hydrolysed by crude extract bromelain. IOP Conf. Ser. Mater. Sci. Eng. 2019, 509, 012111. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Nussinov, R. Eukaryotic dinucleotide preference rules and their implications for degenerate codon usage. J. Mol. Biol. 1981, 149, 125–131. [Google Scholar] [CrossRef]
- Aktar, H.; Aktar, S.; Jan, S.U.; Khan, A.; Zaidi, N.S.S.; Qadri, I. Over expression of a synthetic gene encoding interferon lambda using relative synonymous codon usage bias in Escherichia coli. Pak. J. Pharm Sci. 2013, 26, 1181–1188. [Google Scholar]
- Souza, P.M.; Aliakbarian, B.; Filho, E.X.F.; Magalhães, P.O.; Junior, A.P.; Converti, A.; Perego, P. Kinetic and thermodynamic studies of a novel acid protease from Aspergillus foetidus. Macromolecules 2015, 81, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Dżugan, M.; Tomczyk, M.; Sowa, P.; Grabek-Lejko, D. Antioxidant activity as biomarker of honey variety. Molecules 2018, 23, 2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menzella, H.G. Comparison of two codon optimization strategies to enhance recombinant protein production in Escherichia coli. Microb. Cell Fac. 2011, 10, 15. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Mei, C.; Zhen, H.; Zhu, J. Codon preference optimization increases prokaryotic Cystatin C expression. J. Biomed. Biotechnol. 2012, 2012, 732017. [Google Scholar] [CrossRef]
- Goh, C.G.K.; Silvester, J.; Mahadi, W.N.S.W.; Chin, L.P.; Ying, L.T.; Leow, T.C.; Kurahashi, R.; Takano, K.; Budiman, C. Expression and characterization of functional domains of FK506-binding protein 35 from Plasmodium knowlesi. Protein Eng. Des. Sel. 2018, 31, 489–498. [Google Scholar] [CrossRef]
- Gustafsson, C.; Govindarajan, S.; Minshull, J. Codon bias and heterologous protein expression. Trends Biotechnol. 2004, 22, 346–353. [Google Scholar] [CrossRef]
- Bhatwa, A.; Wang, W.; Hassan, Y.I.; Abraham, N.; Li, X.-Z.; Zhou, T. Challenges associated with the formation of recombinant protein inclusion bodies in Escherichia coli and strategies to address them for industrial applications. Front. Bioeng. Biotechnol. 2021, 9, 630551. [Google Scholar] [CrossRef]
- Zhou, Z.; Dang, Y.; Zhou, M.; Li, L.; Yu, C.-H.; Fu, J.; Chen, S.; Liu, Y. Codon usage is an important determinant of gene expression levels largely through its effects on transcription. Proc. Natl. Acad. Sci. USA 2016, 113, E6117–E6125. [Google Scholar] [CrossRef]
- Obeng, E.M.; Brossette, T.; Ongkudon, C.M.; Budiman, C.; Maas, R.; Jose, J. The workability of Escherichia coli BL21 (DE3) and Pseudomonas putida KT2440 expression platforms with autodisplayed cellulases: A comparison. Appl. Microbiol. Biotechnol. 2018, 102, 4829–4841. [Google Scholar] [CrossRef]
- Razali, R.; Budiman, C.; Kumar, V. Technical data of heterologous expression and purification of SARS-CoV-2 proteases using Escherichia coli system. Data 2021, 6, 99. [Google Scholar] [CrossRef]
- Kane, J.F.; Hartley, D.L. Formation of recombinant protein inclusion bodies in Escherichia coli. Trends Biotechnol. 1988, 6, 95–101. [Google Scholar] [CrossRef]
- Fahnert, B.; Lile, H.; Neubauer, P. Inclusion bodies: Formation and utilization. Adv. Biochem. Eng. Biotechnol. 2004, 89, 93–142. [Google Scholar] [PubMed]
- Sørensen, H.P.; Mortensen, K.K. Soluble expression of recombinant proteins in the cytoplasm of Escherichia coli. Microb. Cell Fact. 2005, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Arshad, Z.I.M.; Amid, A.; Yusof, F.; Sulaiman, S.Z.; Mudalip, S.K.A.; Man, R.C.; Shaarani, S.M. Comparison of purification methods to purify recombinant bromelain from Escherichia coli BL21-A1. Malays. J. Anim. Sci. 2017, 21, 958–971. [Google Scholar]
- George, S.; Bhasker, S.; Madhav, H.; Nair, A.; Chinnamma, M. Functional characterization of recombinant bromelain of Ananas comosus expressed in a prokaryotic system. Mol. Biotechnol. 2014, 56, 166–174. [Google Scholar] [CrossRef]
- Singh, A.; Upadhyay, V.; Upadhyay, A.K.; Singh, S.M.; Panda, A.K. Protein recovery from inclusion bodies of Escherichia coli using mild solubilization process. Microb. Cell Fact 2015, 14, 41. [Google Scholar] [CrossRef]
- Costa, S.; Almeida, A.; Castro, A.; Domingues, L. Fusion tags for protein solubility, purification, and immunogenicity in Escherichia coli: The novel Fh8 system. Front. Microbiol. 2014, 5, 63. [Google Scholar] [CrossRef]
- Boisselier, E.; Audet, M.L.; Cantin, L.; Salesse, C. A strategy for purifying glutathione S-transferase in the presence of sodium dodecyl sulfate. BioTechniques 2018, 51, 193–194. [Google Scholar] [CrossRef]
- Young, C.L.; Britton, Z.T.; Robinson, A.S. Recombinant protein expression and purification: A comprehensive review of affinity tags and microbial applications. Biotechnol. J. 2012, 7, 620–634. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, L.; Zhang, Y.; Feng, Y.; Lu, X.; Lan, W.; Wang, J.; Wu, H.; Cao, C.; Wang, X. Highly efficient production of soluble proteins from insoluble inclusion bodies by a two-step-denaturing and refolding method. PLoS ONE 2011, 6, e22981. [Google Scholar] [CrossRef] [PubMed]
- Clark, E.D.B. Protein refolding for industrial processes. Curr. Opin. Biotechnol. 2001, 12, 202–207. [Google Scholar] [CrossRef]
- Lilie, H.; Schwarz, E.; Rudolph, R. Advances in refolding of proteins produced in E. coli. Curr. Opin. Biotechnol. 1998, 9, 497–501. [Google Scholar] [CrossRef]
- Bala, M.; Salleh, H.M.; Amid, A.; Mel, M.; Jami, M.S. Recovery of recombinant bromelain from Escherichia coli BL21-A1. Afr. J. Biotechnol. 2011, 10, 18829–18832. [Google Scholar] [CrossRef]
- Robichon, C.; Luo, J.; Casuey, T.B.; Benner, J.S.; Samuelson, J.C. Engineering Escherichia coli BL21(DE3) derivative strains to minimize E. coli protein contamination after purification by immobilized metal affinity chromatography. Appl. Environ. Microbiol. 2011, 7, 4634–4646. [Google Scholar] [CrossRef]
- Bala, M.; Ismail, N.A.; Mel, M.; Jami, M.S.; Salleh, H.M.; Amid, A. Bromelain production: Current trends and perspective. Arch. Des. Sci. 2012, 65, 360–399. [Google Scholar]
- Corzo, C.A.; Waliszewski, K.N.; Welti-Chanes, J. Pineapple fruit bromelain affinity to different protein substrates. Food Chem. 2012, 133, 631–635. [Google Scholar] [CrossRef]
- Bartholomew, D.P.; Paull, R.E.; Rohrbach, K.G. The Pineapple: Botany, Production and Uses; CABI Publishing: Wallingford, UK, 2003; pp. 1–301. [Google Scholar]
- Harrach, T.; Eckert, K.; Maurer, H.R.; Machleidt, I.; Machleidt, W.; Nuck, R. Isolation and characterization of two forms of an acidic bromelain stem proteinase. J. Prot. Chem. 1998, 17, 351–361. [Google Scholar] [CrossRef]
- Suh, H.J.; Yang, H.C.; Lee, H.; Suwon; Cho, H.Y. Purification and characterization of bromelain isolated from pineapple. J. Korean Agric. Chem. Soc. 1992, 35, 300–307. [Google Scholar]
- Ketnawa, S.; Rawdkuen, S.; Chaiwut, P. Two phase partitioning and collagen hydrolysis of bromelain from pineapple peel Nang Lae cultivar. J. Biochem. Eng. 2010, 52, 205–211. [Google Scholar] [CrossRef]
- Silvestre, M.P.C.; Carreira, R.L.; Silva, M.R.; Corgosinho, F.C.; Monteiro, M.R.P.; Moais, H.A. Effect of pH and temperature on the activity of enzymatic extracts from pineapple peel. Food Bioproc. Technol. 2012, 5, 1824–1831. [Google Scholar] [CrossRef]
- Grzonka, Z.; Kasprzykowski, F.; Wiczk, W. Cysteine Proteases. In Industrial Enzymes: Structure, Function and Applications; Springer: Dordrecht, The Netherlands, 2007; pp. 181–195. [Google Scholar]
- Ketnawa, S.; Chaiwut, P.; Rawdkuen, S. Extraction of bromelain from pineapple peels. Food Sci. Technol. Int. 2011, 4, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Vernet, T.; Berti, P.J.; de Montigny, C.; Musil, R.; Tessier, D.C.; Ménard, R.; Magny, M.C.; Storer, A.C.; Thomas, D.Y. Processing of the papain precursor. The ionization state of a conserved amino acid motif within the Pro region participates in the regulation of intramolecular processing. J. Biol. Chem. 1995, 270, 10838–10846. [Google Scholar] [CrossRef]
- Verma, S.; Dixit, R.; Pandey, K.C. Cysteine proteases: Modes of activation and future prospects as pharmacological targets. Front. Pharmacol. 2016, 7, 107. [Google Scholar] [CrossRef]
- Fadhilah, Y.; Shoobihah, A.; Setiasih, S.; Handayani, S.; Hudiyono, S. The effect of Ca2+, Mg2+ ions, cysteine, and benzoic acid on the activity of purified bromelain from pineapple core extract (Ananas comosus [L]. Merr). AIP Conf. Proc. 2018, 2049, 020029-1–020029-5. [Google Scholar]
- Liang, H.Y.; Li, M.; Shi, M.; Liao, A.P.; Wu, R.C. Study on the stability of fruit bromelain. Adv. Mater. Res. 2011, 421, 19–22. [Google Scholar]
- Haq, S.K.; Rasheedi, S.; Sharma, P.; Ahmad, B.; Khan, R.H. Influence of salts and alcohols on the conformation of partially folded intermediate of stem bromelain at low pH. Int. J. Biochem. Cell Biol. 2005, 37, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Shukor, M.Y.; Masdor, N.; Baharom, N.A.; Jamal, J.A.; Abdullah, M.P.A.; Shamaan, N.A.; Syed, M.A. An inhibitive determination method for heavy metals using bromelain, a cysteine protease. Appl. Biochem. Biotechnol. 2008, 144, 283–291. [Google Scholar] [CrossRef]
- Kaul, P.; Sathish, H.A.; Prakash, V. Effect of metal ions on structure and activity of papain from Carica papaya. Nahrung 2002, 46, 2–6. [Google Scholar] [CrossRef]
- Kaur, T.; Kaur, A.; Grewal, R.K. Kinetics studies with fruit bromelain (Ananas comosus) in the presence of cysteine and divalent ions. J. Food Sci. Technol. 2015, 52, 5954–5960. [Google Scholar] [CrossRef]
- Wang, X.; Liu, Z.; Hu, X.; Huanh, H. Effects of Ca~(2+) on thermo stability and secondary structure of bromelain. Chin. Food Addit. 2009, 30, 153–155. [Google Scholar]
- Masdor, N.A.; Said, N.A.M. Partial purification of crude stem bromelain improves its sensitivity as a protease inhibitive assay for heavy metals. Aust. J. Basic Appl. Sci. 2011, 5, 1295–1298. [Google Scholar]
- Marshall, S.; Golden, J. Characterization of bromelain from Morinda citrifolia (Noni). J. Sci. Res. 2012, 4, 445–456. [Google Scholar]
- Saptarini, N.M.; Rahayu, D.; Herawati, I.E. Antioxidant activity of crude bromelain of pineapple (Ananas comosus (L.) Merr) Crown from Subang District, Indonesia. J. Pharm. Bioallied Sci. 2019, 11, S551–S555. [Google Scholar] [CrossRef]
- Huang, C.W.; Lin, I.J.; Liu, Y.M.; Mau, J.L. Composition, enzyme and antioxidant activities of pineapple. Int. J. Food. Prop. 2021, 23, 1244–1251. [Google Scholar] [CrossRef]
- Abbas, S.; Shanbhag, T.; Kothare, A. Applications of bromelain from pineapple waste towards acne. Saudi. J. Biol. Sci. 2021, 28, 1001–2009. [Google Scholar]
- Ataide, J.A.; de Carvalho, N.M.; Rebelo, M.d.; Chaud, M.V.; Grotto, D.; Gerenutti, M.; Rai, M.; Mazzola, P.G.; Jozala, A.F. Bacterial nanocellulose loaded with bromelain: Assessment of antimicrobial, antioxidant and physical-chemical properties. Sci. Rep. 2017, 7, 18031. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Aluko, R.E. Chemometric analysis of the amino acid requirements of antioxidant food protein hydrolysates. Int. J. Mol. Sci. 2011, 12, 3148–3161. [Google Scholar] [CrossRef]
- Chakraborty, A.J.; Mitra, S.; Tallei, T.E.; Tareq, A.M.; Nainu, F.; Cicia, D.; Dhama, K.; Emran, T.B.; Simal-Gandara, J.; Capasso, R. Bromelain a potential bioactive compound: A comprehensive overview from a pharmacological perspective. Life 2021, 11, 317. [Google Scholar] [CrossRef]
- Lasekan, O.; Hussein, F.K. Classification of different pineapple varieties grown in Malaysia based on volatile fingerprinting and sensory analysis. Chem. Cent. J. 2018, 12, 140. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Original MD2-SBro Gene | Optimized MD2-SBro Gene |
|---|---|---|
| GC content | 44.51% | 53.34% |
| Codon adaptation index (CAI) | 0.40 | 0.95 |
| Steps | Volume (mL) | Total Protein (mg) | Total Activity (U) * | Specific Activity (U/mg) * | Yield (%) | Purification (Fold) |
|---|---|---|---|---|---|---|
| Cell lysate | 70 ± 2.80 | 510 ± 12 | 62.30 ± 5.64 | 0.12 ± 0.002 | 100 | 1.0 |
| Glutathione S-transferase (GST) affinity chromatography | 22 ± 1.38 | 14 ± 1.32 | 49.84 ± 3.17 | 3.56 ± 0.08 | 80 | 42.72 |
| Proteins | Vmax (10−3 µM s−1) | Km (µM) | kcat (s−1) | kcat/Km (10−3 µM−1 s−1) | Ref |
|---|---|---|---|---|---|
| MD2-SBro | 7.20 ± 0.52 | 42.1 ± 3.81 | 0.20 ± 0.008 | 4.75 ± 0.23 | This study |
| MD2-MBro | 15 ± 0.5 | 34.24 ± 1.02 | 1.93 ± 0.05 | 56.37 ± 2.08 | [1] |
| Metal Ions | Relative Activity (%) |
|---|---|
| Control | 100 ± 5.01 |
| MgCl2 | 178.32 ± 7.54 |
| CaCl2 | 121.40 ± 8.71 |
| NiCl2 | 114.31 ± 10.32 |
| CuCl2 | 78.09 ± 3.21 |
| ZnCl2 | 47.36 ± 3.98 |
| EDTA | 17.58 ± 1.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Razali, R.; Fahrudin, F.A.; Subbiah, V.K.; Takano, K.; Budiman, C. Heterologous Expression and Catalytic Properties of Codon-Optimized Small-Sized Bromelain from MD2 Pineapple. Molecules 2022, 27, 6031. https://doi.org/10.3390/molecules27186031
Razali R, Fahrudin FA, Subbiah VK, Takano K, Budiman C. Heterologous Expression and Catalytic Properties of Codon-Optimized Small-Sized Bromelain from MD2 Pineapple. Molecules. 2022; 27(18):6031. https://doi.org/10.3390/molecules27186031
Chicago/Turabian StyleRazali, Rafida, Fikran Aranda Fahrudin, Vijay Kumar Subbiah, Kazufumi Takano, and Cahyo Budiman. 2022. "Heterologous Expression and Catalytic Properties of Codon-Optimized Small-Sized Bromelain from MD2 Pineapple" Molecules 27, no. 18: 6031. https://doi.org/10.3390/molecules27186031