

Crystal Structure of Human CD47 in Complex with Engineered SIRPα.D1(N80A)
Abstract
:1. Introduction
2. Results
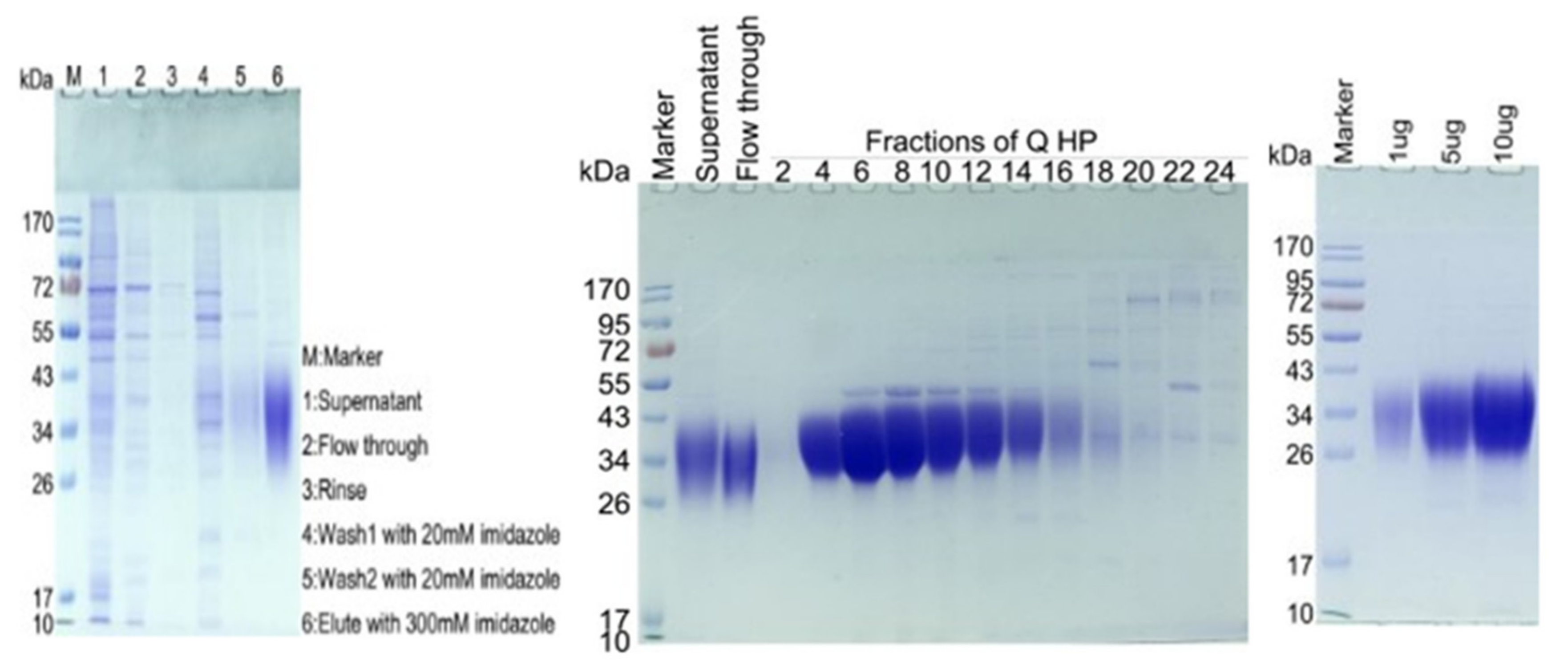
2.1. SDS-PAGE Analysis Results of CD47-His
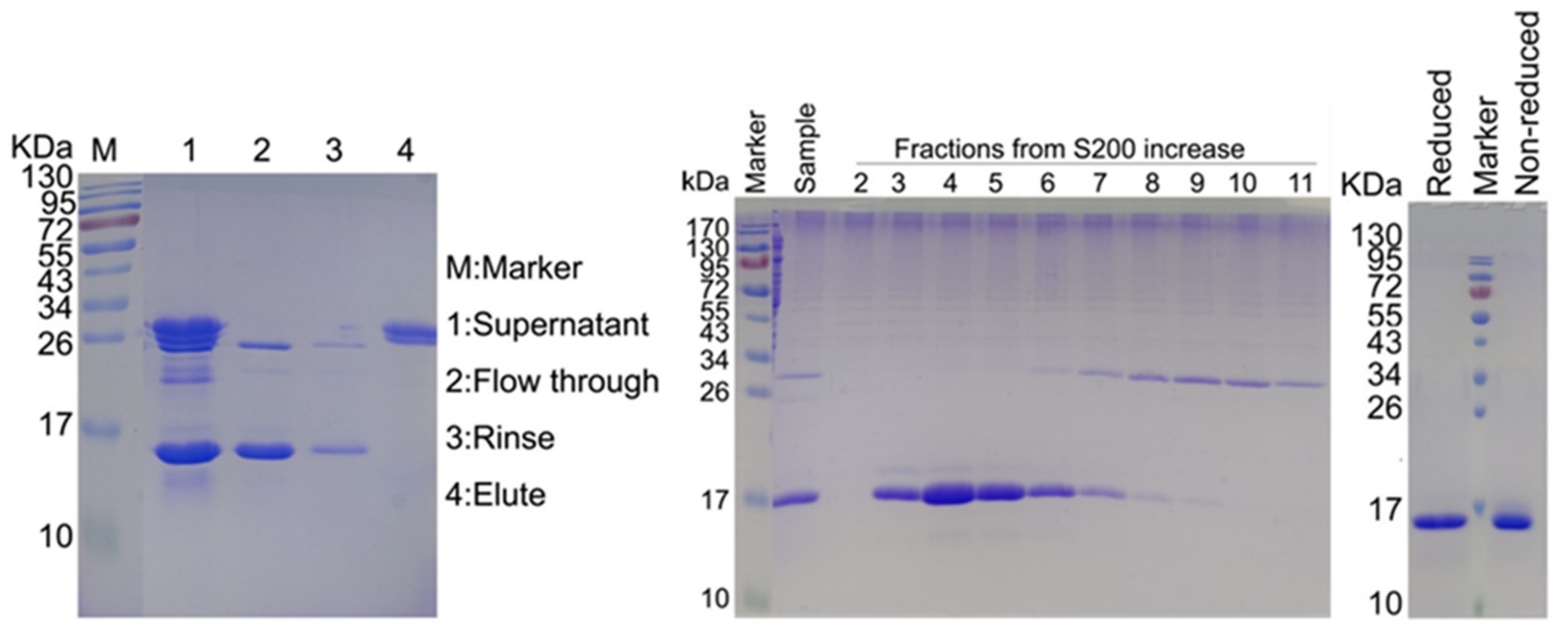
2.2. SDS-PAGE Analysis Results of Purified IMM01-Fab
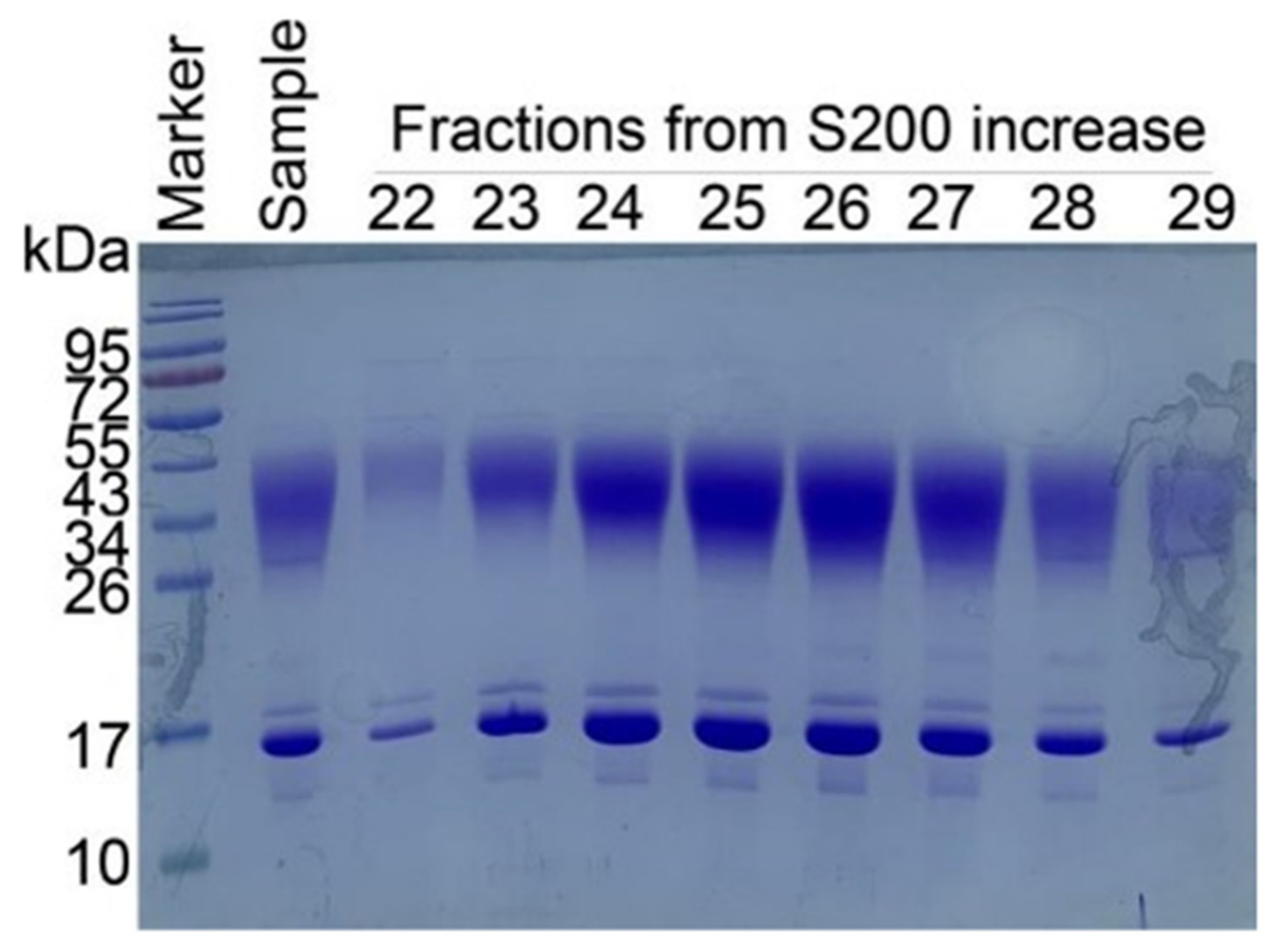
2.3. Confirmation of IMM01 Fusion Protein Expression and Its Molecular Weight
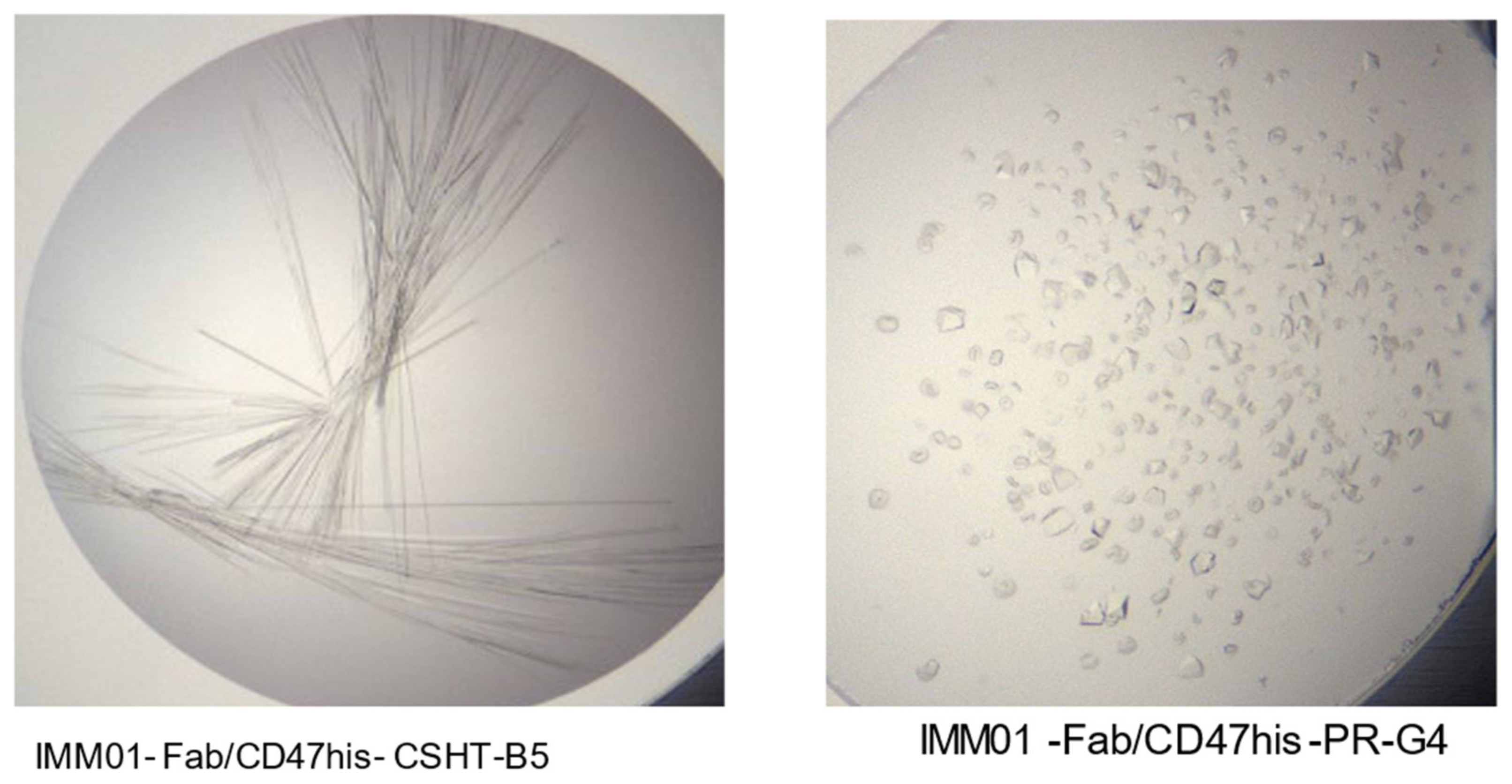
2.4. Crystallization and Structure Determination
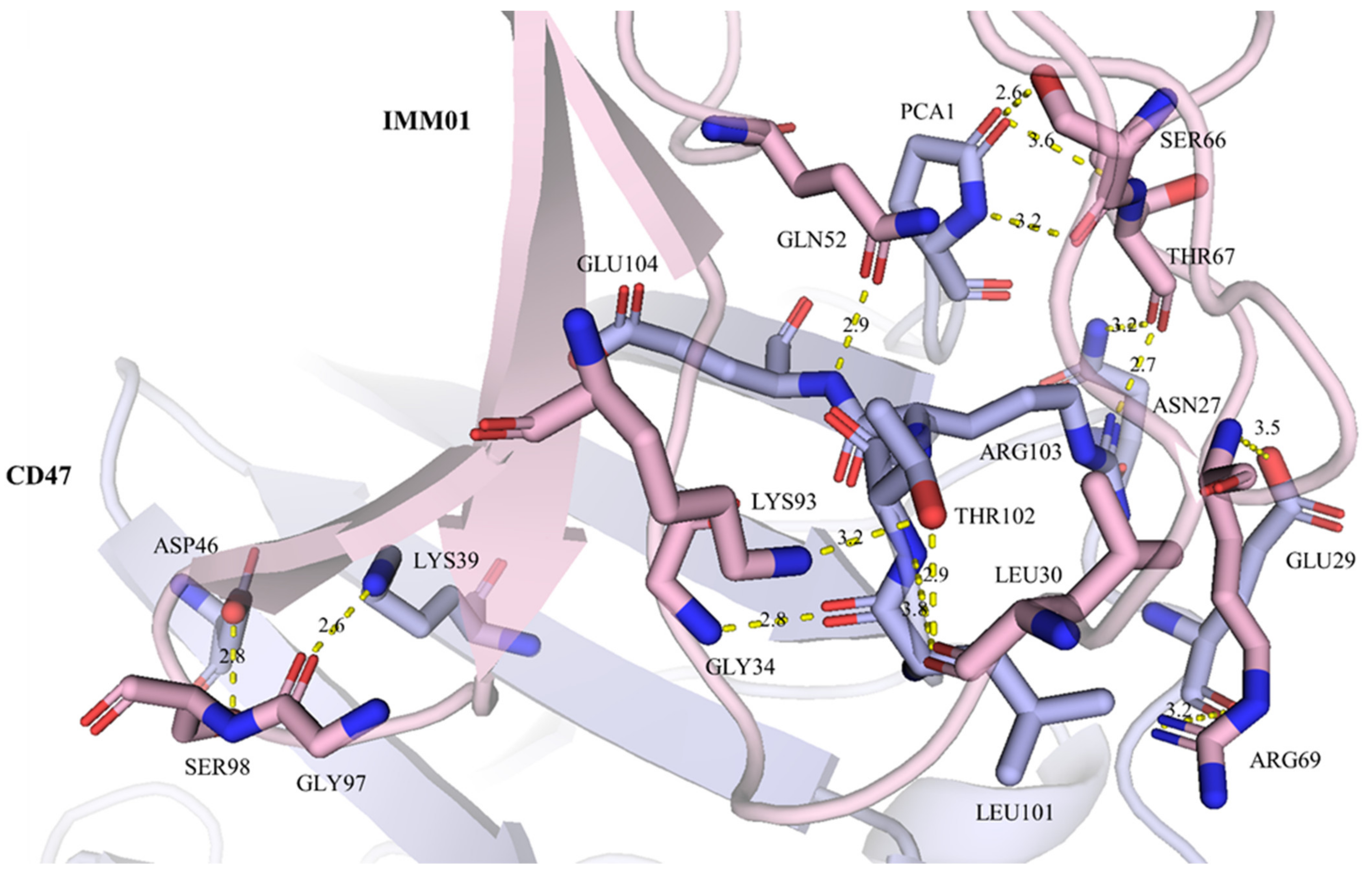
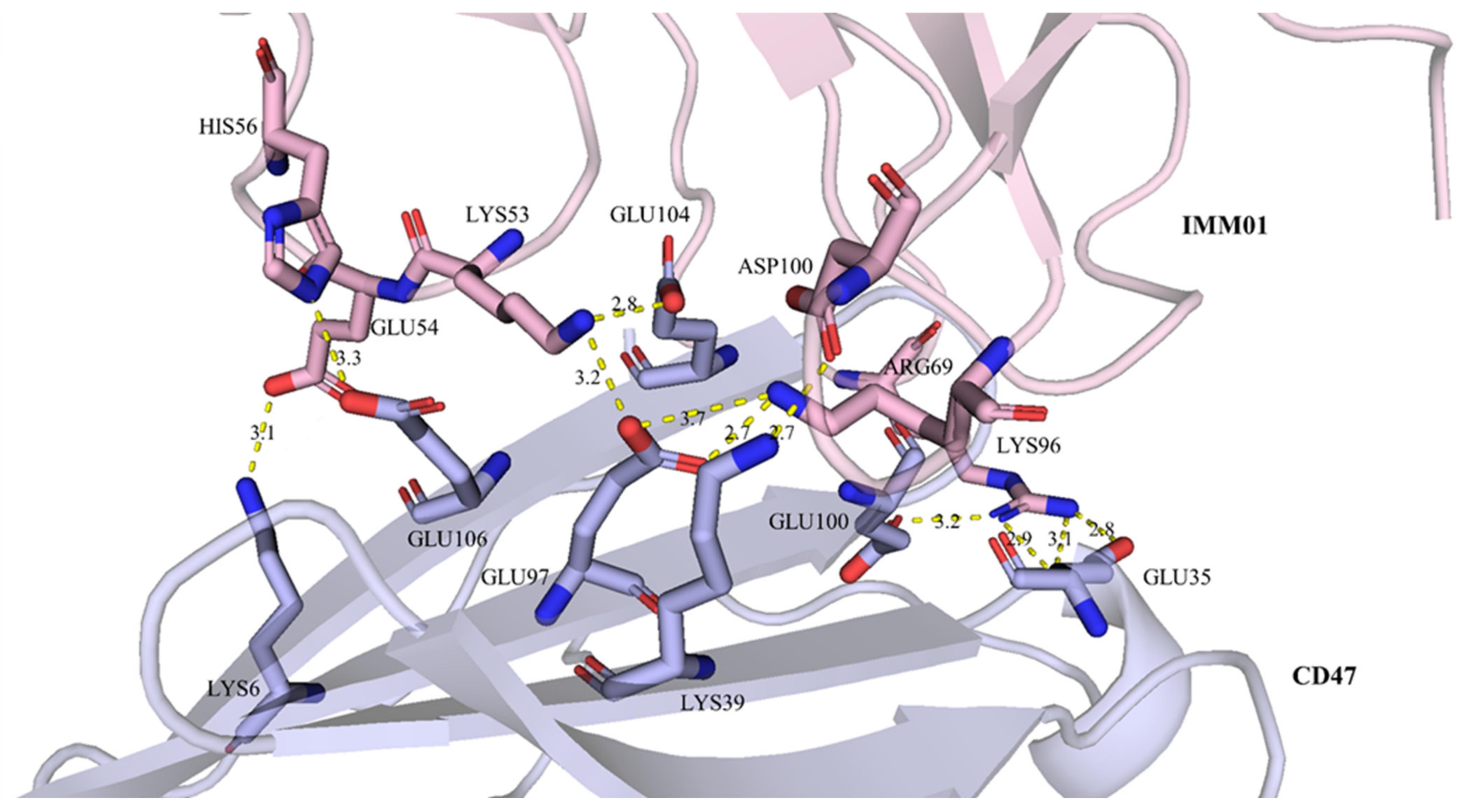
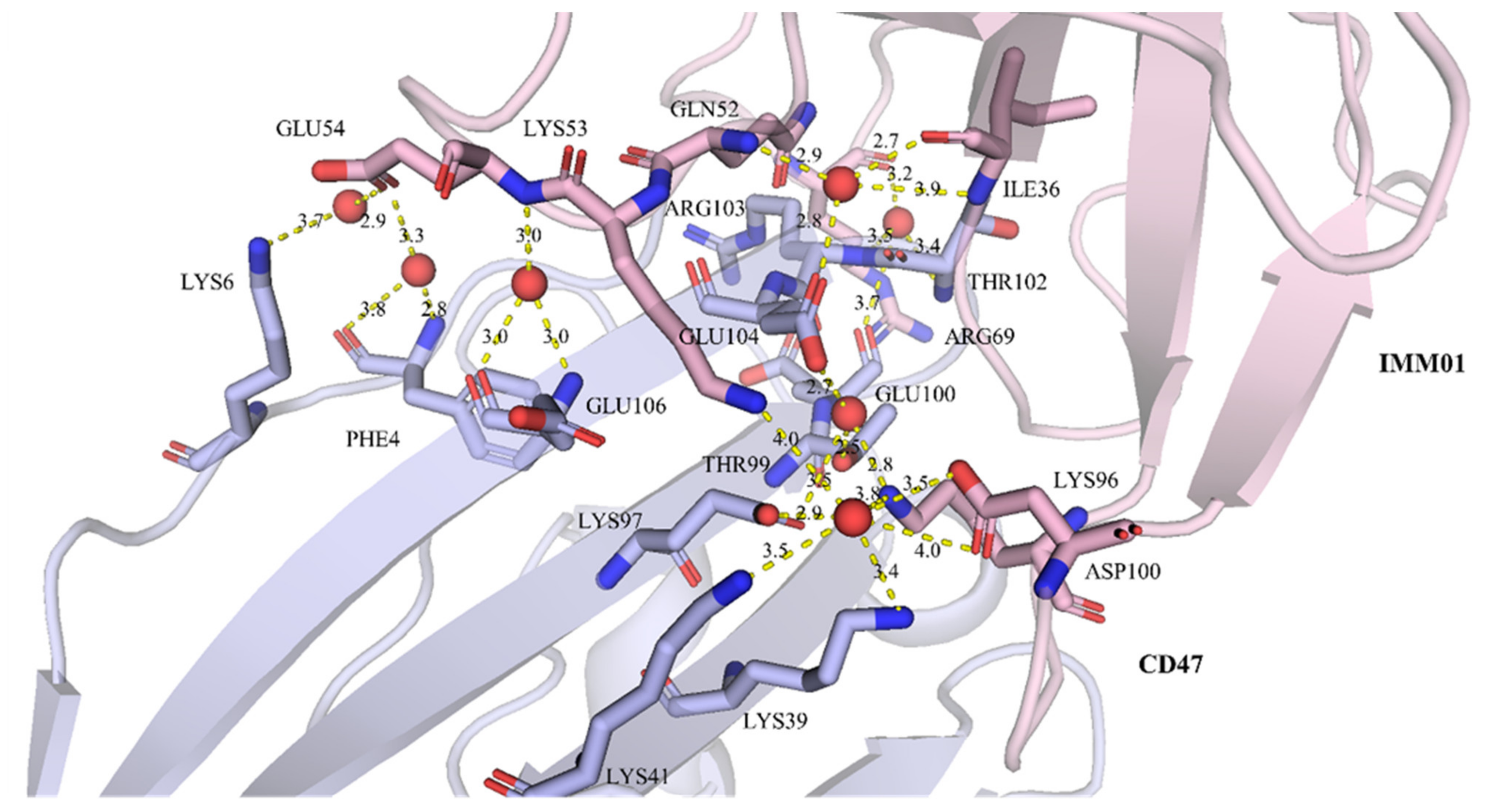
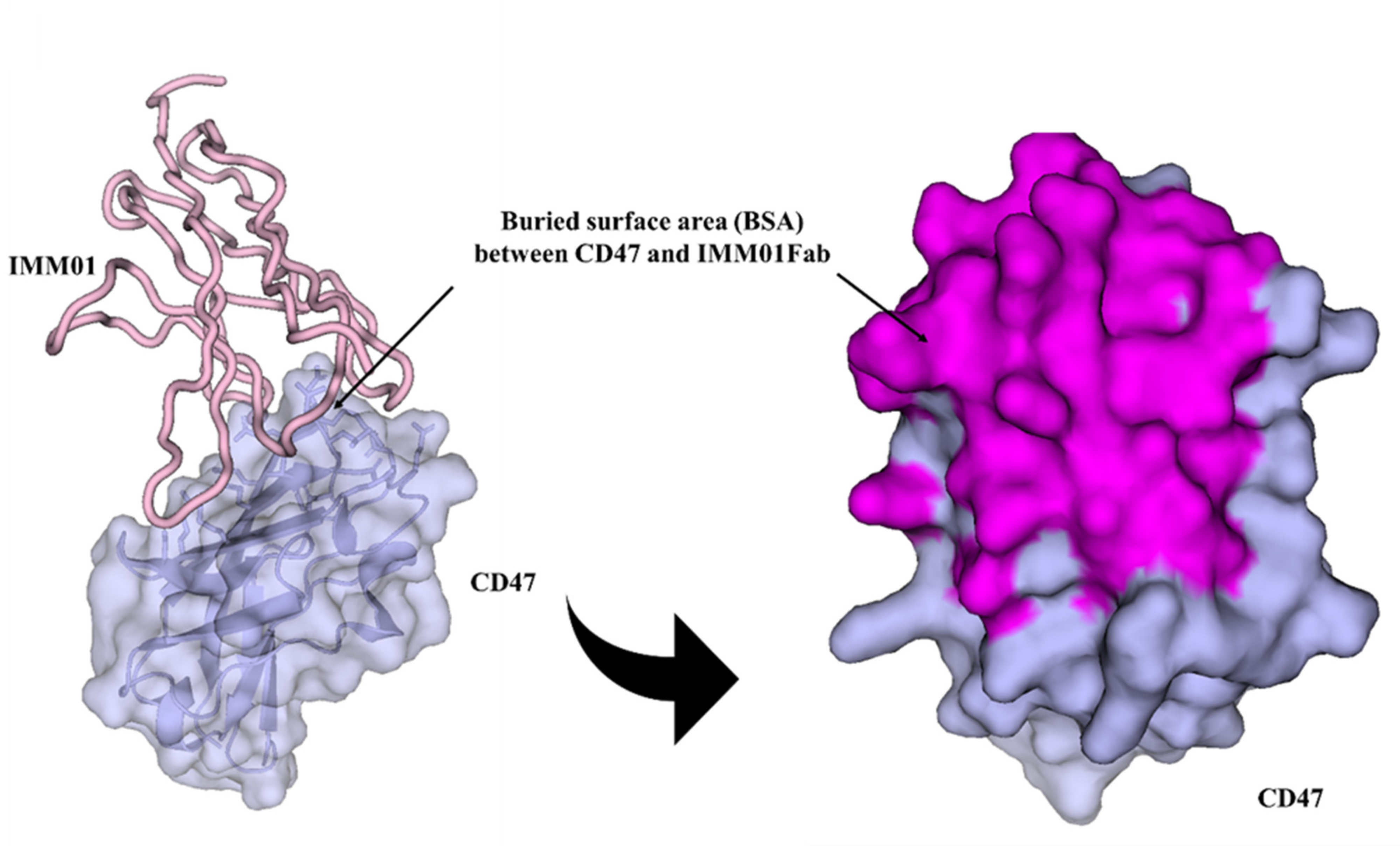
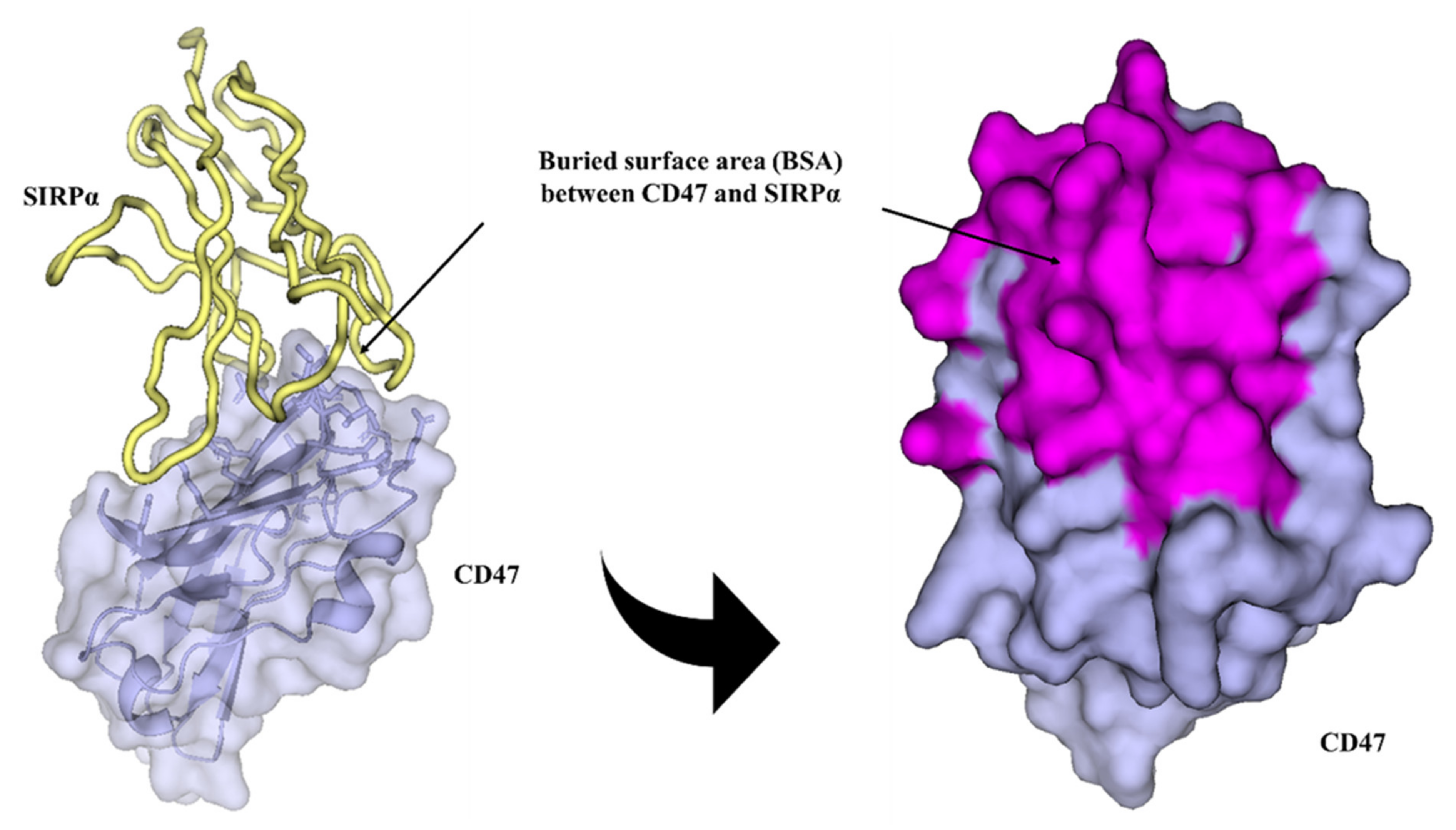
2.5. IMM01/CD47 Complex Interface Analysis
3. Discussion
4. Materials and Methods
4.1. Construction, Expression and Purification of CD47-His
4.1.1. Construction of Plasmid for CD47-His Protein Expression
4.1.2. Protein Expression of CD47-His (for 1 L)
- (1)
- Twenty-four hours prior to transfection, split Expi293F cells back to a density of 2 × 106 cells/mL. Ensure cells are in healthy, log phase growth and viability is >95%.
- (2)
- On the day of transfection, split cells to a density of 2.7–3.0 × 106 cells/mL.
- (3)
- Using the general rule, 1 μg DNA is used per 1 mL of culture, 1 mg of DNA is used for transfection of 1 L culture.
- (4)
- Dilute 1 mg of pTT5-hCD47(19-135)-His plasmid into a final volume of 50 mL Opti-MEM medium.
- (5)
- Dilute 6 mg of PEI transfection reagent into a final volume of 50 mL Opti-MEM medium. Incubate at room temperature for 3 min; the solution will become slightly cloudy.
- (6)
- Mix 10 mL of DNA with 10 mL of PEI and incubate for 15 min at room temperature.
- (7)
- Add 100 mL of the DNA:PEI complex dropwise to the culture. This will bring the final density of the culture to ~2.5 × 106 cells/mL.
- (8)
- Incubate on an orbital shaking platform at 37 °C with 8% CO2 at a speed of 125 rpm.
- (9)
- After incubating cells for 20 h, add cell transfection enhancer to the culture.
- (10)
- Harvest cell supernatants at 120 h post-transfection.
4.1.3. Purification of CD47-His
4.1.4. Q HP Purification
4.2. Construction and Expression of Recombinant Protein IMM01
4.3. Crystallization of IMM01-Fab/CD47 Complex, Data Collection, Structure Determination and Refinement
- (a)
- The hCD47(19-135)-His was constructed into a vector and expressed in Expi293F cells. The Cys at residue 15 was mutated to Gly to minimize disulfide crosslinking. Recombinant CD47 proteins were purified by nickel affinity chromatography and ion exchange chromatography.
- (b)
- IMM01-Fab was fused with the Fc tag in a C-terminal construct into a vector and expressed in CHO cells. The proteins were purified by protein A affinity chromatography and digested with papain to remove the Fc tag.
- (c)
- IMM01-Fab and CD47 were mixed in a 1:1 molar ratio, deglycosylated using endoglycosidase Hf and concentrated to contain each protein at 12.6 mg/mL. Sitting drop vapor diffusion crystallization experiments were performed using an NT8 robot to dispense nanoscale protein precipitant drops that were equilibrated against precipitant reservoirs at 18 °C. Crystals were cryoprotected in mother liquor supplemented with 20% glycerol and flash-frozen in liquid nitrogen.
- (d)
- Diffraction images were collected at 100 K at Beamline 19-ID-D of the Advanced Photon Source (APS, USA) using a Pilatus3 × 6M detector. The dataset, at the resolution of 2.76 Å, was processed with the XDS programs [29] and scaled with AIMLESS from the CCP4 suite [30]. The unit cell parameters for the crystal were a =53.58 Å, b = 99.64 Å, c = 138.93 Å, α = β = γ = 90.0°, in space group P212121 with two copies of the molecules in the IMM01-Fab/CD47 complex in one asymmetric unit. The complex structure was determined by molecular replacement using the program PHASER with the CD47-SIRPαv2 structure (Protein Data Bank identification, PDB ID 2JJT) as a search model. The model was manually built using the COOT program [31] and refined using TLS parameters in REFMAC from the CCP4 program suite [32]. All structural figures were prepared with PyMol [33].
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ADCC | antibody-dependent cell-mediated cytotoxicity |
| ADCP | antibody-dependent cellular phagocytosis |
| CBD | collagen binding domain |
| CD47 | cluster of differentiation 47 |
| IgSF | immunoglobulin superfamily |
| mAbs | monoclonal antibodies |
| PDB ID | Protein Data Bank identification |
| PS | phosphatidylserine |
| SEC | size-exclusion chromatography |
| SE-HPLC | size-exclusion high-performance liquid chromatography |
| SIRPα | signal-regulatory protein alpha |
References
- Jiang, Z.; Sun, H.; Yu, J.; Tian, W.; Song, Y. Targeting CD47 for cancer immunotherapy. J. Hematol. Oncol. 2021, 14, 180. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, C.; Liu, Y.; Wang, C.; Jiang, H.; Hu, Y.; Wu, J. Recent Advances of Tumor Therapy Based on the CD47-SIRPα Axis. Mol. Pharm. 2022, 19, 1273–1293. [Google Scholar] [CrossRef] [PubMed]
- Logtenberg, M.E.; Scheeren, F.A.; Schumacher, T.N. The CD47-SIRPα Immune Checkpoint. Immunity 2020, 52, 742–752. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, K.; Prasolava, T.K.; Wang, J.C.; Mortin-Toth, S.M.; Khalouei, S.; Gan, O.I.; Dick, J.E.; Danska, J.S. Polymorphism in Sirpa modulates engraftment of human hematopoietic stem cells. Nat. Immunol. 2007, 8, 1313–1323. [Google Scholar] [CrossRef]
- Voets, E.; Paradé, M.; Hulsik, D.L.; Spijkers, S.; Janssen, W.; Rens, J.; Reinieren-Beeren, I.; Tillaart, G.V.D.; Van Duijnhoven, S.; Driessen, L.; et al. Functional characterization of the selective pan-allele anti-SIRPα antibody ADU-1805 that blocks the SIRPα-CD47 innate immune checkpoint. J. Immunother. Cancer 2019, 7, 340. [Google Scholar] [CrossRef]
- Huang, B.; Bai, Z.; Ye, X.; Zhou, C.; Xie, X.; Zhong, Y.; Lin, K.; Ma, L. Structural analysis and binding sites of inhibitors targeting the CD47/SIRPα interaction in anticancer therapy. Comput. Struct. Biotechnol. J. 2021, 19, 5494–5503. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Shi, W.; Shi, J.-J.; Lu, J.-J. Progress of CD47 immune checkpoint blockade agents in anticancer therapy: A hematotoxic perspective. J. Cancer. Res. Clin. Oncol. 2022, 148, 1–14. [Google Scholar] [CrossRef]
- Hatherley, D.; Lea, S.; Johnson, S.; Barclay, A.N. Polymorphisms in the human inhibitory signal-regulatory protein α do not affect binding to its ligand CD47. J. Biol. Chem. 2014, 289, 10024–10028. [Google Scholar] [CrossRef]
- Qu, T.; Li, B.; Wang, Y. Targeting CD47/SIRPα as a therapeutic strategy, where we are and where we are headed. Biomark. Res. 2022, 10, 20. [Google Scholar] [CrossRef]
- Hatherley, D.; Graham, S.; Turner, J.; Harlos, K.; Stuart, D.I.; Barclay, A.N. Paired receptor specificity explained by structures of signal regulatory proteins alone and complexed with CD47. Mol. Cell 2008, 31, 266–277. [Google Scholar] [CrossRef]
- Hatherley, D.; Harlos, K.; Dunlop, D.C.; Stuart, D.I.; Barclay, A.N. The structure of the macrophage signal regulatory protein alpha (SIRPalpha) inhibitory receptor reveals a binding face reminiscent of that used by T cell receptors. J. Biol. Chem. 2007, 282, 14567–14575. [Google Scholar] [CrossRef]
- Weiskopf, K.; Ring, A.M.; Ho, C.C.M.; Volkmer, J.-P.; Levin, A.M.; Volkmer, A.K.; Özkan, E.; Fernhoff, N.B.; van de Rijn, M.; Weissman, I.L.; et al. Engineered SIRPα variants as immunotherapeutic adjuvants to anticancer antibodies. Science 2013, 341, 88–91. [Google Scholar] [CrossRef]
- Weiskopf, K.; Jahchan, N.S.; Schnorr, P.; Cristea, S.; Ring, A.; Maute, R.L.; Volkmer, A.K.; Volkmer, J.-P.; Liu, J.; Lim, J.S.; et al. CD47-blocking immunotherapies stimulate macrophage-mediated destruction of small-cell lung cancer. J. Clin. Investig. 2016, 126, 2610–2620. [Google Scholar] [CrossRef]
- Pietsch, E.C.; Dong, J.; Cardoso, R.; Zhang, X.; Chin, D.; Hawkins, R.; Dinh, T.; Zhou, M.; Strake, B.; Feng, P.-H.; et al. Anti-leukemic activity and tolerability of anti-human CD47 monoclonal antibodies. Blood Cancer J. 2017, 7, e536. [Google Scholar] [CrossRef]
- Wu, Z.; Weng, L.; Zhang, T.; Tian, H.; Fang, L.; Teng, H.; Zhang, W.; Gao, J.; Hao, Y.; Li, Y.; et al. Identification of Glutaminyl Cyclase isoenzyme isoQC as a regulator of SIRPα-CD47 axis. Cell Res. 2019, 29, 502–505. [Google Scholar] [CrossRef]
- Logtenberg, M.E.W.; Jansen, J.H.M.; Raaben, M.; Toebes, M.; Franke, K.; Brandsma, A.M.; Matlung, H.L.; Fauster, A.; Gomez-Eerland, R.; Bakker, N.A.M.; et al. Glutaminyl cyclase is an enzymatic modifier of the CD47-SIRPα axis and a target for cancer immunotherapy. Nat. Med. 2019, 25, 612–619. [Google Scholar] [CrossRef]
- Orozco-Morales, M.; Avilés-Salas, A.; Hernández-Pedro, N.; Catalán, R.; Cruz-Rico, G.; Colín-González, A.L.; Dosal-Mancilla, E.; Barrios-Bernal, P.; Arrieta, O. Clinicopathological and Prognostic Significance of CD47 Expression in Lung Neuroendocrine Tumors. J. Immunol. Res. 2021, 2021, 6632249. [Google Scholar] [CrossRef]
- Andrejeva, G.; Capoccia, B.J.; Hiebsch, R.R.; Donio, M.J.; Darwech, I.M.; Puro, R.J.; Pereira, D.S. Novel SIRPα Antibodies That Induce Single-Agent Phagocytosis of Tumor Cells while Preserving T Cells. J. Immunol. 2021, 206, 712–721. [Google Scholar] [CrossRef]
- Sikic, B.I.; Lakhani, N.; Patnaik, A.; Shah, S.A.; Chandana, S.R.; Rasco, D.; Colevas, A.D.; O’Rourke, T.; Narayanan, S.; Papadopoulos, K.; et al. First-in-Human, First-in-Class Phase I Trial of the Anti-CD47 Antibody Hu5F9-G4 in Patients with Advanced Cancers. J. Clin. Oncol. 2019, 37, 946–953. [Google Scholar] [CrossRef]
- Advani, R.; Flinn, I.; Popplewell, L.; Forero, A.; Bartlett, N.L.; Ghosh, N.; Kline, J.; Roschewski, M.; LaCasce, A.; Collins, G.P.; et al. CD47 Blockade by Hu5F9-G4 and Rituximab in Non-Hodgkin’s Lymphoma. N. Engl. J. Med. 2018, 379, 1711–1721. [Google Scholar] [CrossRef] [PubMed]
- Bewersdorf, J.; Zeidan, A. Risk-Adapted, Individualized Treatment Strategies of Myelodysplastic Syndromes (MDS) and Chronic Myelomonocytic Leukemia (CMML). Cancers 2021, 13, 1610. [Google Scholar] [CrossRef] [PubMed]
- Sifniotis, V.; Cruz, E.; Eroglu, B.; Kayser, V. Current Advancements in Addressing Key Challenges of Therapeutic Antibody Design, Manufacture, and Formulation. Antibodies 2019, 8, 36. [Google Scholar] [CrossRef]
- Ren, T.; Tan, Z.; Ehamparanathan, V.; Lewandowski, A.; Ghose, S.; Li, Z.J. Antibody disulfide bond reduction and recovery during biopharmaceutical process development—A review. Biotechnol. Bioeng. 2021, 118, 2829–2844. [Google Scholar] [CrossRef] [PubMed]
- Kitten, O.; Martineau, P. Antibody alternative formats: Antibody fragments and new frameworks. Med. Sci. 2019, 35, 1092–1097. [Google Scholar]
- Ma, H.; Ó’Fágáin, C.; O’Kennedy, R. Antibody stability: A key to performance—Analysis, influences and improvement. Biochimie 2020, 177, 213–225. [Google Scholar] [CrossRef]
- Liu, J.; Meng, Z.; Xu, T.; Kuerban, K.; Wang, S.; Zhang, X.; Fan, J.; Ju, D.; Tian, W.; Huang, X.; et al. A SIRPαFc Fusion Protein Conjugated with the Collagen-Binding Domain for Targeted Immunotherapy of Non-Small Cell Lung Cancer. Front. Immunol. 2022, 13, 845217. [Google Scholar] [CrossRef]
- Huang, Y.; Lv, S.; Liu, P.; Ye, Z.; Yang, H.; Li, L.; Zhu, H.; Wang, Y.; Cui, L.; Jiang, D.; et al. A SIRPα-Fc fusion protein enhances the antitumor effect of oncolytic adenovirus against ovarian cancer. Mol. Oncol. 2020, 14, 657–668. [Google Scholar] [CrossRef]
- Tahk, S.; Vick, B.; Hiller, B.; Schmitt, S.; Marcinek, A.; Perini, E.D.; Leutbecher, A.; Augsberger, C.; Reischer, A.; Tast, B.; et al. SIRPα-αCD123 fusion antibodies targeting CD123 in conjunction with CD47 blockade enhance the clearance of AML-initiating cells. J. Hematol. Oncol. 2021, 14, 155. [Google Scholar] [CrossRef]
- Kabsch, W. Automatic processing of rotation diffraction data from crystals of initially unknown symmetry and cell constants. J. Appl. Cryst. 1993, 26, 795–800. [Google Scholar] [CrossRef]
- Evans, P.R.; Murshudov, G.N. How good are my data and what is the resolution? Acta Crystallogr. D Biol. Crystallogr. 2013, 69, 1204–1214. [Google Scholar] [CrossRef]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [Green Version]
- The CCP4 Suite: Programs for protein crystallography. Acta Crystallogr. D Biol. Crystallogr. 1994, 50, 760–763.
- Schrödinger, L.; DeLano, W. Pymol. Available online: http://www.pymol.org/pymol (accessed on 20 July 2022).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IMM01 | CD47 | Distance (Å) | ||
|---|---|---|---|---|
| Residue | Atom | Residue | Atom | |
| A:SER66 | O | C:GLN1 | N | 2.7 |
| A:SER66 | OG | C:GLN1 | NE2 | 2.4 |
| A:THR67 | O | C:ASN27 | ND2 | 3.1 |
| A:GLY97 | O | C:LYS39 | NZ | 2.6 |
| A:LEU30 | O | C:THR102 | OG1 | 2.9 |
| A:THR67 | O | C:ARG103 | NH1 | 2.6 |
| A:GLN52 | OE1 | C:GLU104 | N | 2.8 |
| A:GLY34 | N | C:LEU101 | O | 2.7 |
| A:ARG69 | N | C:GLU29 | OE2 | 3.4 |
| A:ARG69 | NH1 | C:GLU29 | O | 3.3 |
| A:SER98 | OG | C:ASP46 | OD2 | 2.8 |
| A:LYS93 | NZ | C:THR102 | OG1 | 3.2 |
| A:ASP73 | OD1 | C:TYR113 | OH | 2.4 |
| A:GLU65 | N | C:VAL116 | O | 3.2 |
| A:SER64 | OG | C:VAL116 | O | 2.7 |
| B:SER66 | O | D:GLN1 | N | 3.3 |
| B:SER66 | OG | D:GLN1 | NE2 | 2.2 |
| B:TYR50 | OH | D:GLN1 | NE2 | 3.4 |
| B:THR67 | O | D:ASN27 | ND2 | 3.3 |
| B:GLY97 | O | D:LYS39 | NZ | 2.7 |
| B:LEU30 | O | D:THR102 | N | 3.8 |
| B:LEU30 | O | D:THR102 | OG1 | 2.8 |
| B:THR67 | O | D:ARG103 | NH1 | 2.5 |
| B:GLN52 | OE1 | D:GLU104 | N | 3.2 |
| B:GLY34 | N | D:LEU101 | O | 2.7 |
| B:GLN52 | OE1 | D:GLU104 | N | 3.2 |
| B:ARG69 | N | D:GLU29 | OE2 | 3.5 |
| B:SER98 | OG | D:ASP46 | OD2 | 2.9 |
| B:LYS93 | NZ | D:THR102 | OG1 | 3 |
| B:GLU70 | OE2 | D:TYR113 | OH | 2.7 |
| B:GLU70 | OE1 | D:VAL116 | N | 3.5 |
| B:ARG69 | NH1 | D:VAL116 | O | 2.8 |
| IMM01 | CD47 | Distance (Å) | ||
|---|---|---|---|---|
| Residue | Atom | Residue | Atom | |
| A:GLU54 | OE2 | C:LYS6 | NZ | 3 |
| A:ASP100 | OD1 | C:LYS39 | NZ | 2.7 |
| A:LYS53 | NZ | C:GLU97 | OE2 | 3.2 |
| A:LYS53 | NZ | C:GLU104 | OE2 | 2.8 |
| A:HIS56 | NE2 | C:GLU106 | OE2 | 3.3 |
| A:ARG69 | NH2 | C:GLU35 | OE1 | 3.1 |
| A:ARG69 | NH2 | C:GLU35 | OE2 | 2.9 |
| A:ARG69 | NH1 | C:GLU100 | OE1 | 3.2 |
| A:ARG69 | NH1 | C:GLU35 | OE1 | 2.8 |
| A:LYS96 | NZ | C:GLU97 | OE1 | 2.7 |
| B:GLU54 | OE2 | D:LYS6 | NZ | 2.6 |
| B:ASP100 | OD1 | D:LYS39 | NZ | 2.7 |
| B:LYS53 | NZ | D:GLU106 | OE1 | 3.4 |
| B:LYS53 | NZ | D:GLU97 | OE2 | 2.7 |
| B:ARG69 | NE | D:GLU35 | OE1 | 3 |
| B:ARG69 | NH1 | D:GLU35 | OE1 | 3.4 |
| B:ARG69 | NH1 | D:GLU35 | OE2 | 3 |
| B:LYS96 | NZ | D:GLU97 | OE1 | 2.5 |
| IMM01 | Distance (Å) | Water ID | CD47 | Distance (Å) | ||
|---|---|---|---|---|---|---|
| Residue | Atom | Residue | Atom | |||
| A:ASP100 | OD2 | 3.5 | 24 | C:LYS39 | NZ | 3.4 |
| C:LYS41 | NZ | 3.5 | ||||
| C:GLU97 | OE2 | 2.9 | ||||
| A:LYS96 | NZ | 2.8 | 58 | C:THR99 | OG1 | 2.5 |
| C:GLU104 | OE2 | 2.7 | ||||
| C:GLU97 | OE1 | 3.5 | ||||
| A:ILE36 | O | 2.7 | 57 | C:GLU104 | OE1 | 2.8 |
| A:GLN52 | N | 2.9 | ||||
| A:GLU54 | N | 3 | 55 | C:GLU106 | N | 3 |
| C:GLU106 | O | 3 | ||||
| OE1 | 3.2 | 47 | C:PHE4 | N | 2.9 | |
| A:ARG69 | O | 3.3 | 93 | C:THR102 | N | 3.4 |
| C:ARG103 | N | 3.5 | ||||
| B:ASP100 | OD1 | 3.4 | 5 | D:LYS39 | NZ | 3.3 |
| D:GLU97 | OE2 | 3.1 | ||||
| B:LYS96 | NZ | 2.9 | 41 | D:THR99 | OG1 | 2.7 |
| D:GLU104 | OE2 | 2.8 | ||||
| B:GLU54 | N | 3.1 | 36 | D:GLU106 | O | 2.9 |
| N | 3 | |||||
| B:ARG69 | N | 3.1 | 90 | D:GLU29 | OE1 | 2.6 |
| B:GLU70 | N | 3.1 | OE2 | 3.5 | ||
| B:ARG69 | NH2 | 3 | 16 | D:GLU29 | OE1 | 3.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.; Li, S.; Chen, D.; Liu, D.; Guo, H.; Yang, C.; Zhang, W.; Zhang, L.; Zhao, G.; Tu, X.; et al. Crystal Structure of Human CD47 in Complex with Engineered SIRPα.D1(N80A). Molecules 2022, 27, 5574. https://doi.org/10.3390/molecules27175574
Yu J, Li S, Chen D, Liu D, Guo H, Yang C, Zhang W, Zhang L, Zhao G, Tu X, et al. Crystal Structure of Human CD47 in Complex with Engineered SIRPα.D1(N80A). Molecules. 2022; 27(17):5574. https://doi.org/10.3390/molecules27175574
Chicago/Turabian StyleYu, Jifeng, Song Li, Dianze Chen, Dandan Liu, Huiqin Guo, Chunmei Yang, Wei Zhang, Li Zhang, Gui Zhao, Xiaoping Tu, and et al. 2022. "Crystal Structure of Human CD47 in Complex with Engineered SIRPα.D1(N80A)" Molecules 27, no. 17: 5574. https://doi.org/10.3390/molecules27175574