
Mechanism of Antifungal Action of Monoterpene Isoespintanol against Clinical Isolates of Candida tropicalis

Abstract
:1. Introduction
2. Results
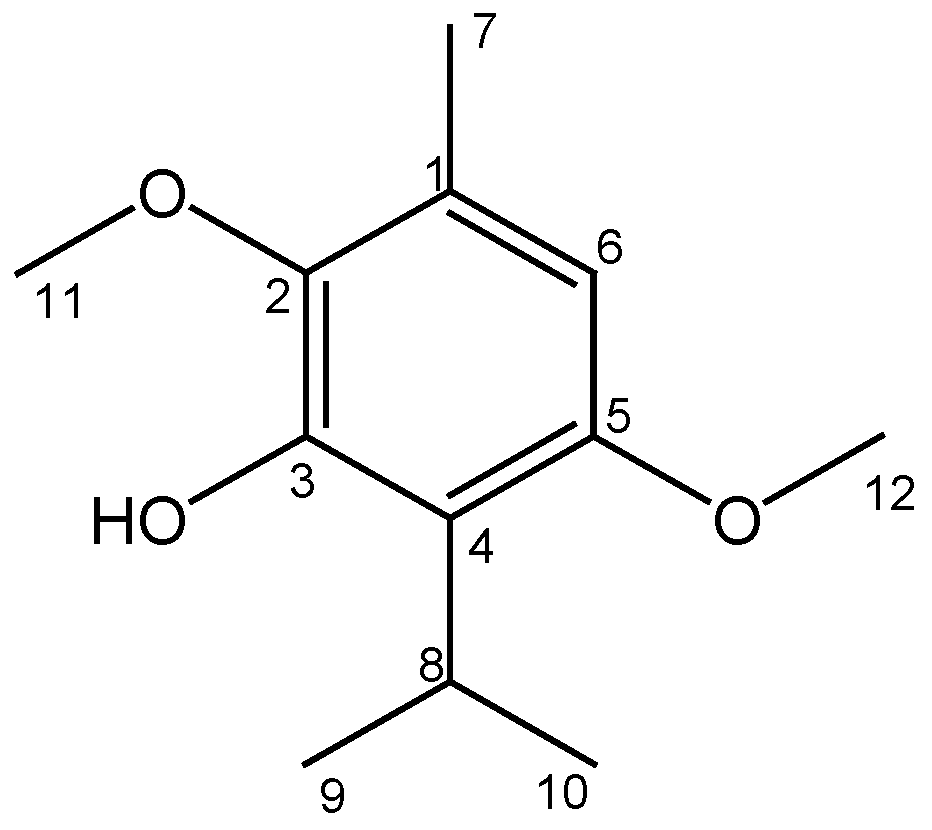
2.1. Obtaining and Identifying Isoespintanol
2.2. Phylogenomics of Candida tropicalis
2.3. Antifungal Susceptibility Testing
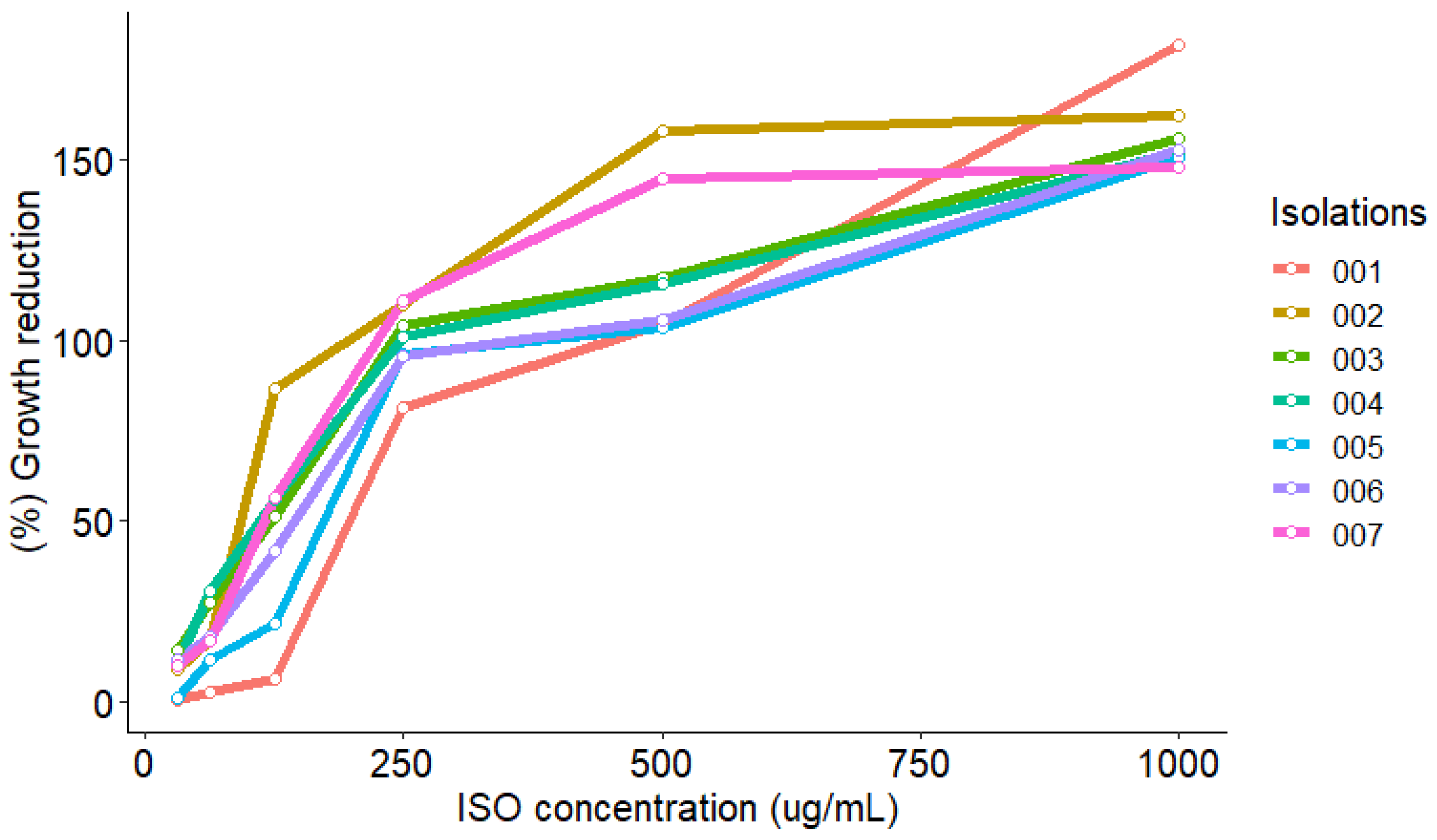
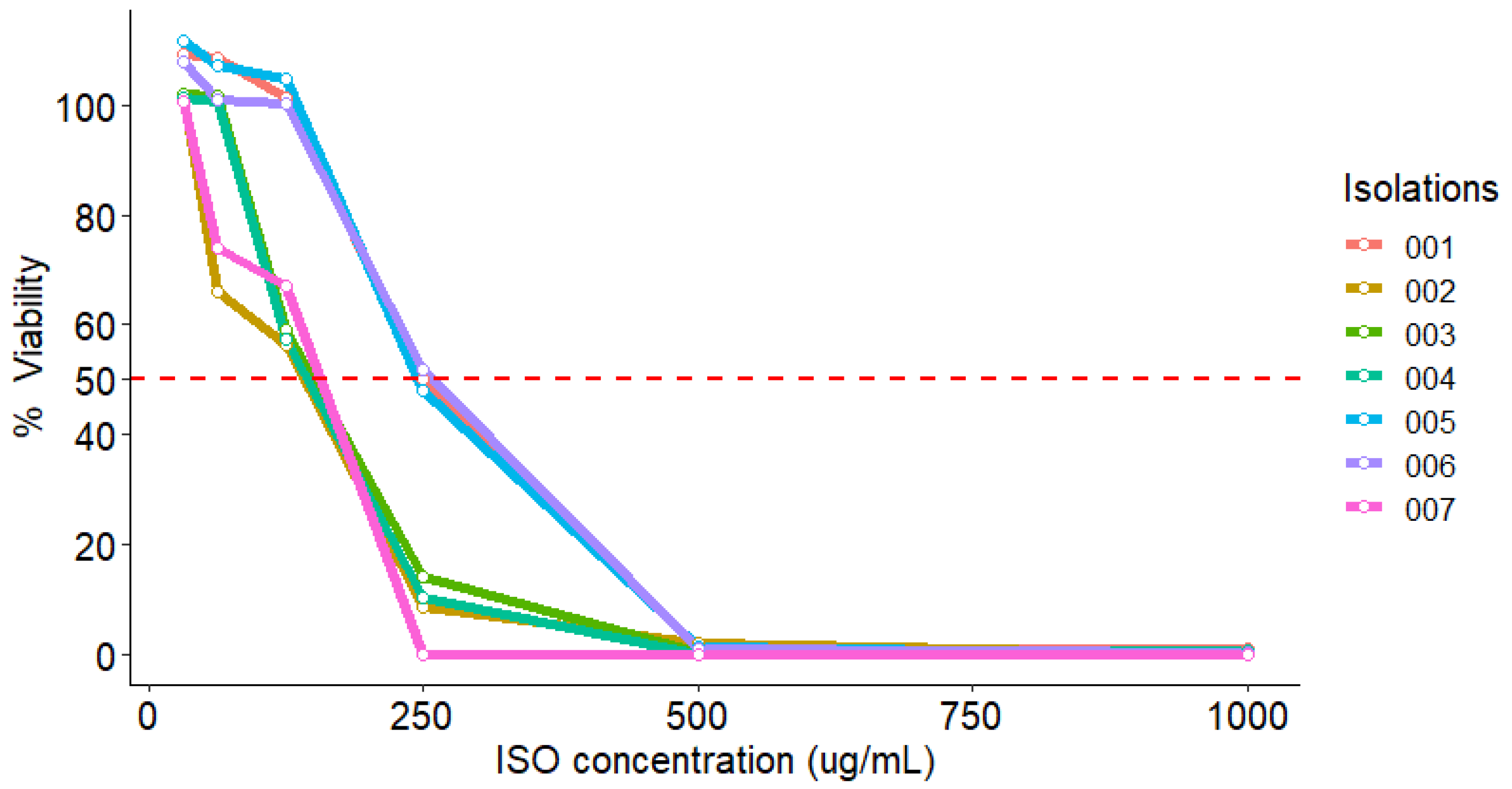
2.4. MTT Reduction Assay
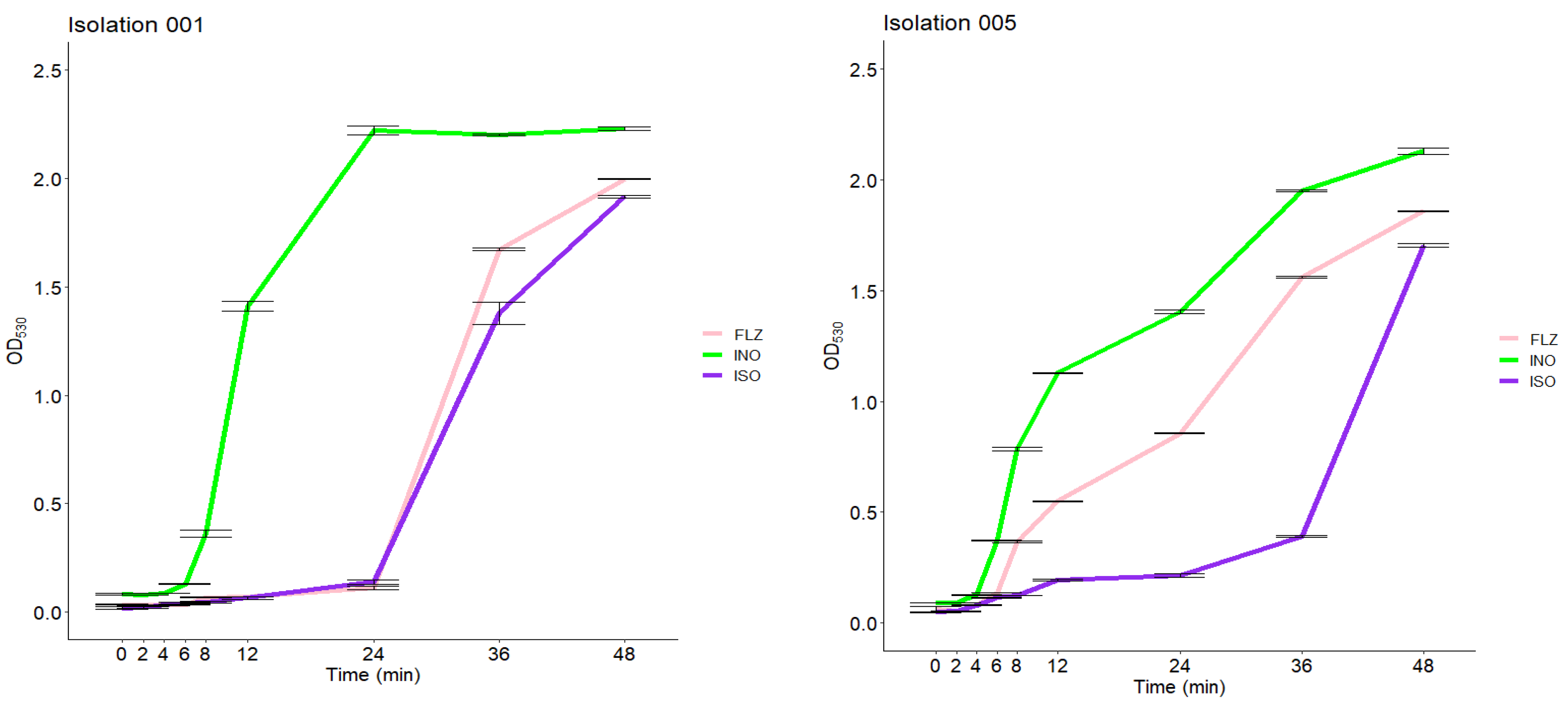
2.5. Growth Inhibition Curve
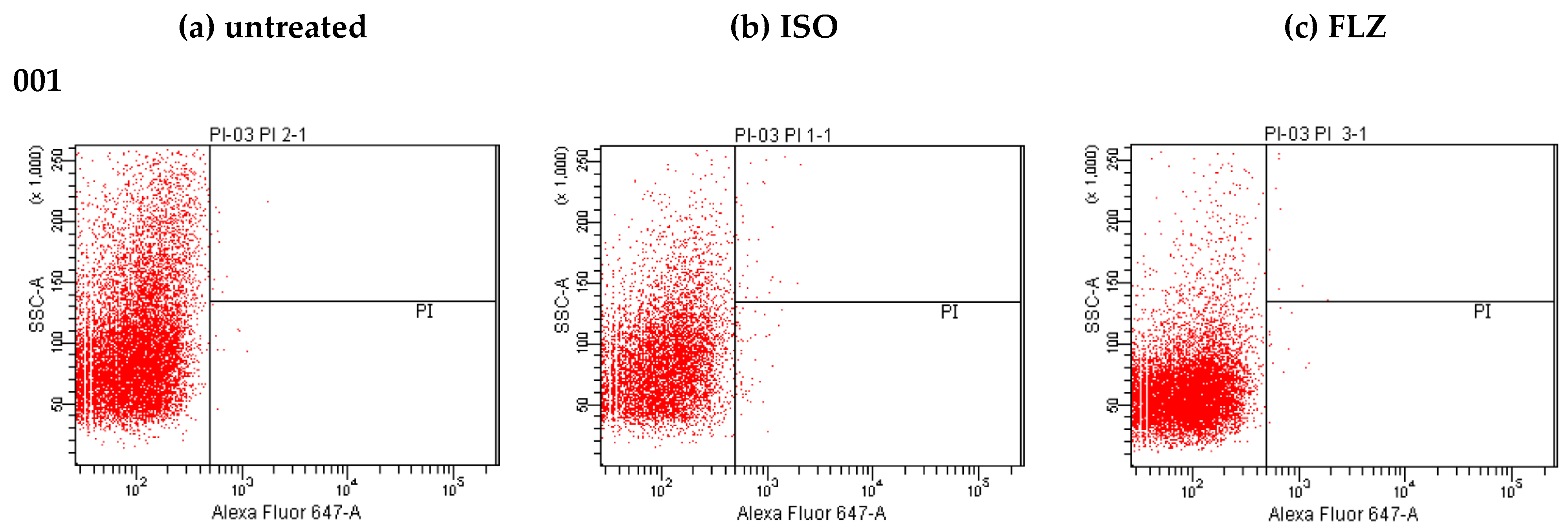
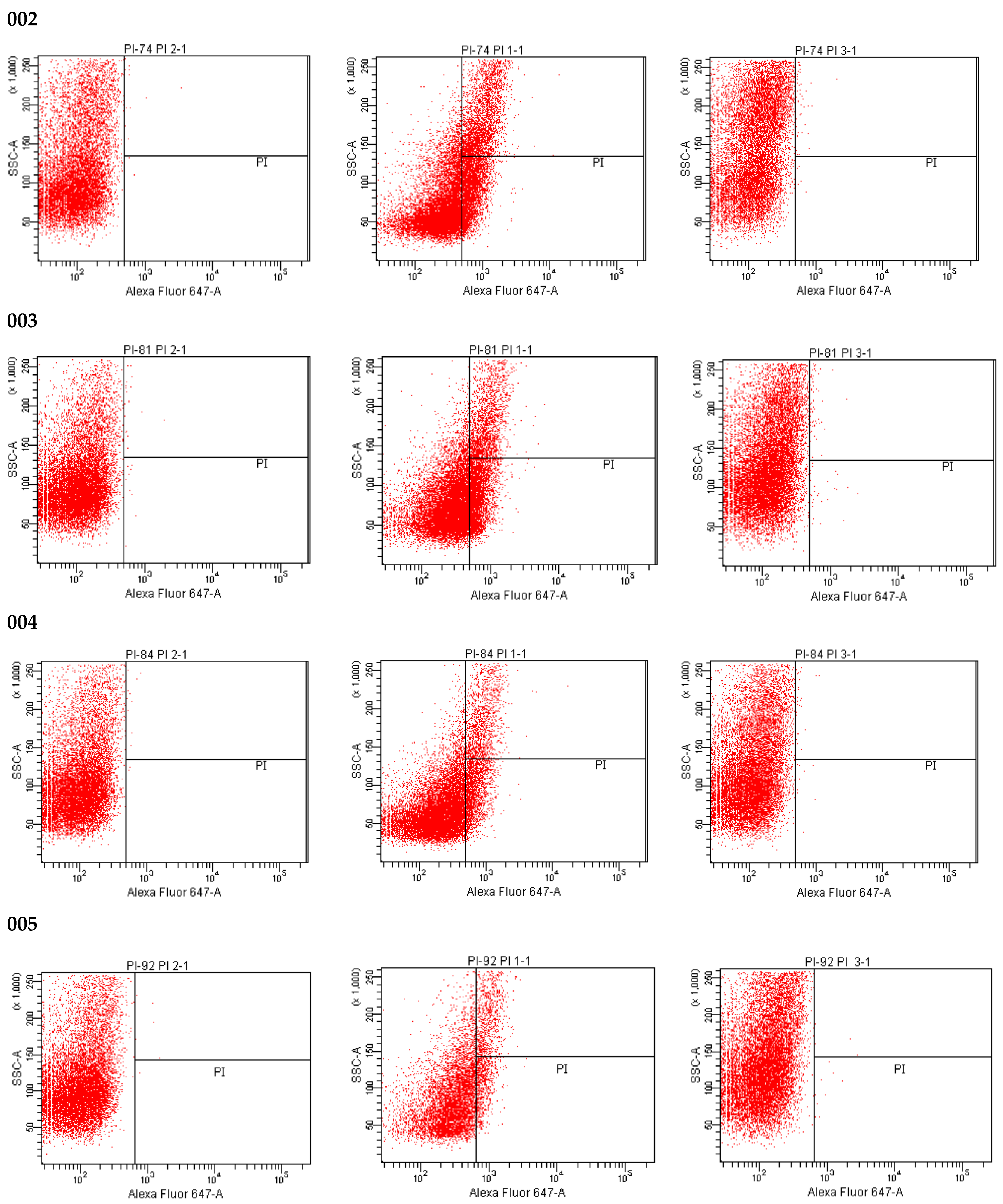
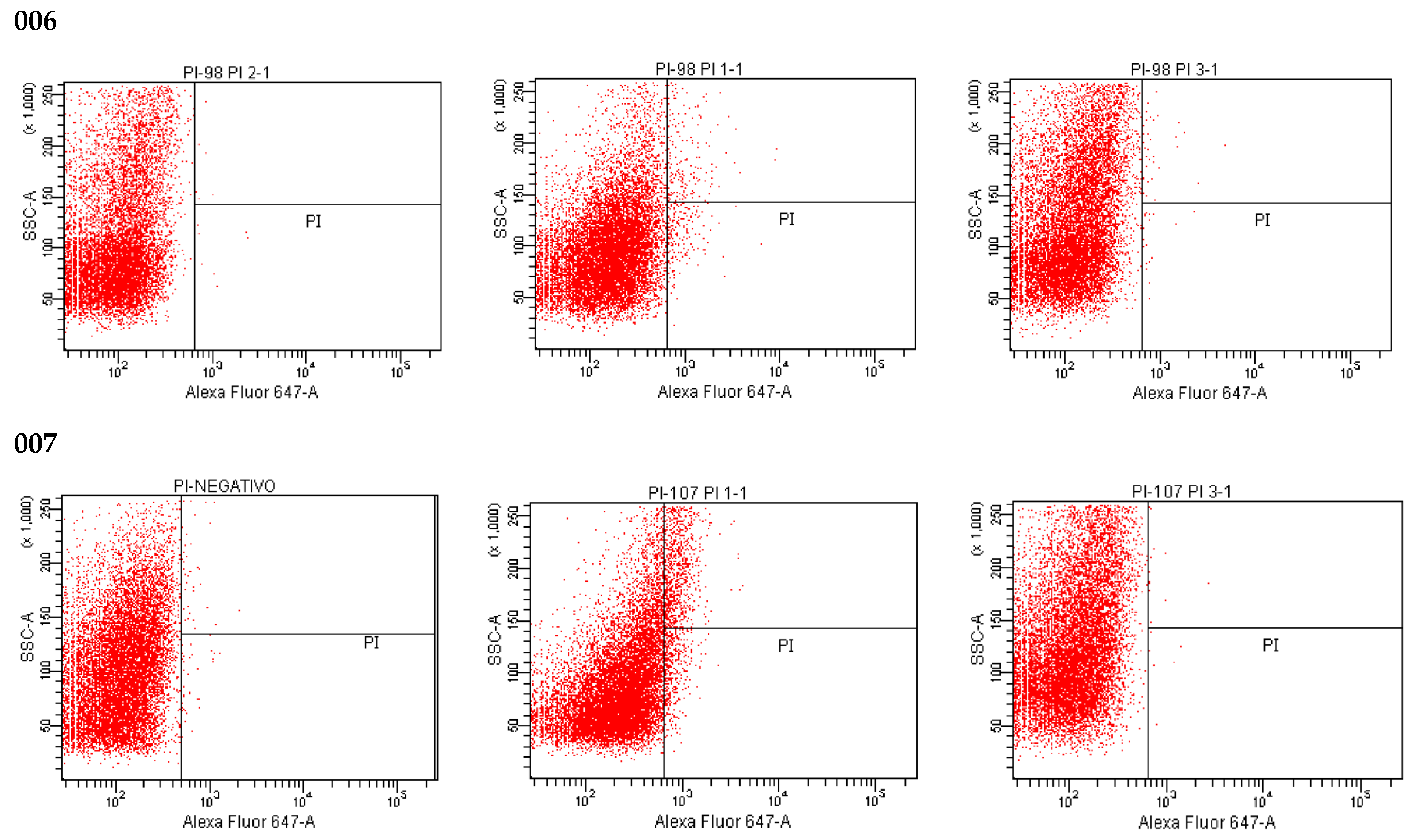
2.6. LIVE/DEAD Assay
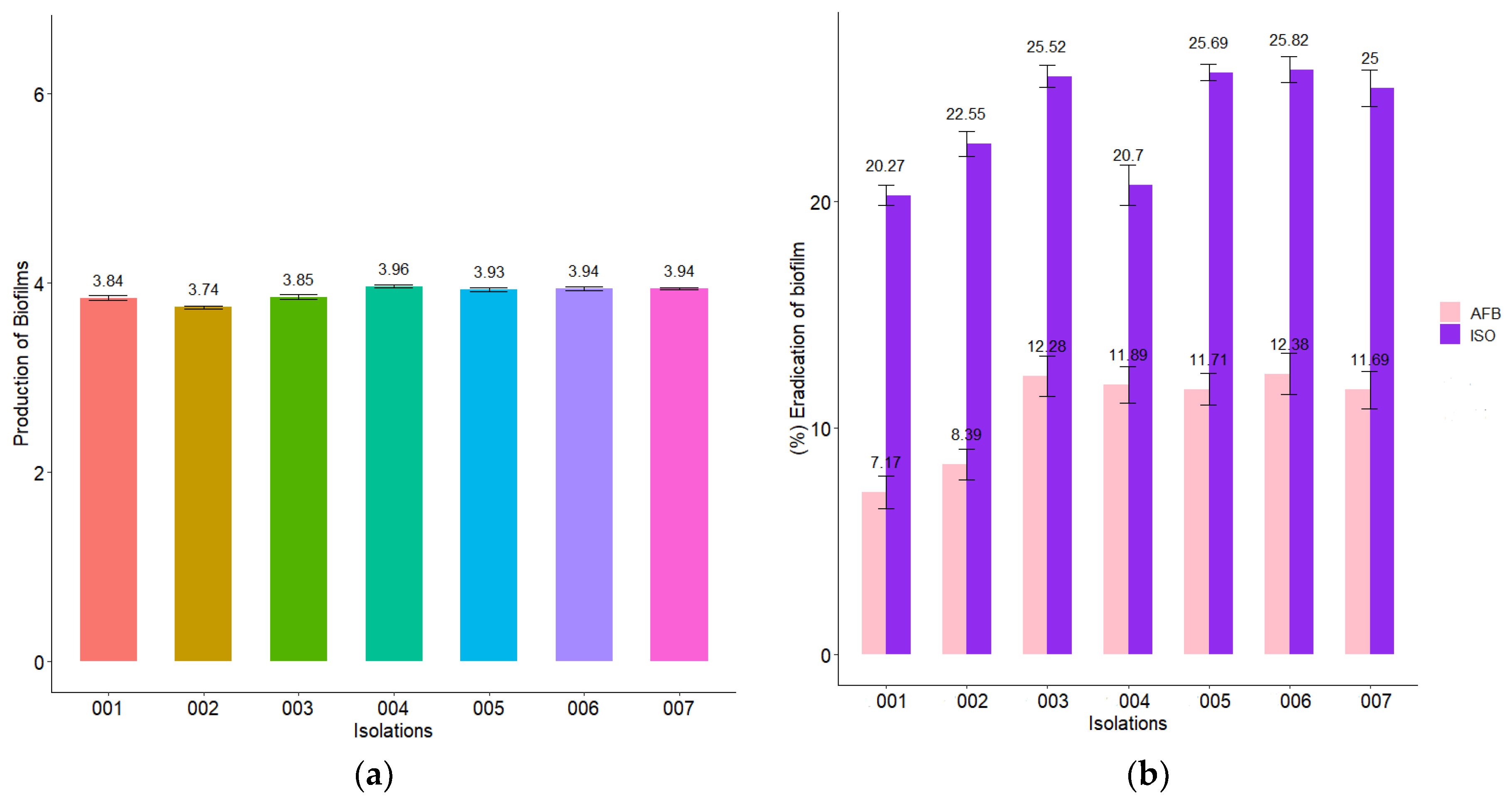
2.7. Biofilm Reduction
2.8. Effect of ISO on Cell Membrane Integrity
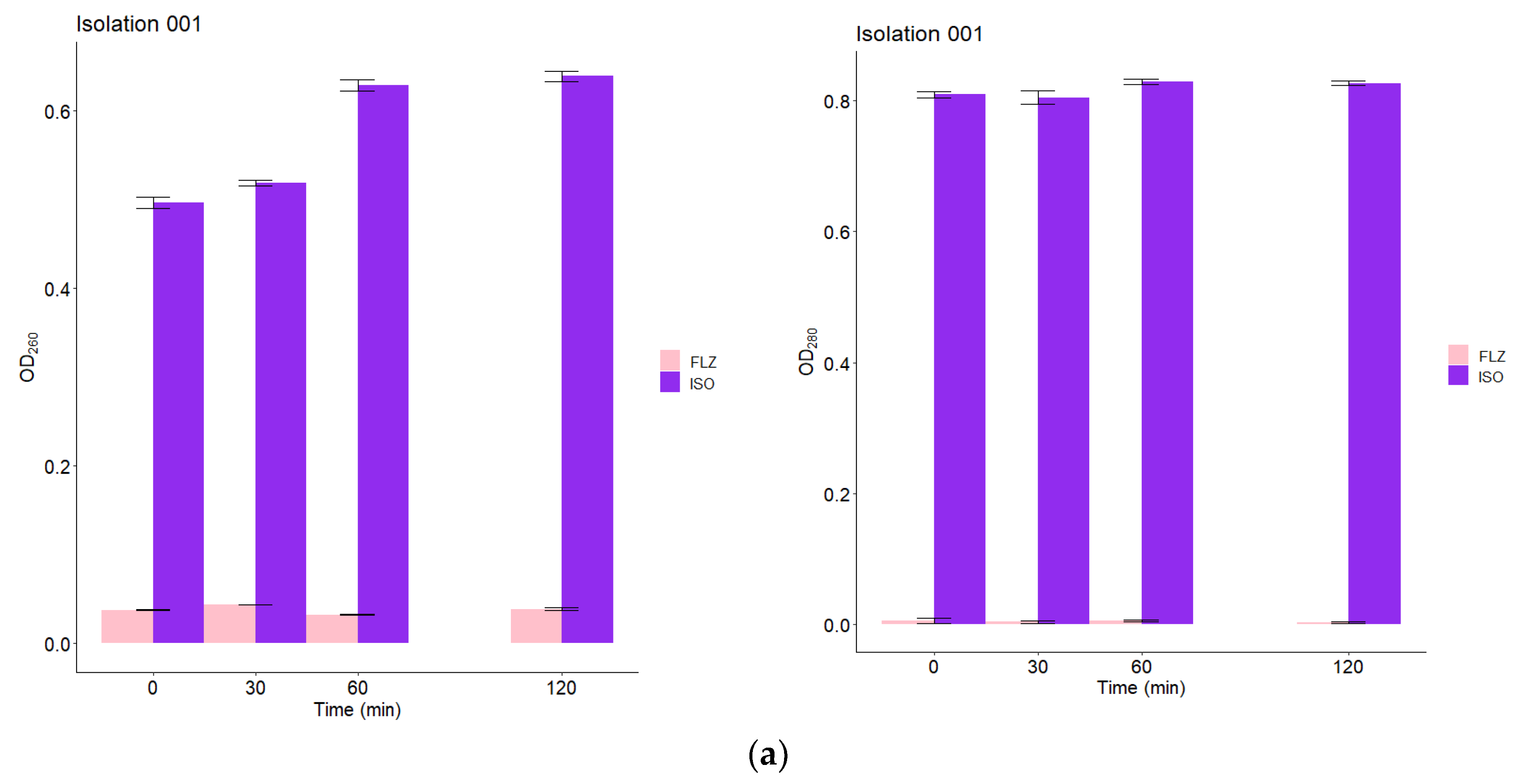
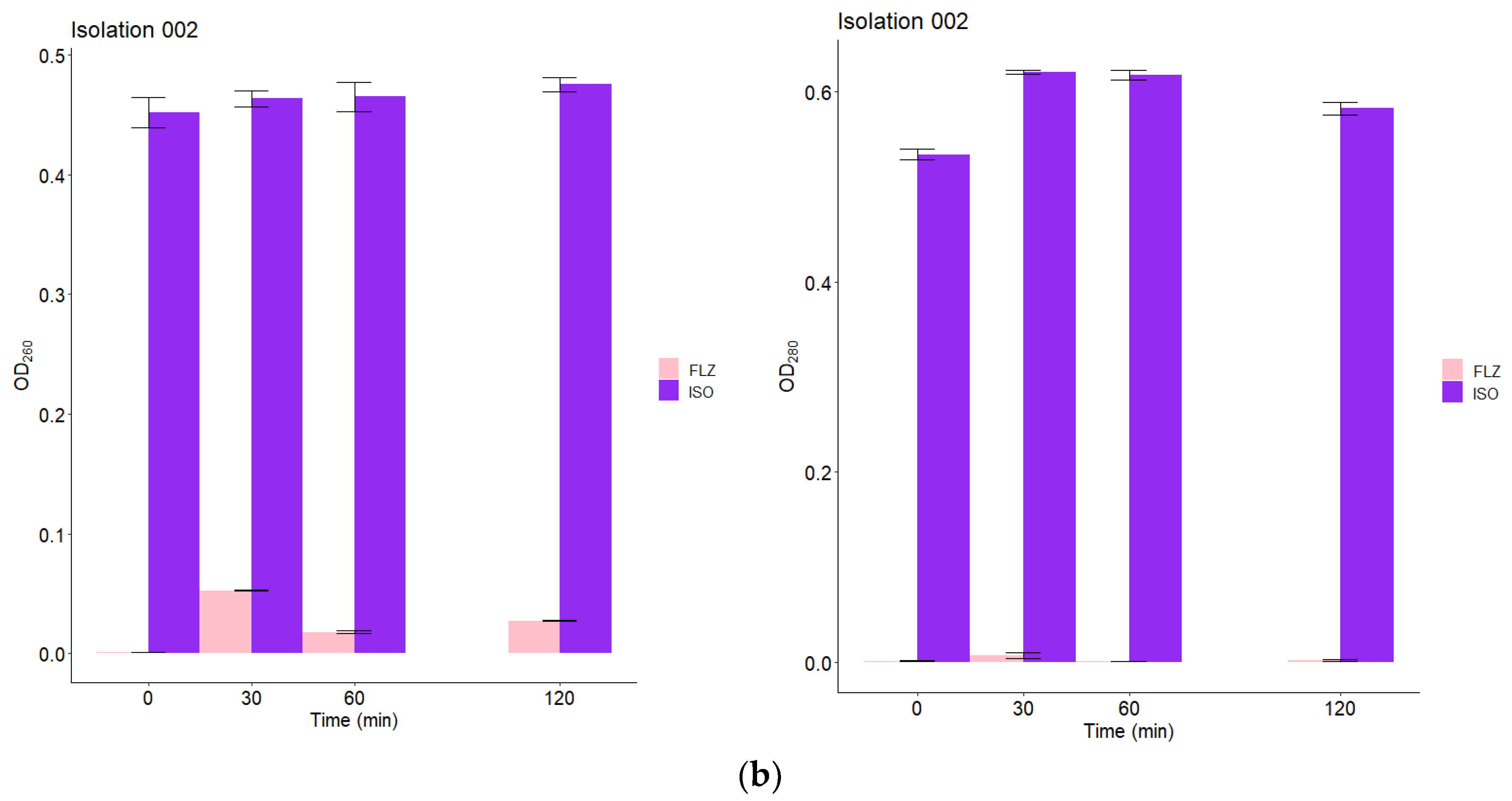
2.9. Leakage of Nucleic Acids and Proteins through the Fungal Membrane
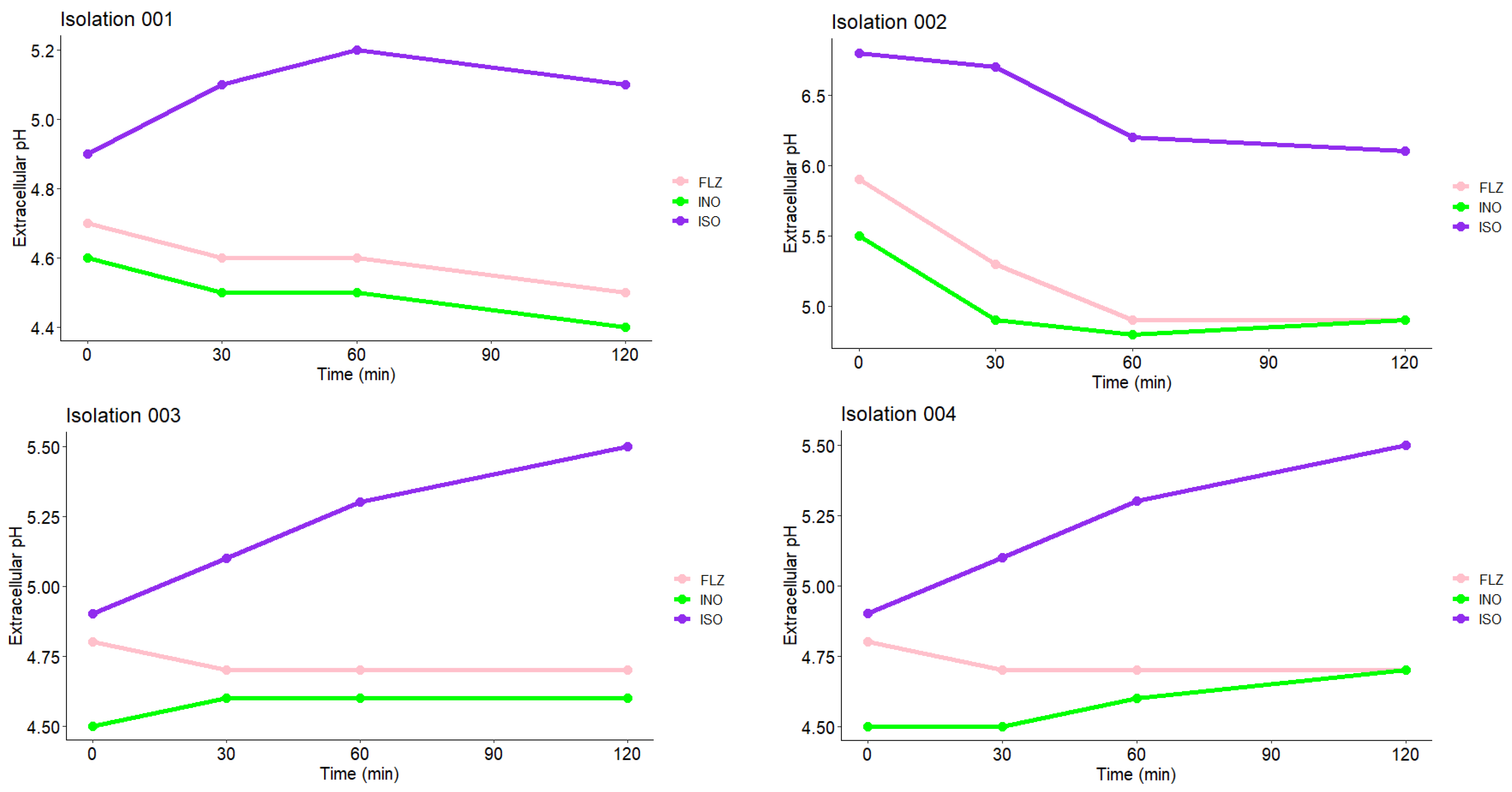
2.10. Measurement of Extracellular pH
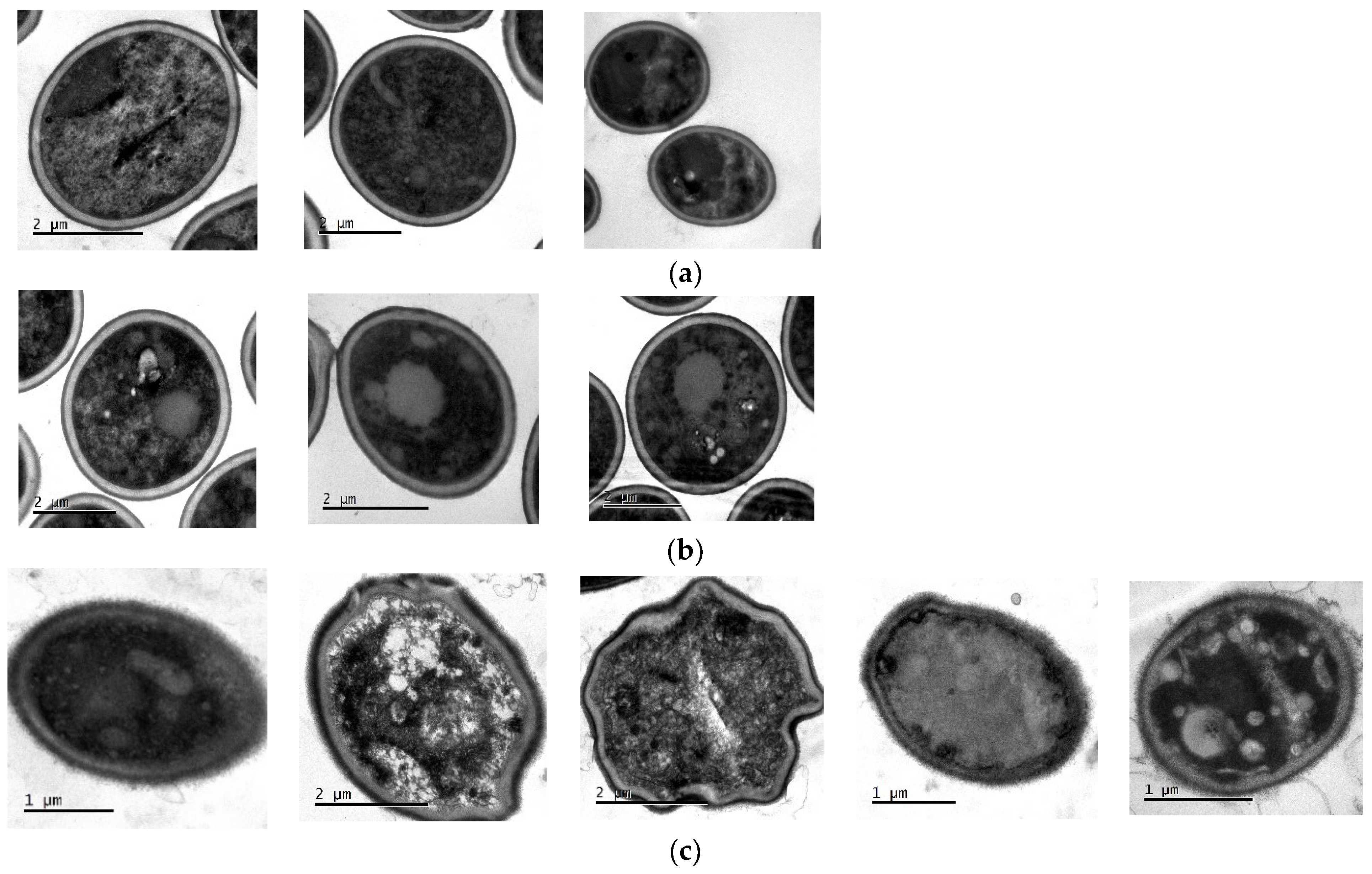
2.11. Transmission Electron Microscopy (TEM)
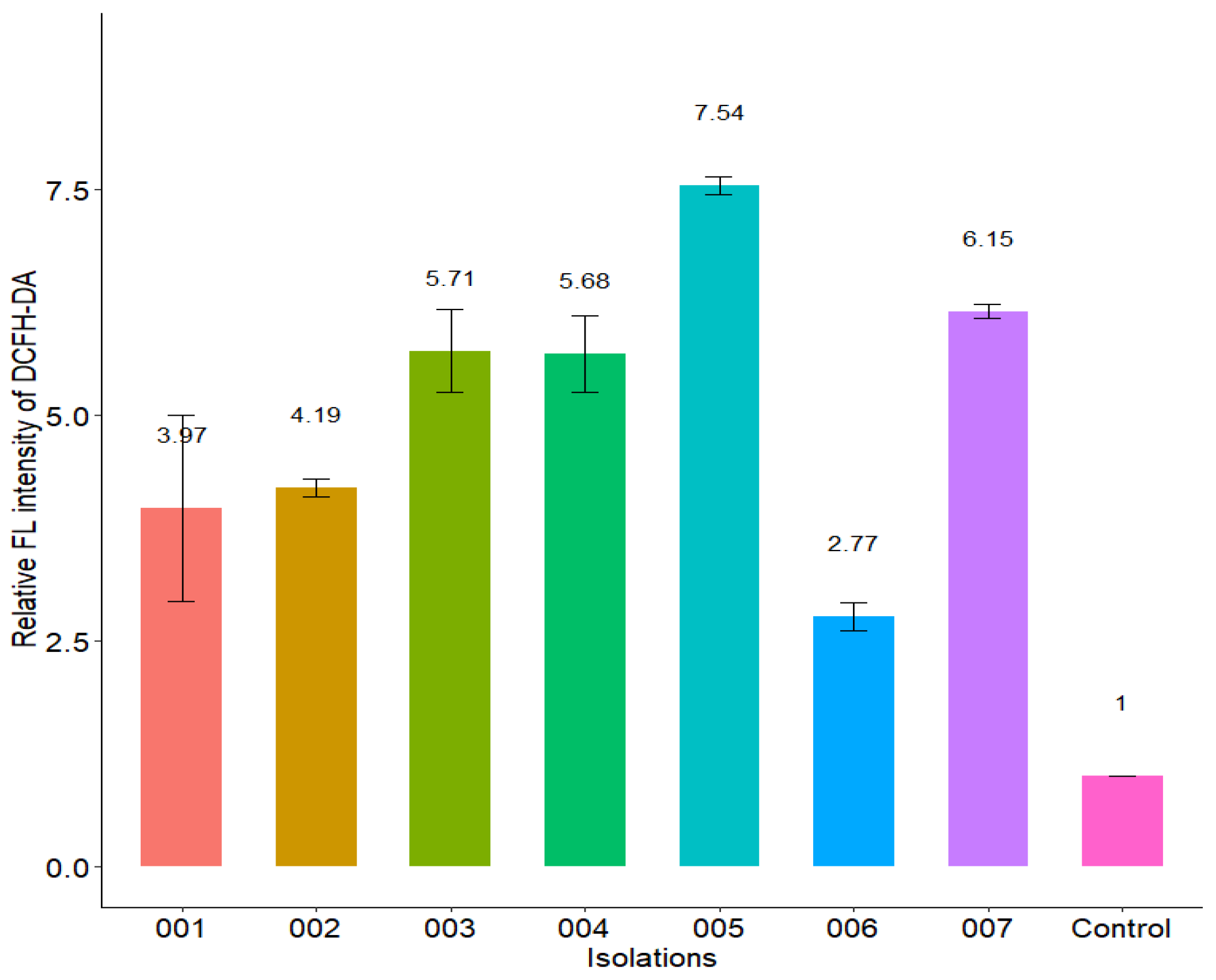
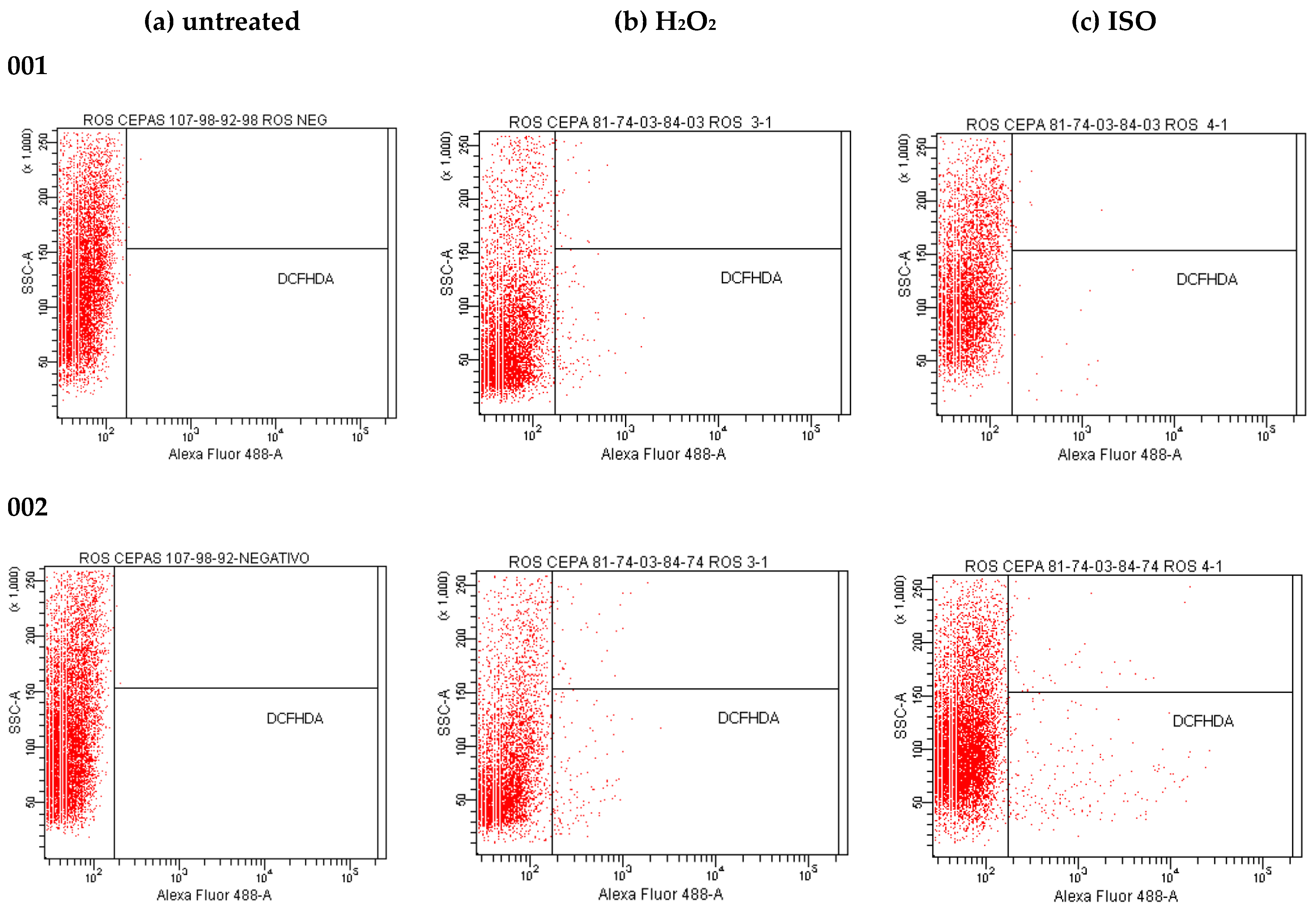
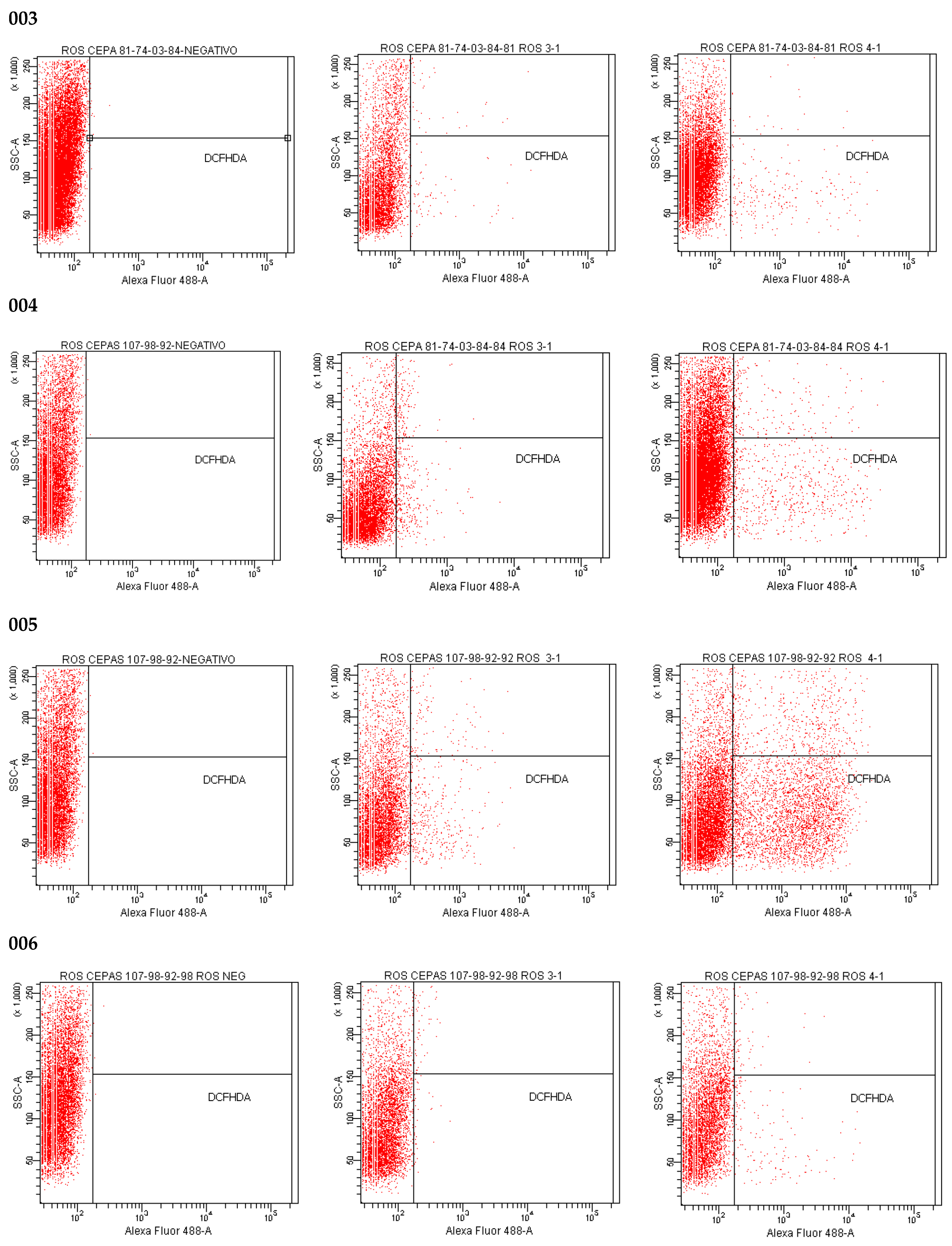
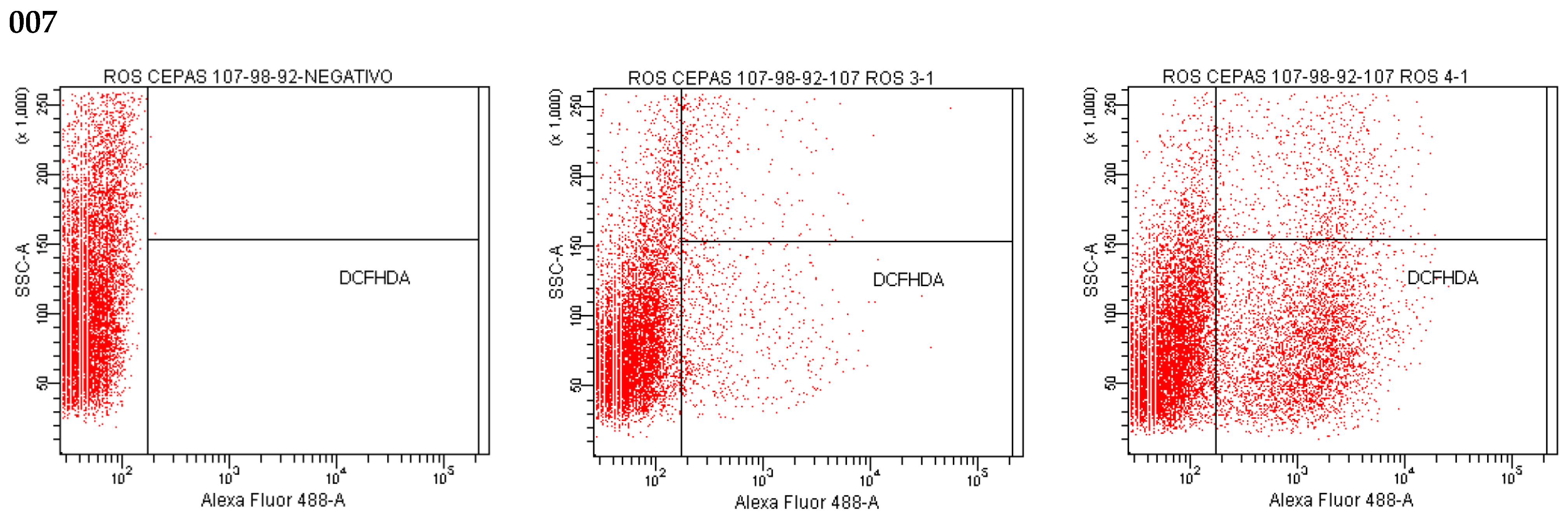
2.12. Effect of Isoespintanol on the Production of Intracellular Reactive Oxygen Species (iROS)
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Obtaining and Identification of Isoespintanol
4.3. Strains
4.4. Phylogenomics of Candida tropicalis
4.4.1. Extraction of Genomic DNA from Candida tropicalis
4.4.2. WGS (Whole Genome Shotgun) Genomic Sequencing of Candida tropicalis on the Illumina Novaseq Platform
4.5. Antifungal Susceptibility Testing
4.6. MTT-Reduction Assay
4.7. Growth Inhibition Curves
4.8. LIVE/DEAD Assays
4.9. Quantitative Assessment of Biofilm Formation
4.10. Effect of Isoespintanol on Cell Membrane Integrity
4.11. Leakage of Nucleic Acids and Proteins through the Fungal Membrane
4.12. Measurement of Extracellular pH
4.13. Transmission Electron Microscopy (TEM)
4.14. Effect of Isoespintanol on the Production of Intracellular Reactive Oxygen Species (ROS)
4.15. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Morais, A.; Araujo, H.; Arias, L.; Ramírez, W.; Porangaba, G.; Penha, S.; Pelim, J.; Monteiro, D. Nanocarriers of miconazole or fluconazole: Effects on three-species Candida biofilms and cytotoxic effects in vitro. J. Fungi 2021, 7, 500. [Google Scholar] [CrossRef]
- Boonsilp, S.; Homkaew, A.; Phumisantiphong, U.; Nutalai, D.; Wongsuk, T. Species distribution, antifungal susceptibility, and molecular epidemiology of Candida species causing candidemia in a tertiary care hospital in Bangkok, Thailand. J. Fungi 2021, 7, 577. [Google Scholar] [CrossRef] [PubMed]
- Steinmann, J.; Schrauzer, T.; Kirchhoff, L.; Meis, J.; Rath, P. Two Candida auris cases in Germany with no recent contact to foreign healthcare—Epidemiological and microbiological investigations. J. Fungi 2021, 7, 380. [Google Scholar] [CrossRef]
- Hassan, Y.; Chew, S.; Lung, L. Candida glabrata: Pathogenicity and resistance mechanisms for adaptation and survival. J. Fungi 2021, 7, 667. [Google Scholar] [CrossRef]
- Chen, P.; Chuang, Y.; Wu, U.; Sun, H.; Wang, J.; Sheng, W.; Chen, Y.; Chang, S. Mechanisms of azole resistance and trailing in Candida tropicalis bloodstream isolates. J. Fungi 2021, 7, 612. [Google Scholar] [CrossRef]
- El-kholy, M.; Helaly, G.; El Ghazzawi, E.; El-Sawaf, G.; Shawky, S. Virulence factors and antifungal susceptibility profile of C. tropicalis isolated from various clinical specimens in Alexandria, Egypt. J. Fungi 2021, 7, 351. [Google Scholar] [CrossRef]
- Zuza, D.; Sila, W.; Chaves, G. An update on Candida tropicalis based on basic and clinical approaches. Front. Microbiol. 2017, 8, 1927. [Google Scholar] [CrossRef]
- Munhoz, N.; Nishiyama, L.; Viero, R.; Bagagli, E.; Schatzmann, P.; Sartori, A.; Fraga, T. Candida tropicalis systemic infection redirects leukocyte infiltration to the kidneys attenuating encephalomyelitis. J. Fungi 2021, 7, 757. [Google Scholar] [CrossRef]
- Cortés, J.; Ruiz, J.; Melgarejo, L.; Lemos, E. Candidemia in Colombia. Biomedica 2020, 40, 195–207. [Google Scholar] [CrossRef]
- Silva, S.; Negri, M.; Henriques, M.; Oliveira, R.; Williams, D.; Azeredo, J. Candida glabrata, Candida parapsilosis and Candida tropicalis: Biology, epidemiology, pathogenicity and antifungal resistance. FEMS Microbiol. Rev. 2012, 36, 288–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aylate, A.; Agize, M.; Ekero, D.; Kiros, A.; Ayledo, G.; Gendiche, K. In-vitro and in-vivo antibacterial activities of Croton macrostachyus methanol extract against E. coli and S. aureus. Adv. Anim. Vet. Sci. 2017, 5, 107–114. [Google Scholar] [CrossRef]
- Avato, P. Editorial to the special issue—“Natural Products and Drug Discovery”. Molecules 2020, 25, 1128. [Google Scholar] [CrossRef]
- Morales, I.; De La Fuente, J.; Sosa, V. Componentes de Eupatorium saltense. An. Asoc. Quim. Argent. 1991, 79, 141–144. [Google Scholar]
- Hocquemiller, R.; Cortes, D.; Arango, G.J.; Myint, S.H.; Cave, A. Isolement et synthese de l’espintanol, nouveau monoterpene antiparasitaire. J. Nat. Prod. 1991, 54, 445–452. [Google Scholar] [CrossRef]
- Rojano, B.; Saez, J.; Schinella, G.; Quijano, J.; Vélez, E.; Gil, A.; Notario, R. Experimental and theoretical determination of the antioxidant properties of isoespintanol (2-isopropyl-3,6-dimethoxy-5-methylphenol). J. Mol. Struct. 2008, 877, 1–6. [Google Scholar] [CrossRef]
- Rojano, B.; Pérez, E.; Figadère, B.; Martin, M.; Recio, M.; Giner, R.; Ríos, J.; Schinella, G.; Sáez, J. Constituents of Oxandra Cf. xylopioides with anti-inflammatory activity. J. Nat. Prod. 2007, 70, 835–838. [Google Scholar] [CrossRef]
- Gavilánez, T.; Colareda, G.; Ragone, M.; Bonilla, M.; Rojano, B.; Schinella, G.; Consolini, A. Intestinal, urinary and uterine antispasmodic effects of isoespintanol, metabolite from Oxandra xylopioides Leaves. Phytomedicine 2018, 51, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, G.; Rojano, B.; Schinella, G.; Mosca, S. Participation of NO in the vasodilatory action of isoespintanol. Vitae 2019, 26, 78–83. [Google Scholar] [CrossRef]
- Usuga, A.; Tejera, I.; Gómez, J.; Restrepo, O.; Rojano, B.; Restrepo, G. Cryoprotective effects of ergothioneine and isoespintanol on canine semen. Animals 2021, 11, 2757. [Google Scholar] [CrossRef]
- Rojano, B.; Montoya, S.; Yépez, F.; Saez, J. Evaluación de isoespintanol aislado de Oxandra cf. xylopioides (Annonaceae) sobre Spodoptera frugiperda J.E. Smith (Lepidoptera: Noctuidae). Rev. Fac. Nac. Agron. Medellín 2007, 60, 3691–3702. [Google Scholar]
- Arango, N.; Vanegas, N.; Saez, J.; García, C.; Rojano, B. Actividad antifungica del isoespintanol sobre hongos del género Colletotricum. Sci. Tech. 2007, 33, 279–280. [Google Scholar] [CrossRef]
- Byvaltsev, V.; Bardonova, L.; Onaka, N.; Polkin, R.; Ochkal, S.; Shepelev, V.; Aliyev, M.; Potapov, A. Acridine orange: A review of novel applications for surgical cancer imaging and therapy. Front. Oncol. 2019, 9, 925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhang, T.; Guo, S.; Zhang, Y.; Sheng, R.; Sun, R.; Chen, L.; Lv, R.; Qi, Y. In vitro antifungal activity and mechanism of Ag3PW12O40 composites against Candida species. Molecules 2020, 25, 6012. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Chen, S.; Levin, R. Application of ethidium bromide monoazide for quantification of viable and dead cells of Salmonella enterica by real-time loop-mediated isothermal amplification. J. Microbiol. Methods 2015, 117, 41–48. [Google Scholar] [CrossRef]
- Donadu, M.; Peralta, Y.; Usai, D.; Maggio, F.; Molina, J.; Rizzo, D.; Bussu, F.; Rubino, S.; Zanetti, S.; Paparella, A.; et al. Colombian essential oil of Ruta graveolens against nosocomial antifungal resistant Candida strains. J. Fungi 2021, 7, 383. [Google Scholar] [CrossRef]
- Iraji, A.; Yazdanpanah, S.; Alizadeh, F.; Mirzamohammadi, S.; Ghasemi, Y.; Pakshir, K.; Yang, Y.; Zomorodian, K. Screening the antifungal activities of monoterpenes and their isomers against Candida species. J. Appl. Microbiol. 2020, 129, 1541–1551. [Google Scholar] [CrossRef]
- Oliveira, I.; De Oliveira, F.; Araújo, W.; De Oliveira, E.; Albuquerque, E.; Cunha, F.; Formiga, M. Antifungal activity and mode of action of carvacrol against Candida albicans strains. J. Essent. Oil Res. 2013, 25, 138. [Google Scholar] [CrossRef]
- De Castro, R.; Pereira, T.; Dornelas, L.; Silva, G.; Melo, E.; Leite, A. Antifungal activity and mode of action of thymol and its synergism with nystatin against Candida species involved with infections in the oral cavity: An in vitro study. BMC Complement. Altern. Med. 2015, 15, 417. [Google Scholar] [CrossRef]
- Kowalczyk, A.; Przychodna, M.; Sopata, S.; Bodalska, A.; Fecka, I. Thymol and thyme essential oil-new insights into selected therapeutic applications. Molecules 2020, 25, 4125. [Google Scholar] [CrossRef]
- Tao, N.; Ouyang, Q.; Jia, L. Citral inhibits mycelial growth of Penicillium italicum by a membrane damage mechanism. Food Control 2014, 41, 116–121. [Google Scholar] [CrossRef]
- De Oliveira, M.; Araújo, A.; Souza, K.; Cardoso, G.; de Oliveira, E.; de Oliveira, F. Investigation of the antifungal potential of linalool against clinical isolates of fluconazole resistant Trichophyton rubrum. J. Mycol. Med. 2017, 27, 195–202. [Google Scholar] [CrossRef]
- Wei, J.; Bi, Y.; Xue, H.; Wang, Y.; Zong, Y.; Prusky, D. Antifungal activity of cinnamaldehyde against Fusarium sambucinum involves inhibition of ergosterol biosynthesis. J. Appl. Microbiol. 2020, 129, 256–265. [Google Scholar] [CrossRef]
- De Oliveira, F.; Moura, J.; De Oliveira, E. Investigation on mechanism of antifungal activity of eugenol against Trichophyton rubrum. Med. Mycol. 2013, 51, 507–513. [Google Scholar] [CrossRef]
- De Castro, A.; De Oliveira, R.; De Oliveira, E.; De Oliveira, W.; De Oliveira, I. Antifungal activity study of the monoterpene thymol against Cryptococcus neoformans. Nat. Prod. Res. 2018, 34, 2630–2633. [Google Scholar] [CrossRef]
- Zhao, F.; Dong, H.; Wang, Y.; Wang, T.; Yan, Z.; Yan, F.; Zhang, D.; Cao, Y.; Jin, Y. Synthesis and synergistic antifungal effects of monoketone derivatives of curcumin against fluconazole-resistant Candida spp. Med. Chem. Commun. 2017, 8, 1093–1102. [Google Scholar] [CrossRef]
- Qin, G.; Liu, J.; Cao, B.; Li, B.; Tian, S. Hydrogen peroxide acts on sensitive mitochondrial proteins to induce death of a fungal pathogen revealed by proteomic analysis. PLoS ONE 2011, 6, e21945. [Google Scholar] [CrossRef]
- Kawai, A.; Yamagishi, Y.; Mikamo, H. Time-lapse tracking of Candida tropicalis biofilm formation and the antifungal efficacy of liposomal amphotericin B. Jpn. J. Infect. Dis. 2017, 70, 559–564. [Google Scholar] [CrossRef]
- Trombetta, D.; Castelli, F.; Sarpietro, M.; Venuti, V.; Cristani, M.; Daniele, C.; Saija, A.; Mazzanti, G.; Bisignano, G. Mechanisms of antibacterial action of three monoterpenes. Antimicrob. Agents Chemother. 2005, 49, 2474. [Google Scholar] [CrossRef]
- Scott, B.; Eaton, C. Role of reactive oxygen species in fungal cellular differentiations. Curr. Opin. Microbiol. 2008, 11, 488–493. [Google Scholar] [CrossRef]
- Zorov, D.; Juhaszova, M.; Sollott, S. Mitochondrial ROS-induced ROS release: An update and review. Biochim. Biophys. Acta 2006, 1757, 509–517. [Google Scholar] [CrossRef]
- Choi, H.; Lee, D. Lycopene induces apoptosis in Candida albicans through reactive oxygen species production and mitochondrial dysfunction. Biochimie 2015, 115, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, M.; Baronetti, J.; Páez, P.; Paraje, M. Oxidative imbalance in Candida tropicalis biofilms and its relation with persister cells. Front. Microbiol. 2021, 11, 598834. [Google Scholar] [CrossRef]
- Zhang, Z.; Qin, G.; Li, B.; Tian, S. Effect of cinnamic acid for controlling gray mold on table grape and its possible mechanisms of action. Curr. Microbiol. 2015, 71, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Rojano, B.; Gaviria, C.; Sáez, J. Determinación de la actividad antioxidante en un modelo de peroxidación lipídica de mantequilla inhibida por el isoespintanol. Vitae 2008, 15, 212–218. [Google Scholar]
- Zapata, K.; Arias, J.; Cortés, F.; Alarcon, C.; Durango, D.; Rojano, B. Oxidative stabilization of palm olein with isoespintanol (2-isopropyl-3,6-dimethoxy-5-methylphenol) isolated from Oxandra cf xylopioides. J. Med. Plants Res. 2017, 11, 218–225. [Google Scholar] [CrossRef]
- Guembe, M.; Cruces, R.; Peláez, T.; Muñoz, P.; Bouza, E. Assessment of biofilm production in Candida isolates according to species and origin of infection. Enferm. Infecc. Microbiol. Clin. 2017, 35, 37–40. [Google Scholar] [CrossRef]
- Tascini, C.; Sozio, E.; Corte, L.; Sbrana, F.; Scarparo, C.; Ripoli, A.; Bertolino, G.; Merelli, M.; Tagliaferri, E.; Corcione, A.; et al. The role of biofilm forming on mortality in patients with candidemia: A study derived from real world data. Infect. Dis. 2017, 50, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Karpiński, T.; Ożarowski, M.; Seremak, A.; Wolski, H.; Adamczak, A. Plant preparations and compounds with activities against biofilms formed by Candida spp. J. Fungi 2021, 7, 360. [Google Scholar] [CrossRef]
- Marquez, M.; Munoz, D.; Bautista, J.; Zapata, K.; Puertas, M.; Lopez, C.; Rojano, B. Effect of isoespintanol isolated from Oxandra cf. xylopioides against DNA damage of human lymphocytes. Pak. J. Pharm. Sci. 2018, 31, 1777–1782. [Google Scholar]
- Ramírez, R.; Páez, M.; Angulo, A. Obtención de isoespintanol por hidrodestilación y cristalización a partir del extracto bencínico de Oxandra xylopioides. Inf. Tecnol. 2015, 26, 13–18. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Approved Standard, 3rd ed.; CLSI document M27-A3; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008. [Google Scholar]
- Rodriguez, J.; Barchiesi, F.; Bille, J.; Chryssanthou, E.; Cuenca, M.; Denning, D.; Donnelly, J.; Dupont, B.; Fegeler, W.; Moore, C.; et al. Method for determination of minimal inhibitory concentration (MIC) by broth dilution of fermentative yeasts. Clin. Microbiol. Infect. 2003, 9, i–viii. [Google Scholar] [CrossRef]
- Quave, C.; Plano, L.; Pantuso, T.; Bennett, B. Effects of extracts from italian medicinal plants on planktonic growth, biofilm formation and adherence of methicillin-resistant Staphylococcus aureus. J. Ethnopharmacol. 2008, 118, 418–428. [Google Scholar] [CrossRef]
- Maldonado, J.; Casaña, R.; Martínez, I.; San Martín, E. La espectroscopia UV-Vis en la evaluación de la viabilidad de células de cáncer de mama. Lat. Am. J. Phys. Educ. 2018, 12, 1–7. [Google Scholar]
- Da Silva, C.; Campos, R.; Neto, J.; Sampaio, L.; do Nascimento, F.; do AV Sa, L.; Candido, T.; Magalhaes, H.; da Cruz, E.; da Silva, E.; et al. Antifungal activity of β-lapachone against azole-resistant Candida spp. and its aspects upon biofilm formation. Future Microbiol. 2020, 15, 1543–1554. [Google Scholar] [CrossRef]
- Kakar, A.; Holzknecht, J.; Dubrac, S.; Gelmi, M.; Romanelli, A.; Marx, F. New perspectives in the antimicrobial activity of the amphibian temporin B: Peptide analogs are effective inhibitors of Candida albicans growth. J. Fungi 2021, 7, 457. [Google Scholar] [CrossRef]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C. tropicalis | ISO | FLZ | ||
|---|---|---|---|---|
| MIC90 | MIC50 | MFC | MIC90 | |
| CLI 001 | 470 | 261.2 | 500 | 875.1 |
| CLI 002 | 326.6 | 59.38 | 350 | 751 |
| CLI 003 | 413.3 | 124.4 | 400 | 875.1 |
| CLI 004 | 420.8 | 121.5 | 450 | 751 |
| CLI 005 | 500 | 234.6 | 500 | 256 |
| CLI 006 | 463.9 | 179.8 | 450 | 128 |
| CLI 007 | 391.6 | 107 | 400 | 751 |
| Isolations | |||||||
|---|---|---|---|---|---|---|---|
| ISO (µg/mL) | 001 | 002 | 003 | 004 | 005 | 006 | 007 |
| 31.25 | 109.1 | 101.8 | 102.0 | 101.3 | 111.7 | 107.7 | 100.6 |
| 62.5 | 108.6 | 65.9 | 101.7 | 100.6 | 107.1 | 100.8 | 74.0 |
| 125 | 101.3 | 56.3 | 59.2 | 57.4 | 104.6 | 100.1 | 67.0 |
| 250 | 50.0 | 8.7 | 14.2 | 10.2 | 48.0 | 51.7 | 0.0 |
| 500 | 0.9 | 1.8 | 0.3 | 0.0 | 1.3 | 0.9 | 0.0 |
| 1000 | 1.1 | 0.0 | 0.8 | 0.6 | 0.0 | 0.2 | 0.0 |
| MIC50 | 236.8 | 103.1 | 141.8 | 136.4 | 230.9 | 235.2 | 113.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Contreras Martínez, O.I.; Angulo Ortíz, A.; Santafé Patiño, G. Mechanism of Antifungal Action of Monoterpene Isoespintanol against Clinical Isolates of Candida tropicalis. Molecules 2022, 27, 5808. https://doi.org/10.3390/molecules27185808
Contreras Martínez OI, Angulo Ortíz A, Santafé Patiño G. Mechanism of Antifungal Action of Monoterpene Isoespintanol against Clinical Isolates of Candida tropicalis. Molecules. 2022; 27(18):5808. https://doi.org/10.3390/molecules27185808
Chicago/Turabian StyleContreras Martínez, Orfa Inés, Alberto Angulo Ortíz, and Gilmar Santafé Patiño. 2022. "Mechanism of Antifungal Action of Monoterpene Isoespintanol against Clinical Isolates of Candida tropicalis" Molecules 27, no. 18: 5808. https://doi.org/10.3390/molecules27185808