
Organic Acids Secreted by Lactobacillus spp. Isolated from Urine and Their Antimicrobial Activity against Uropathogenic Proteus mirabilis


Abstract
:1. Introduction
2. Results
2.1. Antibiotic Susceptibility Profile of Proteus Mirabilis Strains
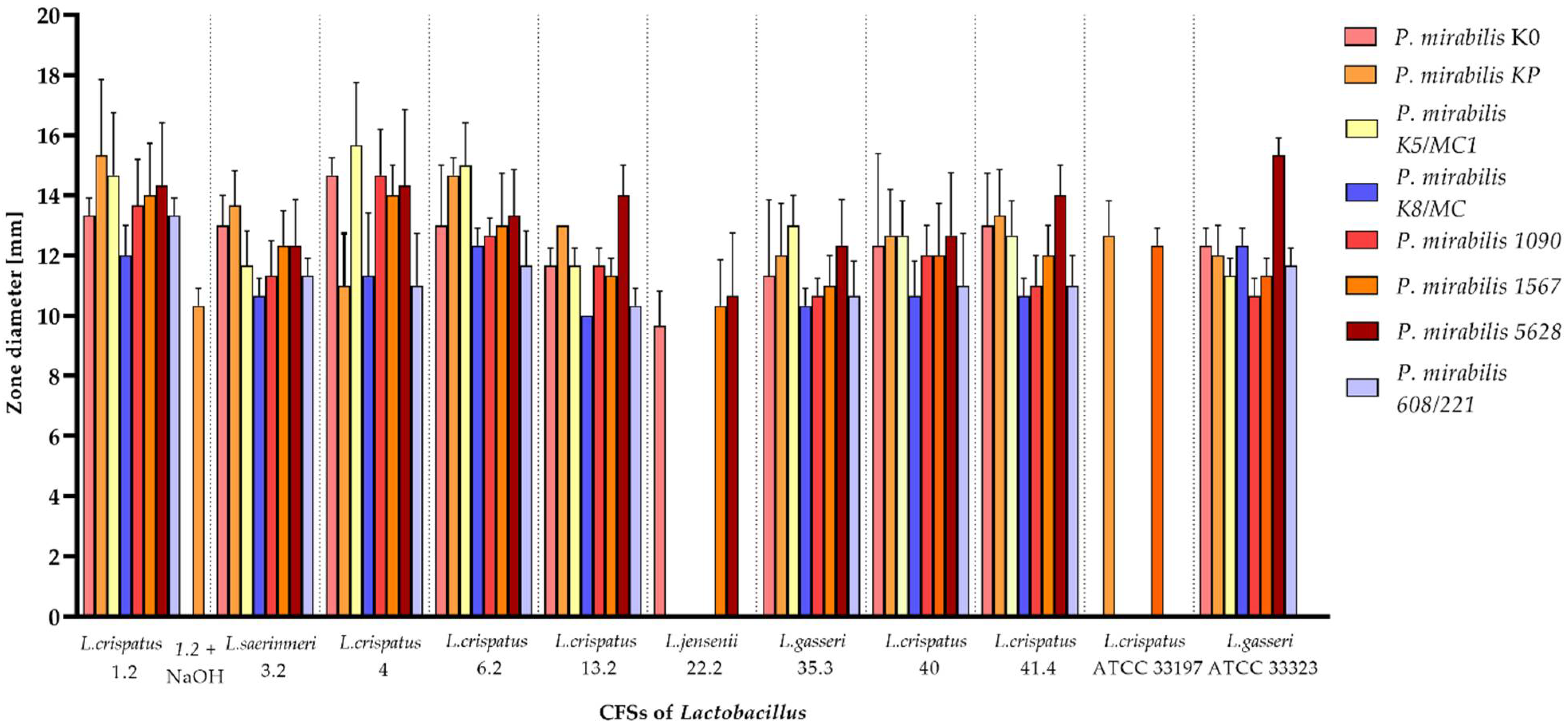
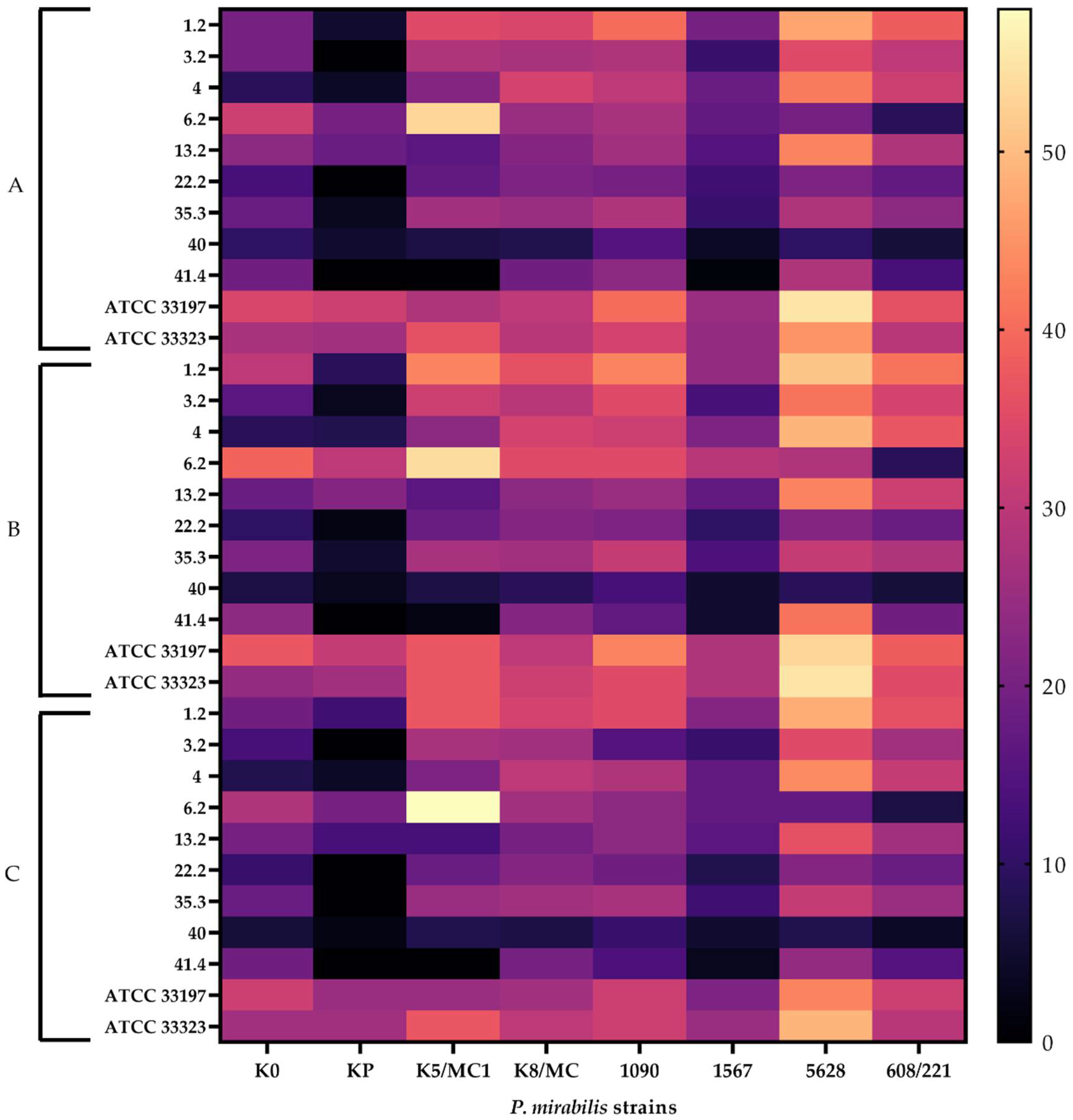
2.2. Antibacterial Effect of Lactobacillus CFSs on Pathogenic P. mirabilis Strains
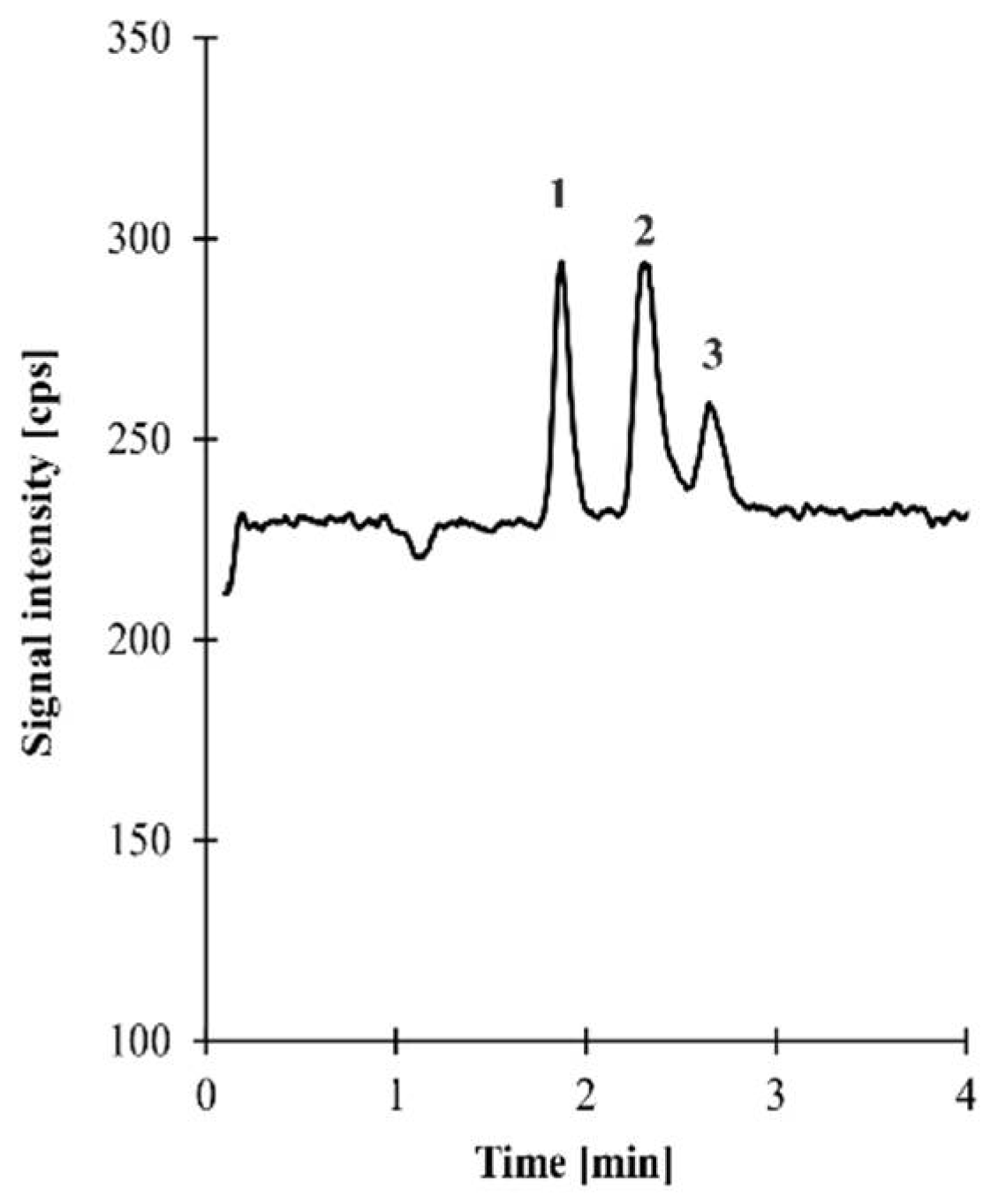
2.3. Profile of Organic Acids Produced by the Tested Lactobacillus Strains
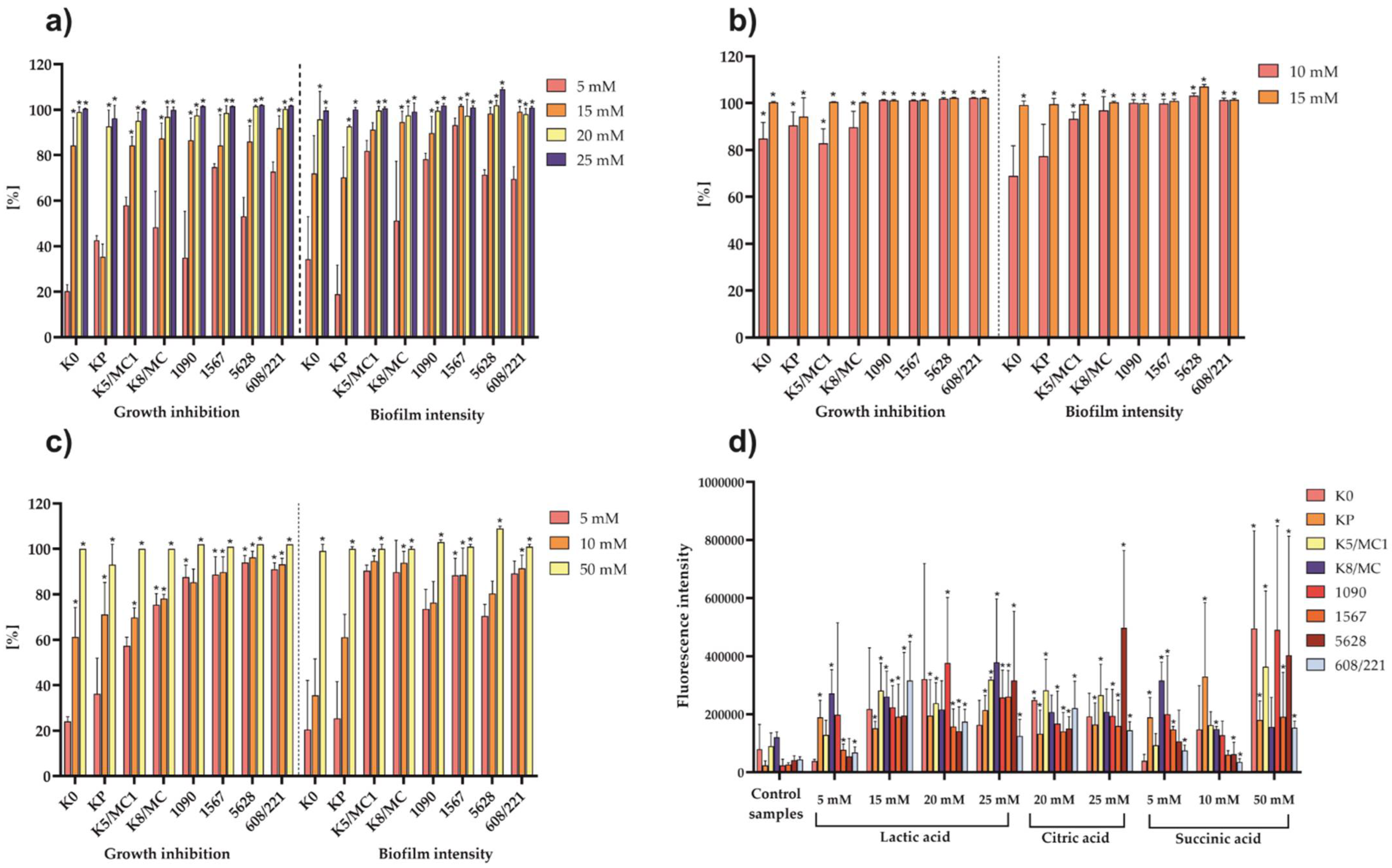
2.4. The Effect of Selected Organic Acids on P. mirabilis Cells
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Lactobacillus Cell-Free Supernatant (CFS) Preparation
4.3. Well Diffusion Assay
4.4. Broth Microdilution Method
4.5. Analysis of the Profile of Organic Acids Produced by Lactobacillus
4.6. Determination of the Effects of Organic Acids on P. mirabilis Growth Inhibition and Biofilm Formation
4.7. The Impact of Organic Acids on P. mirabilis Membrane Integrity
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Martinez, R.M.; Hulten, K.G.; Bui, U.; Clarridge, J.E. Molecular Analysis and Clinical Significance of Lactobacillus spp. Recovered from Clinical Specimens Presumptively Associated with Disease. J. Clin. Microbiol. 2014, 52, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Abdelmaksoud, A.A.; Koparde, V.N.; Sheth, N.U.; Serrano, M.G.; Glascock, A.L.; Fettweis, J.M.; Strauss, J.F.; Buck, G.A.; Jefferson, K.K. Comparison of Lactobacillus crispatus isolates from Lactobacillus-dominated vaginal microbiomes with isolates from microbiomes containing bacterial vaginosis-associated bacteria. Microbiology. 2016, 162, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Petrova, M.I.; Lievens, E.; Mail, S.; Imholz, N.; Lebeer, S. Lactobacillus species as biomarkers and agents that can promote various aspects of vaginal health. Front. Physiol. 2015, 6, 81. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, A.J.; Brubaker, L. Urobiome Updated: Advances in Urinary Microbiome Research. Nat. Rev. Urol. 2019, 16, 73–74. [Google Scholar] [CrossRef] [PubMed]
- Thomas-White, K.; Forster, S.C.; Kumar, N.; Van Kuiken, M.; Putonti, C.; Stares, M.D.; Hilt, E.E.; Price, T.K.; Wolfe, A.J.; Lawley, T.D. Culturing of female bladder bacteria reveals an interconnected urogenital microbiota. Nat. Commun. 2018, 9, 1557. [Google Scholar] [CrossRef] [PubMed]
- Aragón, I.M.; Herrera-Imbroda, B.; Queipo-Ortuño, M.I.; Castillo, E.; Del Moral, J.S.; Gómez-Millán, J.; Yucel, G.; Lara, M.F. The urinary tract microbiome in health and disease. Eur. Urol. Focus. 2018, 4, 128–138. [Google Scholar] [CrossRef]
- Gottschick, C.; Deng, Z.L.; Vital, M.; Masur, C.; Abels, C.; Pieper, D.H.; Wagner-Döbler, I. The urinary microbiota of men and women and its changes in women during bacterial vaginosis and antibiotic treatment. Microbiome. 2017, 5, 99. [Google Scholar] [CrossRef]
- Torzewska, A.; Wiewiura, P.; Brodecka, D.; Szczerbiec, D.; Różalski, A. Potentially Probiotic Lactobacillus Strains Derived from Food Intensify Crystallization Caused by Proteus mirabilis in Urine. Probiotics. Antimicrob. Proteins. 2021, 13, 441–452. [Google Scholar] [CrossRef]
- Shim, Y.H.; Lee, S.J.; Lee, J.W. Antimicrobial activity of lactobacillus strains against uropathogens. Pediatr. Int. 2016, 58, 1009–1013. [Google Scholar] [CrossRef]
- Oliveira, L.C.; Silveira, A.M.M.; Monteiro, A.S.; Dos Santos, V.L.; Nicoli, J.R.; Azevedo, V.A.C.; Soares, S.C.; Dias-Souza, M.V.; Nardi, R.M.D. In silico Prediction, in vitro Antibacterial Spectrum and Physicochemical Properties of a Putative Bacteriocin Produced by Lactobacillus rhamnosus Strain L156.4. Front. Microbiol. 2017, 19, 876. [Google Scholar] [CrossRef] [Green Version]
- Todorov, S.D.; Perin, L.M.; Carneiro, B.M.; Rahal, P.; Holzapfel, W.; Nero, L.A. Safety of Lactobacillus plantarum ST8Sh and Its Bacteriocin. Probiotics Antimicrob. Proteins. 2017, 9, 334–344. [Google Scholar] [CrossRef]
- Jiang, Y.H.; Xin, W.G.; Yang, L.Y.; Ying, J.P.; Zhao, Z.S.; Lin, L.B.; Li, X.Z.; Zhang, Q.L. A novel bacteriocin against Staphylococcus aureus from Lactobacillus paracasei isolated from Yunnan traditional fermented yogurt: Purification, antibacterial characterization, and antibiofilm activity. J. Dairy. Sci. 2022, 105, 2094–2107. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Smail, G.A.; Alzeer, A.F.; Arasu, M.V.; Vijayaraghavan, P.; Choi, K.C.; Al.-Dhabi, N.A. Probiotic characteristic of Lactobacillus strains isolated from cheese and their antibacterial properties against gastrointestinal tract pathogens. Saudi J. Biol. Sci. 2022, 27, 3505–3513. [Google Scholar] [CrossRef]
- Martin, R.; Suarez, J.E. Biosynthesis and Degradation of H2O2 by Vaginal Lactobacilli. Appl. Environ. Microbiol. 2010, 76, 400–405. [Google Scholar] [CrossRef]
- Hu, C.H.; Ren, L.Q.; Zhou, Y.; Ye, B.C. Characterization of antimicrobial activity of three Lactobacillus plantarum strains isolated from Chinese traditional dairy food. Food Sci. Nutr. 2019, 7, 1997–2005. [Google Scholar] [CrossRef] [PubMed]
- Khalil, E.S.; Abd Manap, M.Y.; Mustafa, S.; Alhelli, A.M.; Shokryazdan, P. Probiotic Properties of Exopolysaccharide-Producing Lactobacillus Strains Isolated from Tempoyak. Molecules. 2018, 23, 398. [Google Scholar] [CrossRef]
- Armbruster, C.E.; Mobley, H.L.T.; Pearson, M.M. Pathogenesis of Proteus mirabilis infection. EcoSal. Plus. 2018, 8, 10. [Google Scholar] [CrossRef]
- Yuan, F.; Huang, Z.; Wang, G.; Li, P.; Yang, B.; Li, J. Pathogenesis of Proteus mirabilis in Catheter-Associated Urinary Tract Infections. Urol. Int. 2021, 105, 354–361. [Google Scholar] [CrossRef]
- Jones, S.M.; Yerly, J.; Hu, Y.; Ceri, H.; Martinuzzi, R. Structure of Proteus mirabilis biofilms grown in artificial urine and standard laboratory media. FEMS. Microbiol. Lett. 2007, 268, 16–21. [Google Scholar] [CrossRef]
- Klein, R.D.; Hultgren, S.J. Urinary tract infections: Microbial pathogenesis, host-pathogen interactions and new treatment strategies. Nat. Rev. Microbiol. 2020, 18, 211–226. [Google Scholar] [CrossRef]
- Shokouhfard, M.; Kermanshahi, R.K.; Feizabadi, M.M.; Teimourian, S.; Safari, F. Lactobacillus spp. derived biosurfactants effect on expression of genes involved in Proteus mirabilis biofilm formation. Infect. Genet. Evol. 2022, 100, 105264. [Google Scholar] [CrossRef] [PubMed]
- Jayalath, S.; Magana-Arachchi, D. Dysbiosis of the Human Urinary Microbiome and its Association to Diseases Affecting the Urinary System. Indian J. Microbiol. 2022, 62, 153–166. [Google Scholar] [CrossRef]
- Lee, H.Y.; Wang, J.W.; Juan, Y.S.; Li, C.C.; Liu, C.J.; Cho, S.Y.; Yeh, H.C.; Chueh, K.S.; Wu, W.J.; Wu, D.C. The impact of urine microbiota in patients with lower urinary tract symptoms. Ann. Clin. Microbiol. Antimicrob. 2021, 20, 23. [Google Scholar] [CrossRef]
- Kawalec, A.; Zwolińska, D. Emerging Role of Microbiome in the Prevention of Urinary Tract Infections in Children. Int. J. Mol. Sci. 2022, 23, 870. [Google Scholar] [CrossRef]
- Lewis, A.L.; Gilbert, N.M. Roles of the vaginal microbiota in urinary tract infection: Evidence from clinical correlations and experimental models. GMS Infect. Dis. 2020, 8, Doc02. [Google Scholar]
- Perez-Carrasco, V.; Soriano-Lerma, A.; Soriano, M.; Gutiérrez-Fernández, J.; Garcia-Salcedo, J.A. Urinary microbiome: Yin and Yang of the Urinary Tract. Front. Cell Infect. Microbiol. 2021, 11, 617002. [Google Scholar] [CrossRef]
- Hrbacek, J.; Tlaskal, V.; Cermak, P.; Hanacek, V.; Zachoval, R. Bladder Microbiota Are Associated with Clinical Conditions That Extend beyond the Urinary Tract. Microorganisms 2022, 10, 874. [Google Scholar] [CrossRef]
- Argentini, C.; Fontana, F.; Alessandri, G.; Lugli, G.A.; Mancabelli, L.; Ossiprandi, M.C.; van Sinderen, D.; Ventura, M.; Milani, C.; Turroni, F. Evaluation of Modulatory Activities of Lactobacillus crispatus Strains in the Context of the Vaginal Microbiota. Microbiol. Spectr. 2022, 27, e0273321. [Google Scholar] [CrossRef]
- Bungenstock, L.; Abdulmawjood, A.; Reich, F. Evaluation of antibacterial properties of lactic acid bacteria from traditionally and industrially produced fermented sausages from Germany. PLoS ONE 2020, 15, e0230345. [Google Scholar] [CrossRef]
- Schaffer, J.N.; Pearson, M.M. Proteus mirabilis and Urinary Tract Infections. Microbiol. Spectr. 2015, 3, UTI-0017-2013. [Google Scholar] [CrossRef]
- Filipiak, A.; Chrapek, M.; Literacka, E.; Wawszczak, M.; Głuszek, S.; Majchrzak, M.; Wróbel, G.; Łysek-Gładyńska, M.; Gniadkowski, M.; Adamus-Białek, W. Pathogenic Factors Correlate with Antimicrobial Resistance among Clinical Proteus mirabilis Strains. Front. Microbiol. 2020, 11, 579389. [Google Scholar] [CrossRef] [PubMed]
- Mokracka, J.; Gruszczyńska, B.; Kaznowski, A. Integrons, β-lactamase and qnr genes in multidrug resistant clinical isolates of Proteus mirabilis and P. vulgaris. APMIS. 2012, 120, 950–958. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, A.; Habibi, M.; Bouzari, S.; Asadi Karam, M.R. Characterization of Antibiotic-Susceptibility Patterns, Virulence Factor Profiles and Clonal Relatedness in Proteus mirabilis Isolates from Patients with Urinary Tract Infection in Iran. Infect. Drug. Resist. 2019, 12, 3967–3979. [Google Scholar] [CrossRef] [PubMed]
- Shaaban, M.; Abd El-Rahman, O.A.; Al-Qaidi, B.; Ashour, H.M. Antimicrobial and Antibiofilm Activities of Probiotic Lactobacilli on Antibiotic-Resistant Proteus mirabilis. Microorganisms 2020, 8, 960. [Google Scholar] [CrossRef]
- Koohestani, M.; Moradi, M.; Tajik, H.; Badali, A. Effects of cell-free supernatant of Lactobacillus acidophilus LA5 and Lactobacillus casei 431 against planktonic form and biofilm of Staphylococcus aureus. Vet. Res. Forum. 2018, 9, 301–306. [Google Scholar]
- Tejero-Sarinena, S.; Barlow, J.; Costabile, A.; Gibson, G.R.; Rowland, I. In vitro evaluation of the antimicrobial activity of a range of probiotics against pathogens: Evidence for the effects of organic acids. Anaerobe 2012, 18, 530–538. [Google Scholar] [CrossRef]
- Ju, H.; Chen, H.; Xiang, A.; Wang, Y.; Yue, T.; Yuan, Y. Identification and characterization of Lactobacillus paracasei strain MRS-4 antibacterial activity against Alicyclobacillus acidoterrestris. LWT 2021, 150, 111991. [Google Scholar] [CrossRef]
- Bukhari, S.A.; Salman, M.; Numan, M.; Javed, M.R.; Zubair, M.; Mustafa, G. Characterization of antifungal metabolites produced by Lactobacillus plantarum and Lactobacillus coryniformis isolated from rice rinsed water. Mol. Biol. Rep. 2020, 47, 1871–1881. [Google Scholar] [CrossRef]
- Neal-McKinney, J.M.; Lu, X.; Duong, T.; Larson, C.L.; Call, D.R.; Shah, D.H.; Konkel, M.E. Production of organic acids by probiotic lactobacilli can be used to reduce pathogen load in poultry. PLoS ONE 2012, 7, e43928. [Google Scholar]
- Gezginc, Y.; Topcal, F.; Comertpay, S.; Akyol, I. Quantitative analysis of the lactic acid and acetaldehyde produced by Streptococcus thermophilus and Lactobacillus bulgaricus strains isolated from traditional Turkish yogurts using HPLC. J. Dairy Sci. 2015, 98, 1426–1434. [Google Scholar] [CrossRef]
- Burns, J.; McCoy, C.P.; Irwin, N.J. Synergistic activity of weak organic acids against uropathogens. J. Hosp. Infect. 2021, 111, 78–88. [Google Scholar] [PubMed]
- Kwiecińska-Piróg, J.; Skowron, K.; Zniszczol, K.; Gospodarek, E. The assessment of Proteus mirabilis susceptibility to ceftazidime and ciprofloxacin and the impact of these antibiotics at subinhibitory concentrations on Proteus mirabilis biofilms. Biomed. Res. Int. 2013, 2013, 930876. [Google Scholar] [PubMed]
- Srivastava, D.; Srivastava, S.; Singh, P.C.; Kumar, A. Mechanisms of Biofilm Development, Antibiotic Resistance and Tolerance and Their Role in Persistent Infections. In Antibacterial Drug Discovery to Combat MDR; Ahmad, I., Ahmad, S., Rumbaugh, K., Eds.; Springer: Singapore, 2019; pp. 115–130. [Google Scholar]
- Yang, K.M.; Kim, J.S.; Kim, H.S.; Kim, Y.Y.; Oh, J.K.; Jung, H.W.; Park, D.S.; Bae, K.H. Lactobacillus reuteri AN417 cell-free culture supernatant as a novel antibacterial agent targeting oral pathogenic bacteria. Sci. Rep. 2021, 11, 1631. [Google Scholar]
- Rossoni, R.D.; Velloso, M.D.S.; de Barros, P.P.; de Alvarenga, J.A.; Santos, J.D.D.; Santos Prado, A.C.C.D.; Ribeiro, F.C.; Anbinder, A.L.; Junqueira, J.C. Inhibitory effect of probiotic Lactobacillus supernatants from the oral cavity on Streptococcus mutans biofilms. Microb. Pathog. 2018, 123, 361–367. [Google Scholar] [PubMed] [Green Version]
- Ray Mohapatra, A.; Jeevaratnam, K. Inhibiting bacterial colonization on catheters: Antibacterial and antibiofilm activities of bacteriocins from Lactobacillus plantarum SJ33. J. Glob. Antimicrob. Resist. 2019, 19, 85–92. [Google Scholar] [PubMed]
- Peh, E.; Kittler, S.; Reich, F.; Kehrenberg, C. Antimicrobial activity of organic acids against Campylobacter spp. and development of combinations- A synergestic effect? PLoS ONE 2020, 15, e0239312. [Google Scholar]
- Alakomi, H.L.; Skytta, E.; Saarela, M.; Mattila-Sandholm, T.; Latva-Kala, K.; Helander, I.M. Lactic acid permeabilizes gram-negative bacteria by disrupting the outer membrane. Appl. Environ. Microbiol. 2010, 66, 2001–2005. [Google Scholar]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters, Version 11.0, 2021. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_12.0_Breakpoint_Tables.pdf (accessed on 19 April 2022).
- Prabhurajeshwar, C.; Chandrakanth, K. Evaluation of antimicrobial properties and their substances against pathogenic bacteria in-vitro by probiotic Lactobacilli strains isolated from commercial yoghurt. Clin. Nutr. 2018, 23, 97–115. [Google Scholar] [CrossRef]
- Chen, C.C.; Lai, C.C.; Huang, H.L.; Huang, W.Y.; Toh, H.S.; Weng, T.C.; Chuang, Y.C.; Lu, Y.C.; Tang, H.J. Antimicrobial Activity of Lactobacillus Species against Carbapenem-Resistant Enterobacteriaceae. Front. Microbiol. 2019, 10, 789. [Google Scholar]
- Borshchevskaya, L.N.; Gordeeva, T.L.; Kalinina, A.N.; Sineokii, S.P. Spetrophotometric Determination of Lactic Acid. J. Anal. Chem. 2016, 71, 755–758. [Google Scholar] [CrossRef]
- Maszewska, A.; Moryl, M.; Wu, J.; Liu, B.; Feng, L.; Rozalski, A. Amikacin and bacteriophage treatment modulates outer membrane proteins composition in Proteus mirabilis biofilm. Sci. Rep. 2021, 11, 1522. [Google Scholar] [PubMed]
- Stolarek, P.; Różalska, S.; Bernat, P. Lipidomic adaptations of the Metarhizium robertsii strain in response to the presence of butyltin compounds. Biochim. Biophys. Acta. Biomembr. 2019, 1861, 316–326. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic Used | Resistant Strains | % of Resistance |
|---|---|---|
| Ampicillin | K0; KP; 1090; 1567; 5628; 608/221 | 75% |
| Ampicillin/sulbactam | K0; KP; 1090; 1567; 5628; 608/221 | 75% |
| Piperacillin/tazobactam | 1090; 608/221 | 25% |
| Cefepime | 1090; 5628; 608/221 | 38% |
| Cefotaxime | KP, 1090; 1567; 5628; 608/221 | 63% |
| Ceftazidime | KP; 1090; 5628; 608/221 | 50% |
| Imipenem | - | 0% |
| Meropenem | KP | 13% |
| Aztreonam | - | 0% |
| Ciprofloxacin | 608/221 | 13% |
| Norfloxacin | - | 0% |
| Amikacin | KP | 13% |
| Gentamicin | K0; KP; 1090; 1567; 5628; 608/221 | 75% |
| Tigecycline | 1090; 608/221 | 25% |
| Nitrofurantoin | K0; K5/MC1; K8/MC; 1090; 1567 | 63% |
| P. mirabilis Strains | 1.2 | 3.2 | 4 | 6.2 | 13.2 | 22.2 | 35.3 | 40 | 41.4 | ATCC 33197 | ATCC 33323 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| K0 | 98% | 98% | 95% | 79% | 97% | 73% | 97% | 97% | 97% | 98% | 100% |
| KP | 97% | 98% | 95% | 74% | 96% | 64% | 96% | 96% | 97% | 98% | 100% |
| K5/MC1 | 97% | 97% | 93% | 72% | 72% | 95% | 72% | 94% | 95% | 96% | 100% |
| K8/MC | 98% | 99% | 94% | 69% | 96% | 65% | 96% | 96% | 96% | 97% | 100% |
| 1090 | 97% | 98% | 95% | 65% | 96% | 70% | 96% | 96% | 96% | 97% | 100% |
| 1567 | 98% | 98% | 95% | 66% | 97% | 62% | 96% | 96% | 97% | 97% | 100% |
| 5628 | 97% | 97% | 93% | 92% | 95% | 69% | 95% | 89% | 95% | 96% | 100% |
| 608/221 | 97% | 98% | 94% | 63% | 95% | 69% | 95% | 91% | 96% | 96% | 99% |
| Lactobacillus Strains. | Lactic Acid | Citric Acid | Succinic Acid |
|---|---|---|---|
| Concentration (mM) | |||
| L. crispatus 1.2 | 25.09 | - | 11.21 |
| L. saerimneri 3.2 | 21.46 | - | 4.91 |
| L. crispatus 4 | 19.24 | - | 36.51 |
| L. crispatus 6.2 | 19.24 | - | 17.12 |
| L. crispatus 13.2 | 17.87 | 6.37 | 4.4 |
| L. jensenii 22.2 | 13.32 | - | 52.11 |
| L. gasseri 35.3 | 10.28 | - | 0.91 |
| L. crispatus 40 | 12.49 | 14.12 | 1.81 |
| L. crispatus 41.4 | 16.06 | - | 8.51 |
| L. crispatus ATCC 33197 | 9.58 | - | 4.71 |
| L. gasseri ATCC 33323 | 7.59 | 11.52 | 4.61 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szczerbiec, D.; Piechocka, J.; Głowacki, R.; Torzewska, A. Organic Acids Secreted by Lactobacillus spp. Isolated from Urine and Their Antimicrobial Activity against Uropathogenic Proteus mirabilis. Molecules 2022, 27, 5557. https://doi.org/10.3390/molecules27175557
Szczerbiec D, Piechocka J, Głowacki R, Torzewska A. Organic Acids Secreted by Lactobacillus spp. Isolated from Urine and Their Antimicrobial Activity against Uropathogenic Proteus mirabilis. Molecules. 2022; 27(17):5557. https://doi.org/10.3390/molecules27175557
Chicago/Turabian StyleSzczerbiec, Dominika, Justyna Piechocka, Rafał Głowacki, and Agnieszka Torzewska. 2022. "Organic Acids Secreted by Lactobacillus spp. Isolated from Urine and Their Antimicrobial Activity against Uropathogenic Proteus mirabilis" Molecules 27, no. 17: 5557. https://doi.org/10.3390/molecules27175557